Of the most abundant protozoa, free-living amoebae (FLA) have been described from freshwater, marine, and soil habitats as well as extreme environments (Page, 1967; Rogerson and Patterson, 2002; Park et al., 2009). The term ‘FLA’ is regarded as a single functional group, but FLA are systematically composed of diverse groups. They mainly belong to Lobosea (or Lobosa) and Heterolobosea at the class level (Smirnov and Brown, 2004). Lobosea is a major amoeboid group showing cylindrical or blunt and broad pseudopodia within the phylum Amoebozoa. This group may be closely related to the common ancestor of animals and fungi (Cavalier-Smith, 2013). Recently, the class Lobosea moved to the subphylum ‘Lobosa’ including the classes Tubulinea and Discosea (Smirnov et al., 2011). Another important amoeboid group is Heterolobosea within the phylum Excavata. Heterolobosea typically have eruptive movement, and are morphologically diverse groups including flagellates, amoeboflagellates, and amoebae (Patterson et al., 2002; Rogerson and Patterson, 2002; Park et al., 2007, 2009, 2012; Park and Simpson, 2011). Some of these groups are commonly thought of as potentially human pathogens. For instance, Acanthamoeba on contact lens was the causative agent of human keratitis that significantly decreased visual acuity (Duguid et al., 1997). Naegleria fowleri is an opportunistic pathogen associated with primary amoebic meningoencephalitis, a lethal brain infection (De Jonkheere, 2002; Patterson et al., 2002). However, due to similar morphological features and moving behavior of FLA, classification of FLA tends to be restrictive under light microscopy only. Therefore, light microscopic observations, ultrastructures (not common), and molecular phylogenetic analysis are usually required for identification and classification of FLA (Corliss, 2001). Potentially pathogenic FLA are still insufficiently described using modern techniques to date, particularly in Korea.
Vermamoeba vermiformis was amoeboid protozoa belonging to Lobosea, and was firstly created as Hartmannella vermiformis by Page 1967. More recently, Smirnov et al. (2011) renamed as Vermamoeba vermiformis because of different length/width ratio and cylindrical form. In addition, all published phylogenetic trees suggested that Vermamoeba formed a sister group with Echinamoeba, rather than other ‘true’ Hartmannella groups (Bolivar et al., 2001; Corsaro et al., 2010; Brown et al., 2011; Smirnov et al., 2011; Watson et al., 2014; Cavalier-Smith et al., 2015). Vermamoeba vermiformis is special interest for human health. Legionella pneumophila, a bacterial parasite associated with Legionnaires’ disease, could infect in V. vermiformis as host, then was propagated in the host cells (Brieland et al., 1997). Furthermore, V. vermiformis seems to be potential causative agents of human keratitis (Aitken et al., 1996), although there is a skeptical view (De Jonckheere and Brown, 1999). Trophozoites of V. vermiformis formed cylindrical monopodia with no uroidal filaments (Page, 1967). Length of V. vermiformis was usually 14-31 μm, and length of cell body was ~6 times more than width (Page, 1967). No flagellate form had been observed (Page, 1967). Cysts were circular or slightly ovoid shape surrounded by double-wall, and lacked cyst pores (Smirnov and Michel, 1999). Vermamoeba vermiformis was commonly found in freshwater and soil environments worldwide. However, this interesting species has not been reported from natural environments in Korea.
Here, Vermamoeba vermiformis strain MG1 was firstly isolated from a freshwater pond, Mulgol, of Dokdo in the East Sea. Morphology and 18S rRNA gene sequence of V. vermiformis strain MG1 were very similar to previously reported species. Morphology and 18S rRNA gene sequence of V. vermiformis is recorded in Korea for the first time.
Vermamoeba (=Hartmannella) vermiformis strain ‘MG1’ was isolated from a freshwater pond called Mulgol (Fig. 1A) in Dokdo (island; ~87 km apart from Ulleung island in Korea, 37°14ʹ22ʺN, 131°52ʹ08ʺE) of the East Sea, Korea. Historically, Mulgol was a unique pond to supply drinking water for residents in Dokdo, but was now reconstructed as a water well with lid to protect from pollutants (Fig. 1B). A culture was established through a plate-cultivation method. A freshwater sample collected in Jun 2011 was used to innoculate plates of Page’s amoeba saline (PAS) (Page, 1988) supplemented with 2.0% agar (w/v, final conc.), with Escherichia coli added as a food source. For routine maintenance the cultures were incubated at 20-25°C in liquid PAS with 0.5% Luria-Bertani broth (v/v, final conc.), and subculturing was performed every 2-4 weeks. Actively growing V. vermiformis strain ‘MG1’ from plates was directly inoculated into sterile seawater (30 mL, 35‰ salinity) supplemented with Escherichia coli to assess its halophilicity for 2-4 weeks.
Live trophozoites and cysts mounted on glass slides were observed with differential interference microscopy using a Leica DM5500B microscope equipped with a DFC550 digital camera (Leica, Wetzlar, Germany). Pictures were taken with a digital camera and the dimensions measured on these pictures.
Nucleic acids from the isolate were prepared using a DNeasy Blood and Tissue Kit (Qiagen, Germantown, MD, USA) as described by the supplied protocol. Amplification of 18S rRNA genes was performed using standard polymerase chain reaction (PCR) protocols with eukaryote-specific primers EukA and EukB (Medlin et al., 1988). The total volume of the PCR mixture was 20 μL. The reaction mixture contained 50-100 ng of DNA, 0.2 mM deoxynucleoside triphosphate, 0.5 μM each primer, 2 mM MgCl2, and 2.5 U of Taq DNA polymerase (Solgent, Daejeon, Korea). PCR amplifications were conducted by the following cycle parameters: an initial denaturation step (5 min, 94°C) was followed by 30 cycles consisting of denaturation (45 s, 94°C), annealing (1 min, 55°C), and extension (3 min, 72°C), with a final extension step for 20 min at 72°C. Amplicons were cloned into a pGEM-T Easy vector, and four or five positive clones were partially sequenced using vector sequencing primer T7, then one positive clone was completely sequenced on both strands using vector sequencing primer SP6, plus various eukaryotic internal sequencing primers.
Representative 18S rRNA gene sequences from 29 FLA species within the Tubulinea (naked amoebae) including Vermamoeba vermiformis isolate ‘MG1’ were used for phylogenetic analysis. A dataset was included 1,355 unambiguously aligned sites, and were aligned by eye. This alignment is available on request. Phylogenetic trees for a dataset were inferred by maximum-likelihood (ML) and by Bayesian analysis. The GTR+gamma+I model of sequence evolution models was selected for a dataset using MrModeltest 2.2 (Nylander, 2004). The ML tree was estimated using RAxML-VI-HPC v.7 (Stamatakis, 2006) with the GTRGAMMAI model setting, 500 random starting taxon addition sequences, and statistical support estimated using bootstrapping with 10,000 replicates. The Bayesian analysis was conducted in MrBayes 3.2 (Ronquist et al., 2012) with two independent runs, each with four chains running for 5,000,000 generations (burn-in 1,500,000 generations, average standard deviation of split frequencies for last 75% of generations was <0.003), with default heating parameter (0.1) and sampling frequency (0.01).
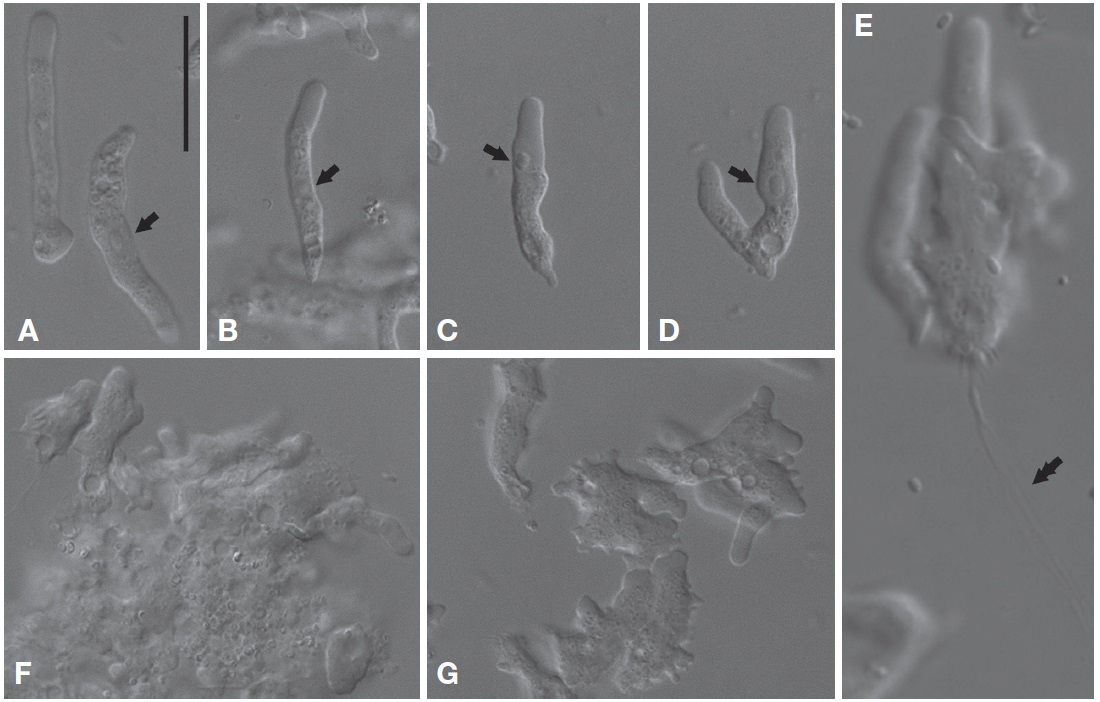
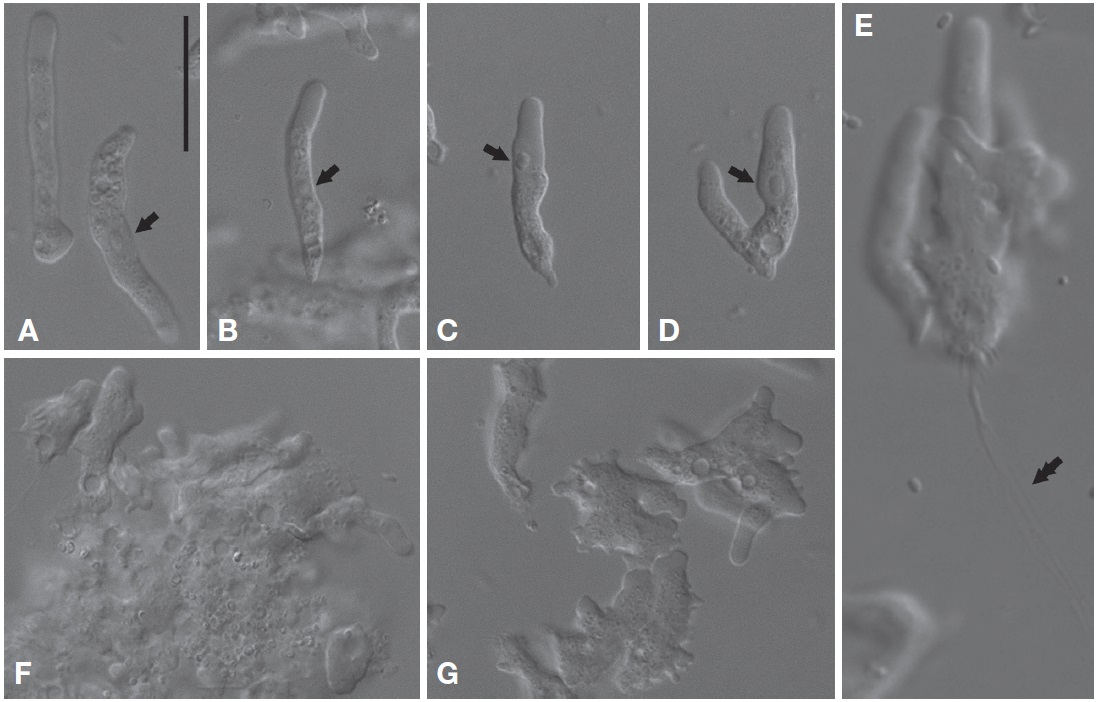
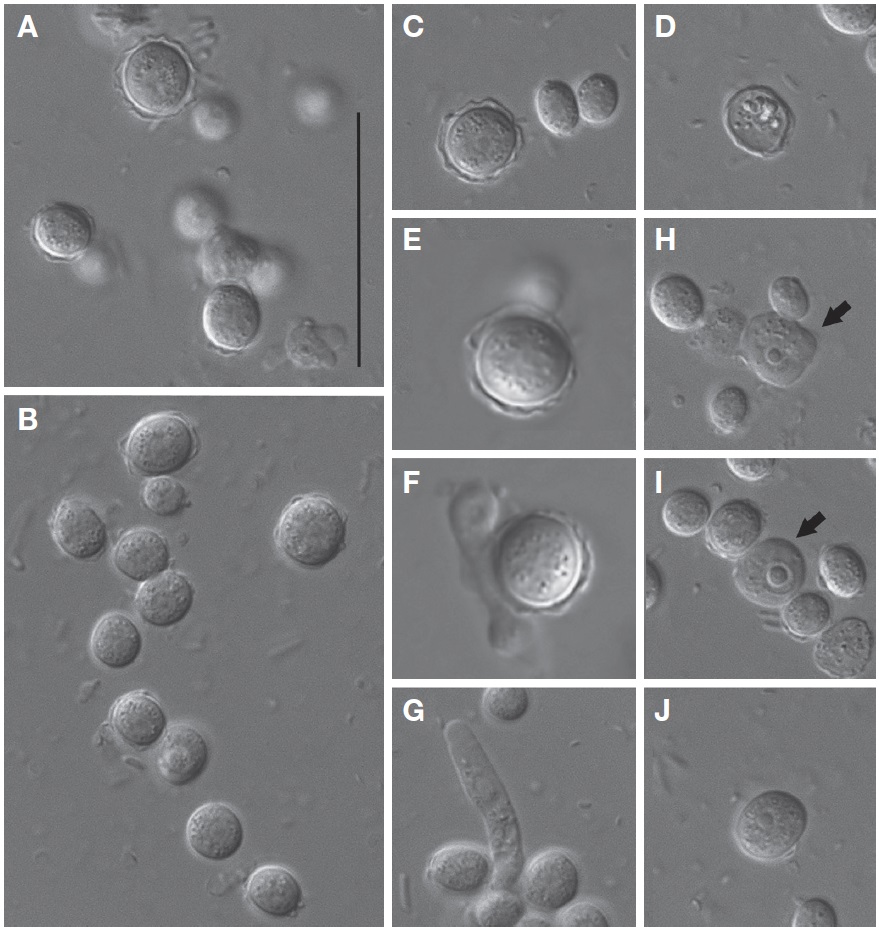
Morphological description. Trophozoites of V. vermiformis usually had well-visible cylindrical monopodia in liquid medium, rather than eruptive hyaline front (Fig. 2A-C). The length and width of the isolate were 19.9-36.6 μm (mean±SD, 28.8±5.2 μm; n=20) and 3.0-6.3 μm (mean±SD, 4.5±0.9 μm; n=20), respectively (Fig. 2A-D). The average ratio of length to width was approximately 6.5 (range, 4.7-11.4). Some cells temporarily had two pseudopods (or more), and formed irregular shapes (Fig. 2F, G). Later, the cells were changed to predominant cylindrical form with monopodia. When locomotive amoebae started moving in one direction, new protruding pseudopods were elongated to the direction of movement, such that the cells had branch or horseshoe shapes (Fig. 2B, D). Intriguingly, some cells uncommonly displayed adhesive uroidal filaments in the posterior part of the cells (Fig. 2E). Cells had a large sub-circular nucleus with a central nucleolus in a middle part of the cells (Fig. 2A, B), or sometimes in an anterior part of the cells nearby hyaline cap (Fig. 2C, D). Flagellates form was not observed in liquid culture or on solid plates to date. Several contractile vacuoles were commonly observed in a posterior end of the cells (Fig. 2A-C).
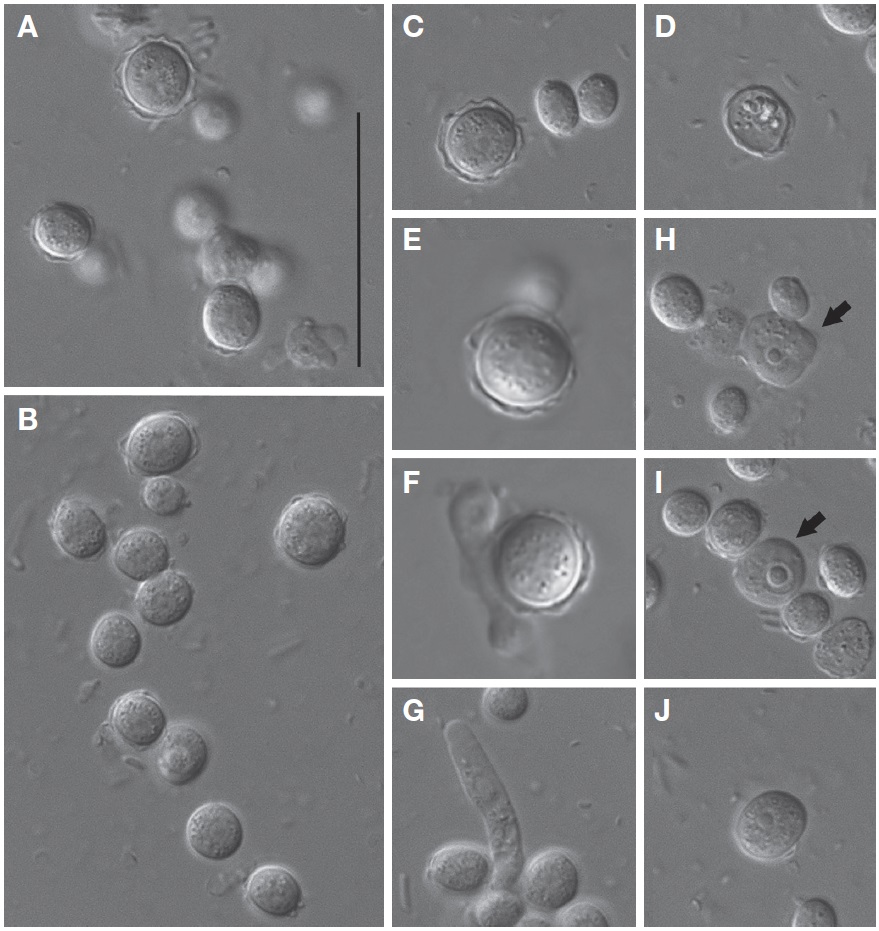
Cysts of V. vermiformis strain MG1 formed a spherical or slightly ovoidal shape with smooth-walled or irregular outline, and ranged from 5.1 to 10.0 μm (mean±SD, 7.5±1.3 μm; n=20) in diameter (Fig. 3A-C). The cyst cytoplasm was commonly granular (Fig. 3A-C). Cysts appeared to separate the inner wall (endocyst) from the outer wall (ectocyst) (Fig. 3A, C). Endocyst fit closely to the cyst cytoplasm, but ectocyst appeared to have variable thickness (Fig. 3A-C). Typical cysts had no distinct bulges of ectocyst layer, and were smooth-walled (Fig. 3C). Cysts in the absence of cytoplasm were sometimes observed, assuming excysted profile (Fig. 3D). Interestingly, processes of excystment (Fig. 3E-G) and encystment (Fig. 3H, J) were observed in V. vermiformis strain MG1. Cytoplasmic projections were generated from aged cysts during excystment (Fig. 3E, F), then cylindrical monopodia was escaped from aged cysts (Fig. 3G). Typical cylindrical amoebae firstly formed irregular shapes during encystment (Fig. 3H). Irregular-shaped amoebae gradually changed to the circular forms, and which of them formed cysts with smooth-walled later (Fig. 3I, J). No cyst pores were observed.
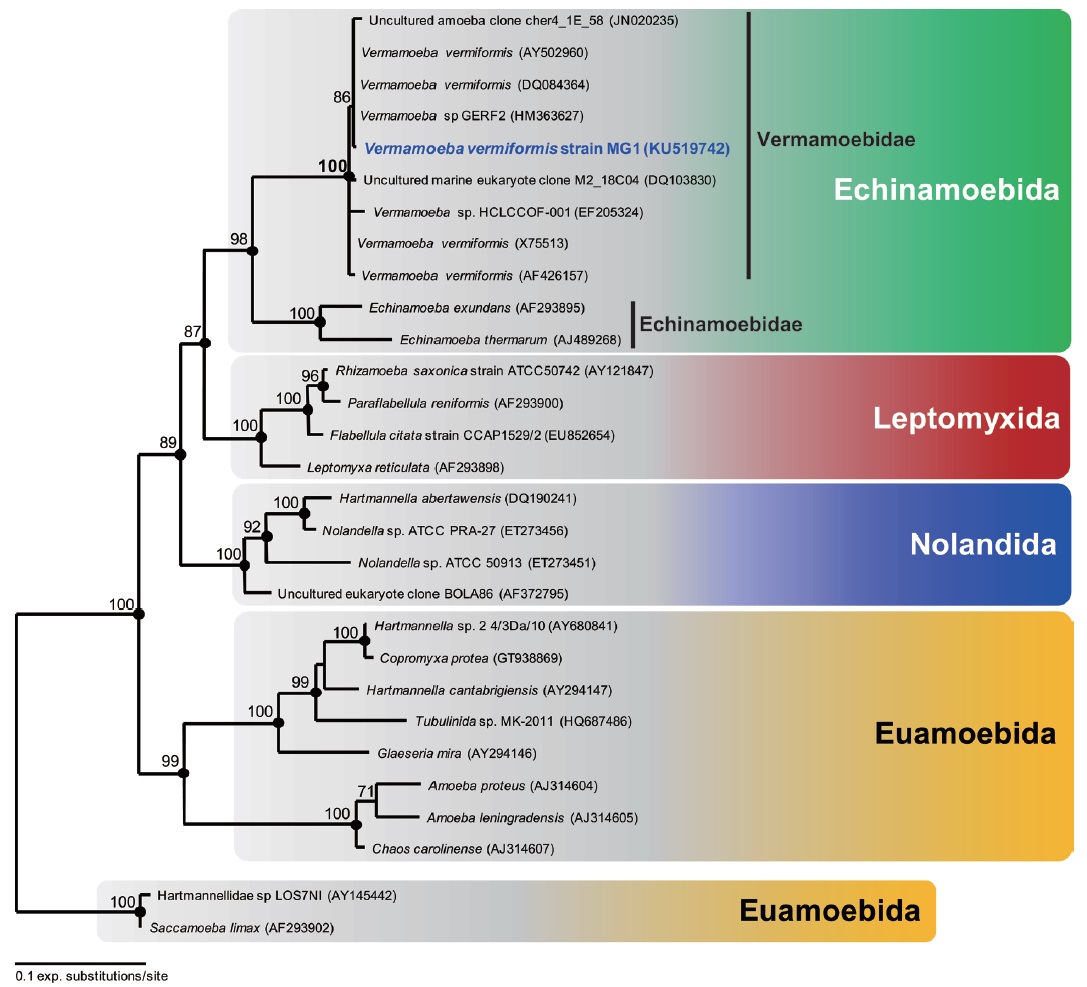
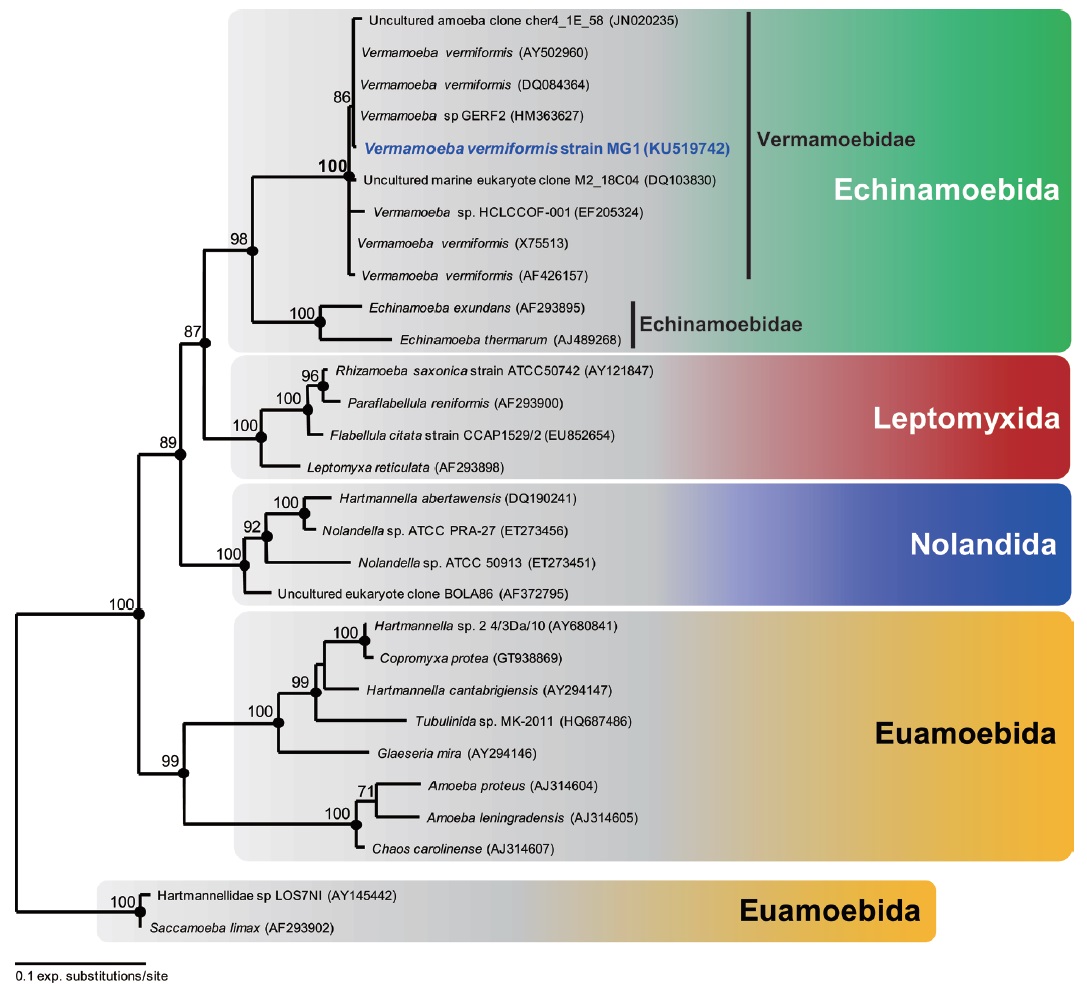
Molecular phylogeny. The 18S rRNA gene sequence of V. vermiformis strain MG1 was 1,838 bp long. The most similar sequence returned by BLASTN search was Vermamoeba vermiformis strains. All other V. vermiformis, including V. vermiformis strain MG1 and two environmental sequences, clearly showed a monophyletic group belonging to Vermamoebidae with maximum bootstrap support (maximum likelihood, 100%), and Bayesian posterior probability of 1 (Fig. 4). The order Echinamoebida was divided into Vermamoebidae and Echinamoebidae at the family level (Fig. 4), and Vermameobidae formed a sister group with Echinamoebidae with high bootstrap support (maximum likelihood, 98%) and Bayesian posterior probability of 1 (Fig. 4).
Gene sequence. The 18S rRNA gene from isolate MG1 has been deposited in GenBank under the accession number KU519742.
Distribution. Global distribution in freshwater and soil environments.
Remarks. Morphology of Vermamoeba vermiformis strain MG1 was very similar to previously reported species under a light microscope (Page, 1967; Smirnov and Brown, 2004). Smirnov et al. (2011) reported that typical trophozoites of Vermamoeba displayed worm-like cell body, cell length/width ratio >6, stable anterior hyaline cap, and possessing a tendency branch when they changed to a direction. Morphology of cyst is a very useful key for the classification of FLA in Lobosea (Page, 1988). Cyst structure of V. vermiformis strain MG1 corresponded to previous description of V. vermiformis (Page, 1967; Smirnov and Brown, 2004; Fouque et al., 2015). As described by Smirnov and Michel (1999), cysts of V. vermiformis strain MG1 have distinct endocyst and ectocyst layers, variable thickness of ectocyst, and lack of cyst pore. However, smooth-walled ectocysts of the isolate were also observed as reported by Page (1967). Distinct shapes of ectocyst outlines may be due to the age of cysts (Dyková et al., 2005). It seems that younger cysts may be smooth-walled, rather than irregular outline (elder form). In addition, V. vermiformis strain MG1 shared 99.4%-99.9% similarity with other V. vermiformis strains. The 18S rRNA gene sequences of all reported V. vermiformis strains showed extremely high similarity with more than 98.8% (Walochnik et al., 2002). Furthermore, molecular phylogenetic analysis of 18S rRNA gene sequences confirmed that the isolate should be a member of V. vermiformis. Therefore, V. vermiformis strain MG1 from Dokdo is not distinguishable from other V. vermiformis strains (Page, 1967; Smirnov and Michel, 1999; Smirnov and Brown, 2004; Smirnov et al., 2011), and increasing known habitat range for V. vermiformis.
Uroidal filaments in the presence of V. vermiformis have not been reported in previous studies (Page, 1967; Smirnov and Brown, 2004). More recently, however, Dyková et al. (2005) reported that V. vermiformis (Hartmannella vermiformis in their paper) strains were in the presence of the posterior uroidal filaments. Therefore, it is conceivable that each strain of V. vermiformis trophozoites may have more diverse features than previously thought. Also, it is reasonable to assume that lack of uroidal filaments may not be a key character to distinct V. vermiformis from other FLA (Dyková et al., 2005).
Morphological characters of Vermamoeba vermiformis were distinct from other ‘true’ Hartmannellidae (see above). Other ‘true’ Hartmannellidae (Cashia, Copromyxa, Copromyxella, Glaeseria, Hartmannella, Ptolemeba, and Saccamoeba, see Smirnov et al., 2011; Watson et al., 2014) formed slightly clavate, and monotactic morphotype (Smirnov et al., 2005). Further, phylogenetic analysis of 18S rRNA gene sequences in Amoebozoa confirmed that Hartmannellidae is a paraphyletic group, whereas Vermamoebidae is a monophyletic group (Dyková et al., 2005; Smirnov et al., 2011; this study). Thus it is reasonable to note that previous Hartmannella vermiformis was amended as Vermamoeba vermiformis, which is a separate lineage.
Vermamoeba vermiformis-like organism was sometimes found in saline water. It is likely that the environmental sequence ‘uncultured marine eukaryote clone M2_18C04’ may be one of V. vermiformis strains (Fig. 4). The sequence was discovered in Mariager Fjord, Denmark, where salinity was between 15‰ and 20‰ (Zuendorf et al., 2006). However, V. vermiformis strain MG1 did not grow at saline water, and in most cases V. vermiformis occurred in freshwater environments. Encystment is usually observed in Vermamoebidae (Smirnov and Michel, 1999), and V. vermiformis strain MG1 showed encysted stage as well. Thus, the environmental sequence from saline Mariager Fjord may be inactive form (i.e., encysted stage).
In conclusion, V. vermiformis strain MG1 was firstly isolated from freshwater of Dokdo (island) in Korea, which was used for residents in Dokdo as drinking water. It is clear that V. vermiformis strain MG1 appears to be previously reported Vermamoeba vermiformis based on morphology and molecular phylogenetic analysis. Therefore, the freshwater of Dokdo (Mulgol) tends to be a source of potentially pathogenic amoeba V. vermiformis strain MG1. In addition, V. vermiformis would be expanded to geographic habitat. Further study is needed to assess whether V. vermiformis strain MG1 is ‘true’ causative agents of human pathogen or not.
Korean name:1*애벌레형아메바속 (신칭), 2*애벌레형아메바 (신칭)