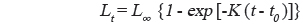가다랑어(
가다랑어는 태평양, 인도양, 대서양 등 전 세계 열대 및 온대 해역에서 주로 선망어업 등에 의해 어획되는 상업적으로 중요한 종으로, 최근 어획량이 250만 톤을 넘어서면서 그 어획량이 세계 2위를 기록하기도 하였다(FAO, 2011). 이러한 과도한 어획으로 인해 세계자연보존연맹(IUCN)에서는 가다랑어를 관심필요종(LC)으로 지정하였으며(IUCN, 2011), 지역수산관리기구(RFMOs)에서는 지속가능한 어업을 위해 보존관리조치를 강화하고 있다.
가다랑어에 대한 연령과 성장에 관한 연구로 국외에서는 등지느러미를 이용한 연령과 성장에 관한 연구(Batts, 1972), 이석을 이용한 일륜 형성 및 연령과 성장에 관한 연구(Uchiyama and Struhsaker, 1981), 가다랑어 치어의 일륜 형성에 관한 연구(Tanabe et al., 2003), 동부 인도양 가다랑어의 일륜에 관한 연구(Kayama et al., 2004) 등이 수행되었으며, 또한 온도와 산소요구량에 기초한 가다랑어의 서식지에 관한 연구(Barkley et al., 1978), 미토콘드리아 DNA 분석에 의한 대서양과 태평양 가다랑어의 유전적 유사성 연구(Graves et al., 1984) 등 다양한 생태학적 연구가 수행되었다. 국내에서는 기후변동 혹은 기후인자에 따른 중서부태평양 해역의 가다랑어 분포 및 한국 다랑어 선망어업의 어장변동에 관한 연구(An et al., 2003; Kim, 2005; Wungen, 2012), 한국 다랑어 선망어업의 조업특성에 관한 연구(Moon et al., 2005), 다랑어 선망 어획물의 종조성에 관한 연구(Lee et al., 2011) 등 대부분이 다랑어의 분포 및 어획특성과 관련된 것으로 생태학적 특성에 관한 연구는 전무한 실정이다.
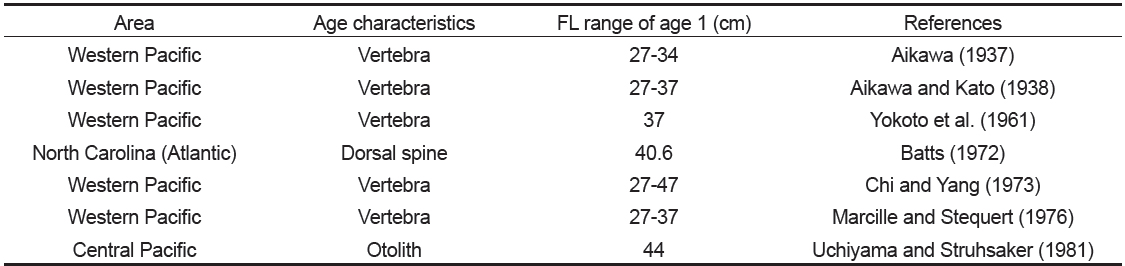
가다랑어의 연령과 성장에 대한 기존의 연구 결과를 살펴보면(Table 1), 1세의 체장(FL)이 연구자에 따라 차이를 보였는데(Aikawa, 1937; Aikawa and Kato, 1938; Batts, 1972; Chi and Yang, 1973; Kayama et al., 2007), Kayama et al. (2007)은 이러한 원인이 연령사정에 사용된 형질의 특성으로부터 기인된다고 하였다.
[Table 1.] Age characteristics used in previous studies for age and growth of Katsuwonus pelamis
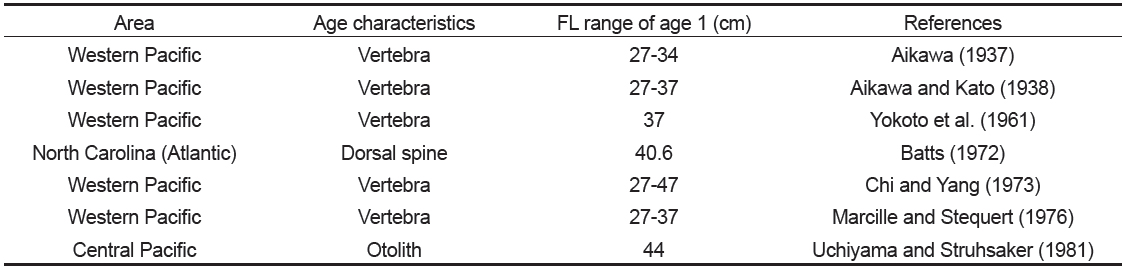
Age characteristics used in previous studies for age and growth of Katsuwonus pelamis
가다랑어의 연령과 성장에 관한 연구에 비늘(Shabotiniets, 1968; Batts, 1972), 척추골 (Aikawa, 1937; Aikawa and Kato, 1938; Yokota et al., 1961; Shomura, 1966; Shabotiniets, 1968; Chi and Yang, 1973; Marcille and Stequert, 1976), 등지느러미(Shabotiniets, 1968; Batts, 1972; Chur and Zharov, 1983; Sosa-Nishizaki et al., 1989), 이석(Uchiyama and Struhsaker, 1981) 등이 연령 형질로 사용되었는데, Batts (1972)에 의하면, 비늘은 재생된 표본의 경우 과소 추정될 가능성이 있고, 척추골은 윤이 중심으로부터 불규칙적이고 연륜으로써의 뚜렷한 특징을 보이지 않기 때문에, 비늘과 척추골보다는 등지느러미의 단면을 통해 윤을 읽는 것이 용이하다고 하였다. 그리고 Josse et al. (1979)은 이석이 다른 형질에 비해 윤문이 보다 뚜렷하므로, 윤문 판독시 이석이 가장 용이하다고 하였다.
따라서 본 연구에서는 중서부태평양 수역에서 우리나라 다랑어 선망어업에 의해 어획된 가다랑어의 이석을 사용하여 이들의 연령과 성장에 관한 생태학적 특성을 구명하고자 한다.
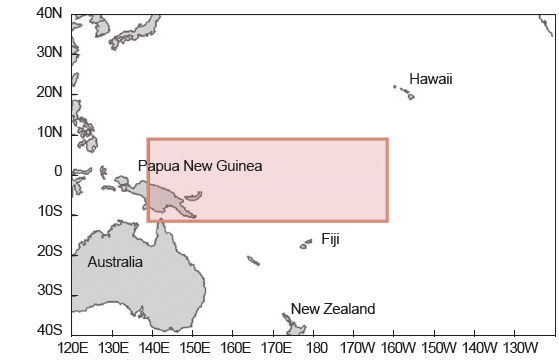
본 연구에서 사용한 가다랑어 표본은 2005년 1월부터 2006년 9월까지 중서부태평양 수역에서 우리나라 다랑어 선망어업에 의해 채집된 것이다(Fig. 1). 채집된 표본은 총 312 개체로, 가랑이체장(FL)은 0.1 cm, 체중(TW)은 0.1 kg 단위까지 측정하고 암수별 분석을 위해 성별을 구분하였다. 그러나 그 중에서 성별 구분이 모호하였던 27 개체는 암수별 분석에서 제외하였다.
가다랑어의 연령사정을 위하여 이석을 추출하였으며, 추출된 이석은 이물질을 제거하여 건조시킨 후 몰딩틀에 넣고, 레진(resin)과 하드너(hardener)를 5:1의 비율로 섞어 24시간 정도 건조시켰다. 몰딩된 이석은 연마기(Ecomet 4000, BUEHLER Co., U.S.A.)를 이용하여 3종류의 연마지(Grit 800, 1200, 2400)로 단계적으로 처리하였다. 연마된 이석은 광택을 내기위해 광택제(alumina powder)를 뿌린 후 한번 더 연마하였다.
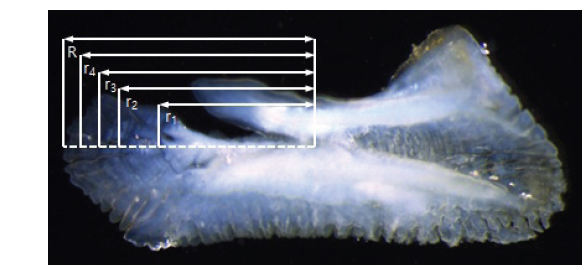
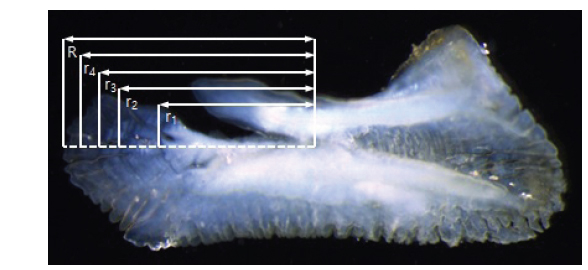
이석의 윤문은 투명대에서 불투명대로 이행하는 경계로 간주하고, 저배율 입체현미경(SZX2-ILLT, OLYMPUS Co., Japan)에 연결된 컴퓨터 영상분석 시스템(image analyzer)을 이용하여 컴퓨터 모니터 상에서 윤경을 0.001 mm 단위로 측정하였다. 이석의 반경은 핵으로부터 가장 먼 가장자리 끝 부분까지의 거리로 간주하였다(Fig. 2).
윤문판독의 정확성을 검토하기 위해 2명의 판독자가 독립적으로 연령사정을 수행하였고(Robillard et al., 2009), 판독된 결과에 대해서는 제1판독자와 제2판독자간의 일치율을 비교하였다. 일치율 비교 결과로부터 일치하지 않는 자료는 제1판독자와 제2판독자간의 합의를 통해 재판독하였다. 그리고 이석이 가다랑어의 연령 형질로써 사용가능한가를 판단하고 윤문의 대응성을 알아보고자 윤문별 윤경과 이석반경간의 관계를 분석하였다.
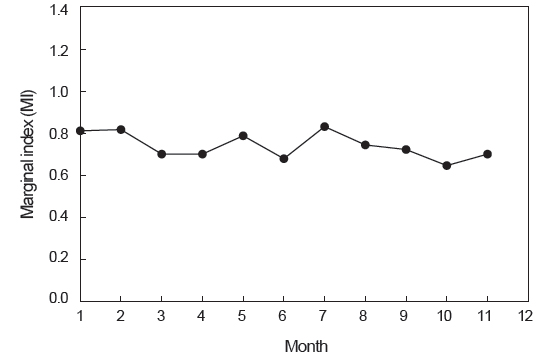
또한 윤문 형성이 주기성을 나타내는지, 그리고 그 형성시기를 알아보기 위해 연역지수(MI)의 월 변화를 추정하였다.

여기서, R은 이석의 반경, rn은 핵에서부터 n번째 윤경까지의 거리이다.
가다랑어에 대한 상대성장을 알아보기 위해 체장(FL)과 체중(TW)간의 관계를 구하였다.

여기서,
성장식의 추정은 판독된 연령과 연령별 체장 자료를 사용하여 일반적으로 가장 널리 사용되는 von Bertalanffy (1938) 성장식을 사용하였다.
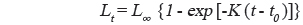
여기서, t는 연령, Lt는 연령 t세의 가랑이체장, L∞는 이론적 최대 가랑이체장, K는 성장계수, t0는 체장이 0일 때의 이론적 연령이다. 먼저 von Bertalanffy 성장식의 매개변수는 Walford방법으로 구한 후, 그 결과를 초기값으로 컴퓨터 패키지인 EXCEL의 solver를 이용하여 비선형 회귀방법으로 분석하였다(Zhang, 2010).
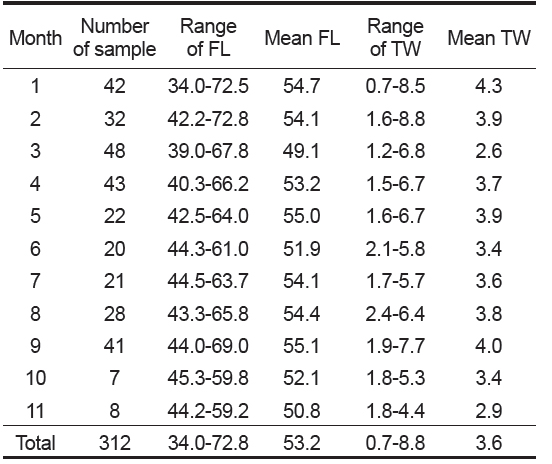
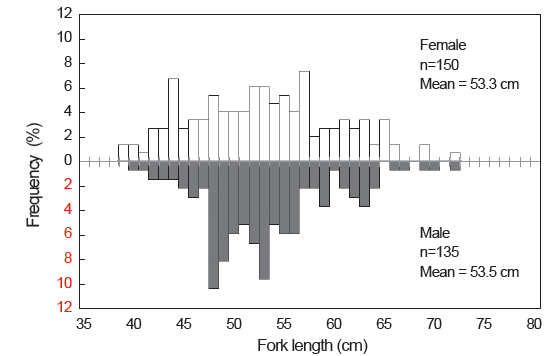
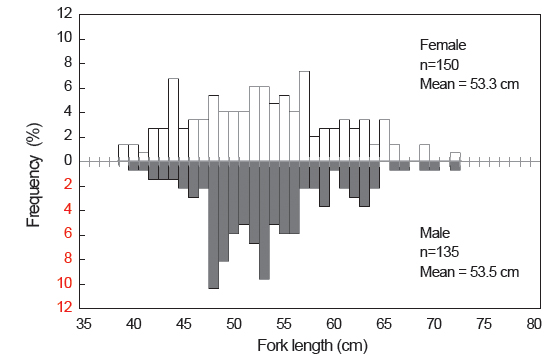
본 연구에 사용된 총 312 개체의 가랑이체장 범위는 34.0-72.8 cm이었고, 평균 가랑이체장은 53.2 cm이었으며, 체중 범위는 0.7-8.8 kg이었고, 평균 체중은 3.6 kg이었다(Table 2). 이중에서 암·수 분석에 사용된 개체는 285 개체로, 암컷이 52.6%, 수컷이 47.4%이었으며, 암컷의 가랑이체장 범위는 39.0-72.8 cm, 수컷은 40.5-72.5 cm이었고, 평균 가랑이체장은 암컷이 53.3 cm, 수컷은 53.5 cm로 서로 유사하였다(Fig. 3). 가랑이체장 조성을 살펴보면(Table 2 및 Fig. 3), 주 가랑이체장 모드는 45-55 cm 였으며, 40 cm 미만의 소형개체는 거의 채집되지 않았다.
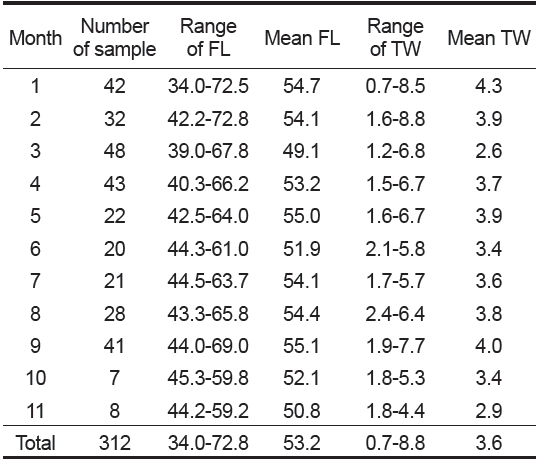
Monthly number of samples and range of fork length (FL, cm) and total weight (TW, kg) of Katsuwonus pelamis collected in the Western and Central Pacific Ocean
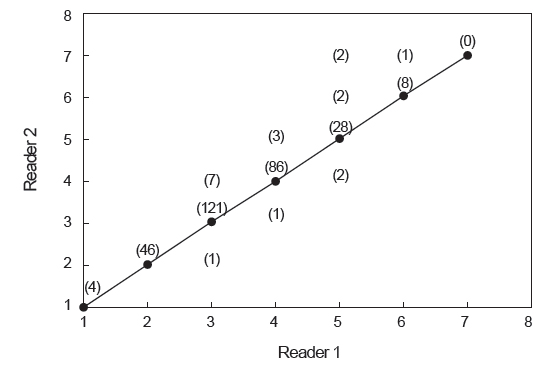
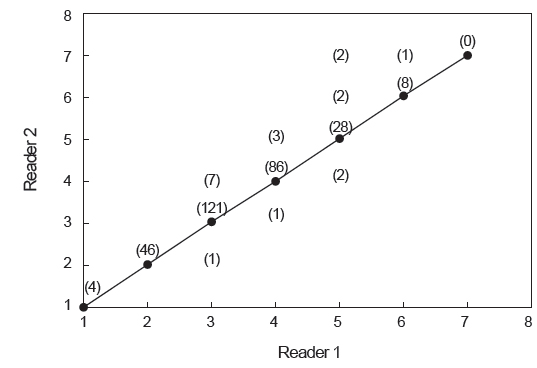
연령사정의 정확성을 검토하기 위해 2명의 판독자가 각각 수행한 연령판독 결과의 일치율은 Fig. 4와 같다. 총 312 개체의 이석 중 291 개체의 결과가 일치하여 93.3%의 일치율을 보였으며, 일치율이 가장 낮은 연령은 3세이었다. 판독 결과가 일치하지 않은 21 개체에 대해서는 두 판독자간 협의를 통해 재판독하였고, 최종 판독된 총 312 개체의 결과를 토대로 분석하였다.
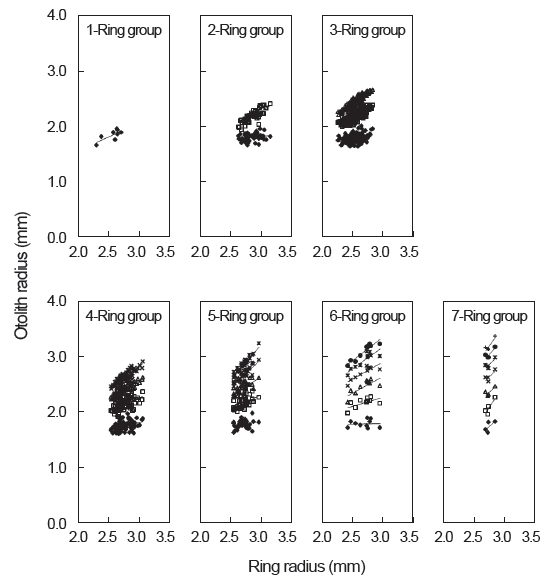
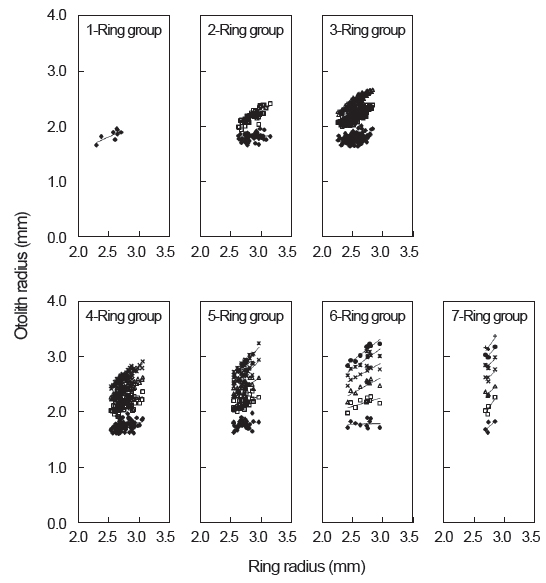
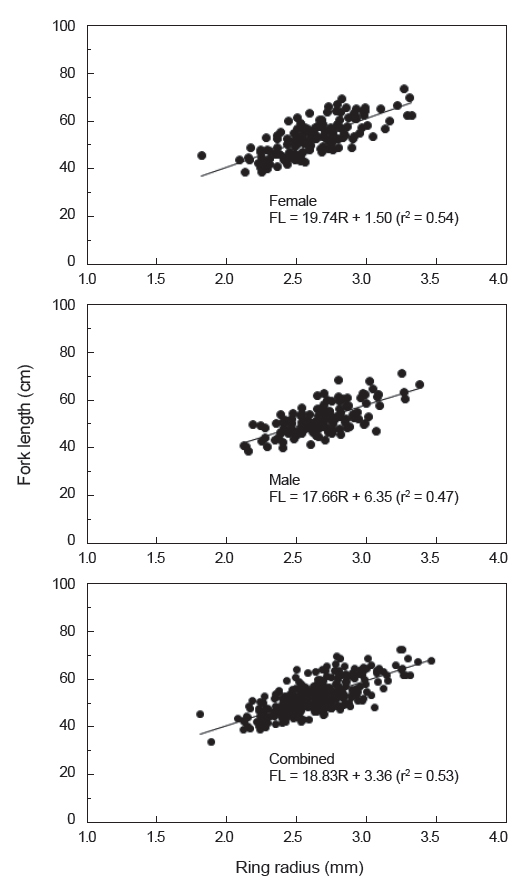
가다랑어 이석의 윤문이 연륜으로 적합한가를 알아보기 위하여 윤문별 윤경과 이석반경간의 관계를 검토한 결과(Fig. 5), 각 윤경은 인접한 윤경과 서로 중복됨이 없이 잘 분리되었고, 이석반경이 커짐에 따라 윤경 역시 커지는 것으로 나타났다. 따라서 이석이 가다랑어의 연령형질로써 이용가능하며, 이석의 윤문을 연륜으로 사용할 수 있는 것으로 판단된다.
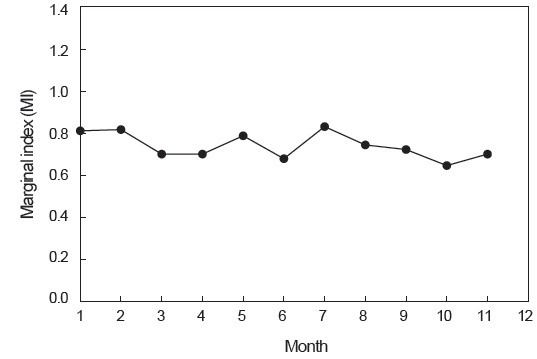
윤문의 형성시기 및 연간 형성횟수를 파악하기 위해 분석한 연역지수(MI)의 월 변화를 살펴보면, 7월에 가장 높은 값을 나타내었고, 10월에 가장 낮은 값을 나타내었으나, 연역지수 값의 범위가 0.649-0.831로 월별로 큰 차이를 보이지 않았다(Fig. 6).
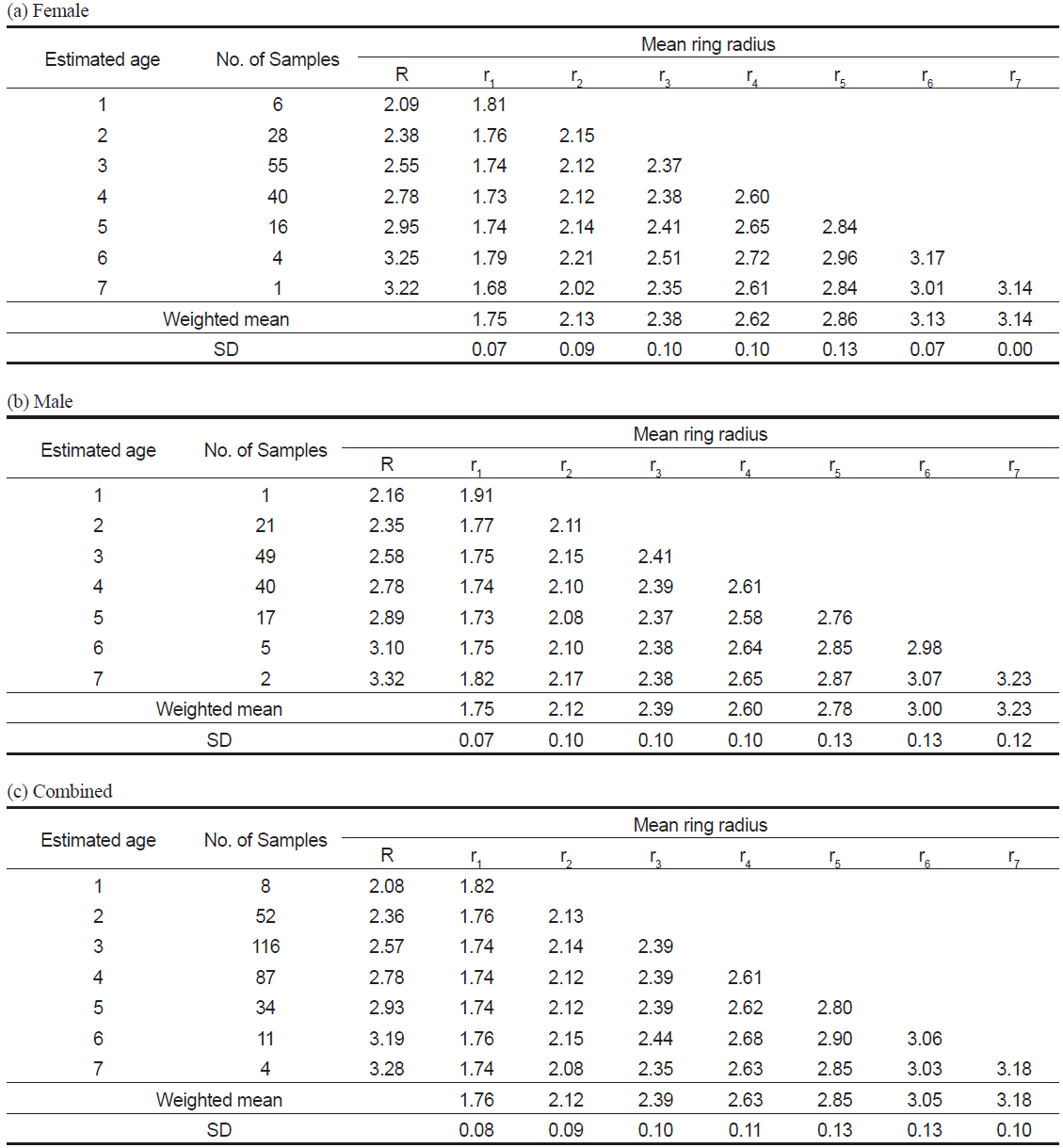
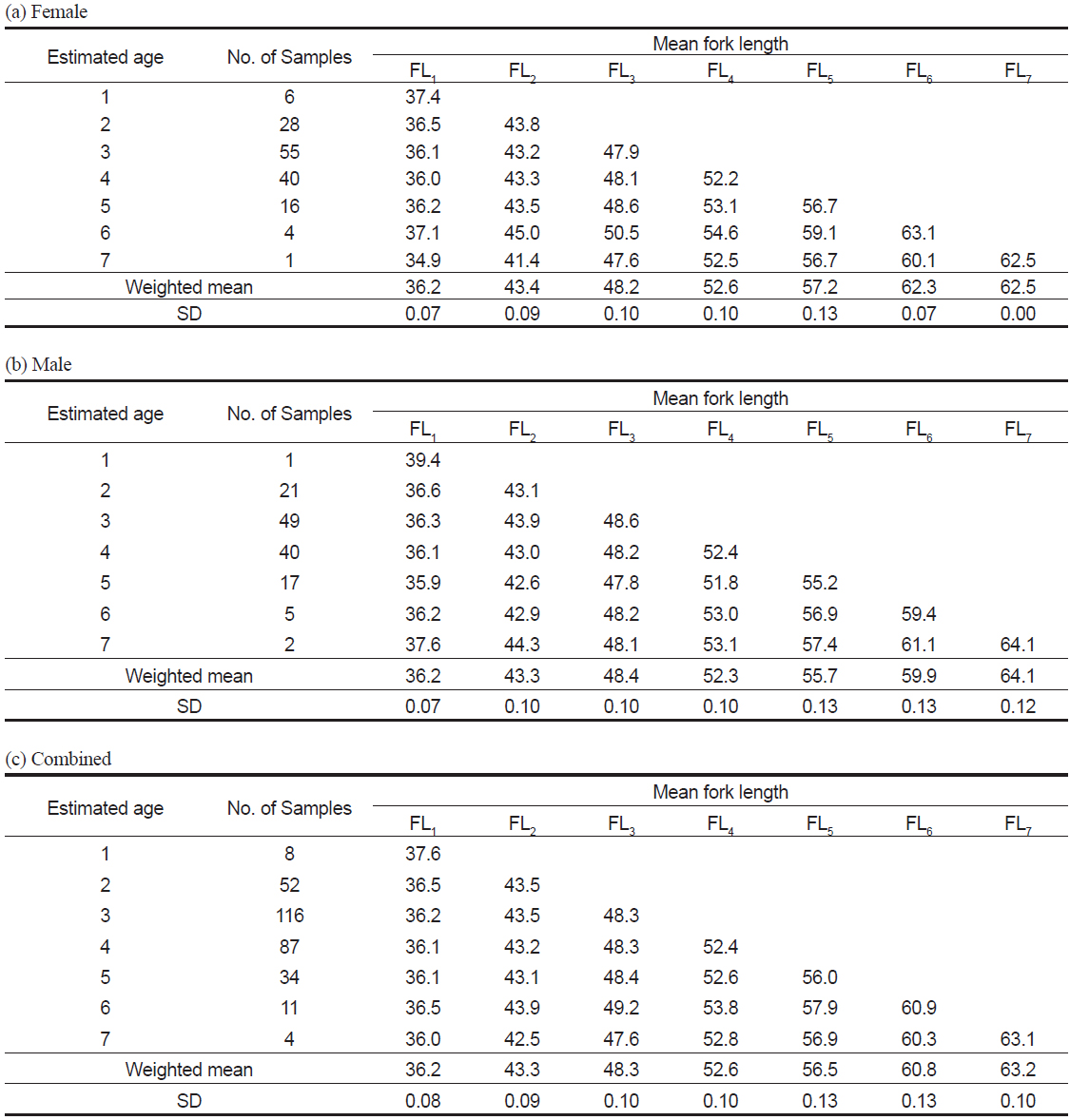
관측된 표본의 연령은 1세에서 7세로, 3세가 116 개체로 가장 많았다(Table 3c). 암∙수 전체에 대한 윤문별 평균윤경은 r1=1.76 mm에서 r7=3.18 mm로, 핵에서부터 r1까지가 가장 큰 증가폭을 보였고, 이후 점차 감소하는 경향을 보였으며(Table 3c), 암수별로도 동일한 경향을 보였다(Table 3a-3b).
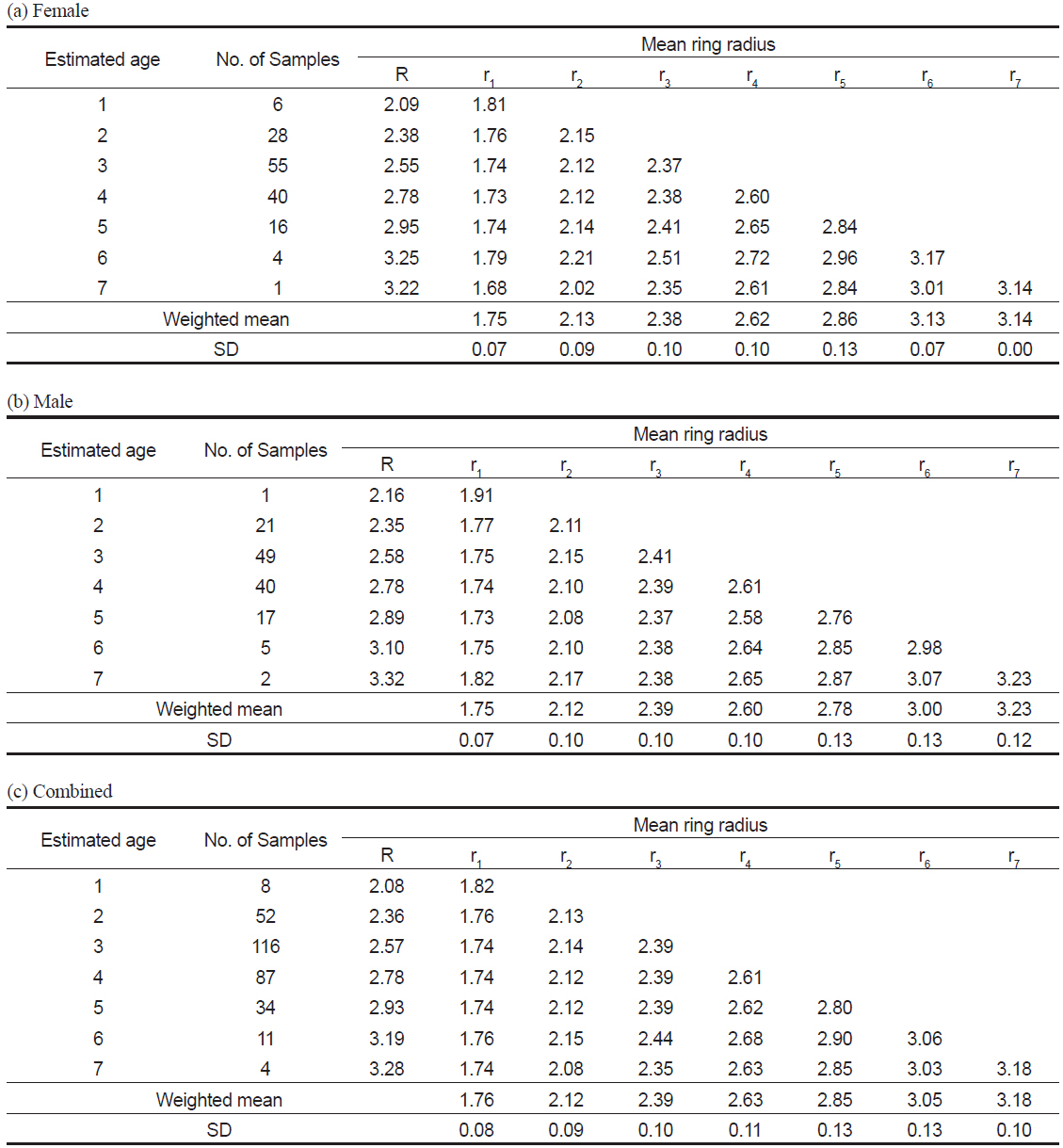
Mean ring radius (mm) when each annulus was formed in the otolith of Katsuwonus pelamis collected in the Western and Central Pacific Ocean
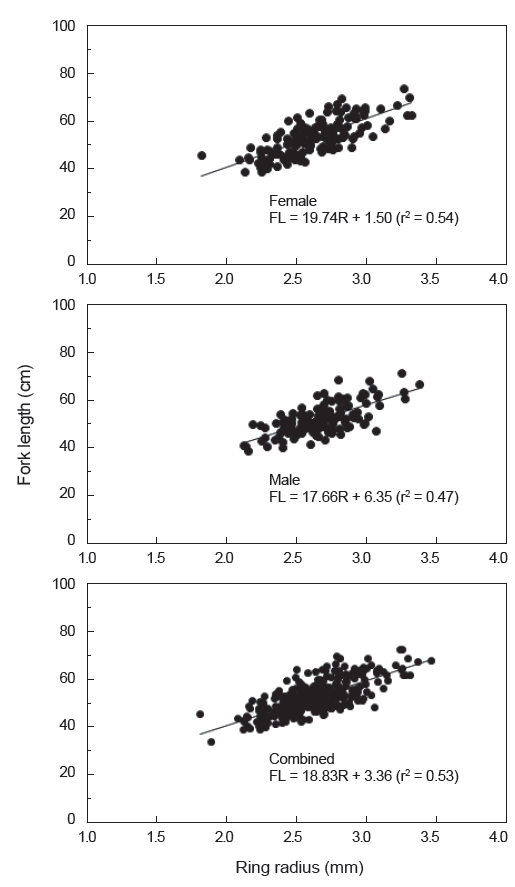
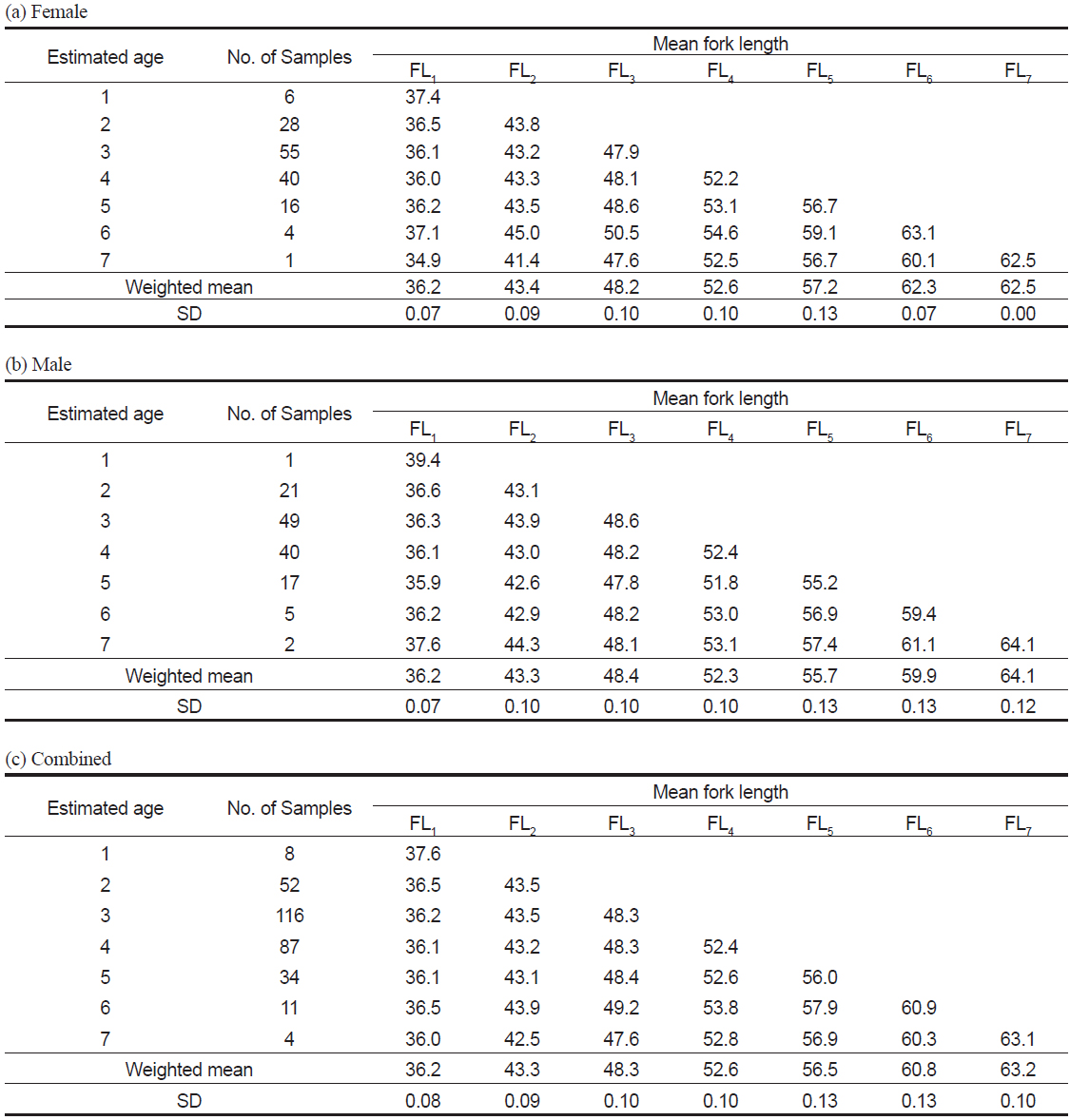
이석반경(R)과 가랑이체장(FL)의 관계식은 암컷은 FL = 19.74R+1.50 (r2=0.54), 수컷은 FL=17.66R+6.35 (r2=0.47)로 나타났으며, 암·수 전체의 관계식은 FL=18.83R+3.36 (r2=0.53) 이었다(Fig. 7). 따라서 이석반경과 체장간의 관계, 그리고 평균 윤경을 사용하여 역계산된 연령별 평균 가랑이체장은 암컷은 FL1=36.2 cm에서 FL7=62.5 cm, 수컷은 FL1=36.2 cm에서 FL7=64.1 cm로 나타났고, 암·수 전체의 평균 가랑이체장은 FL1=36.2 cm에서 FL7=63.2 cm이었다(Table 4).
Back-calculated fork length (cm) corresponding to the age when annulus was formed in the otolith of Katsuwonus pelamis collected in the Western and Central Pacific Ocean
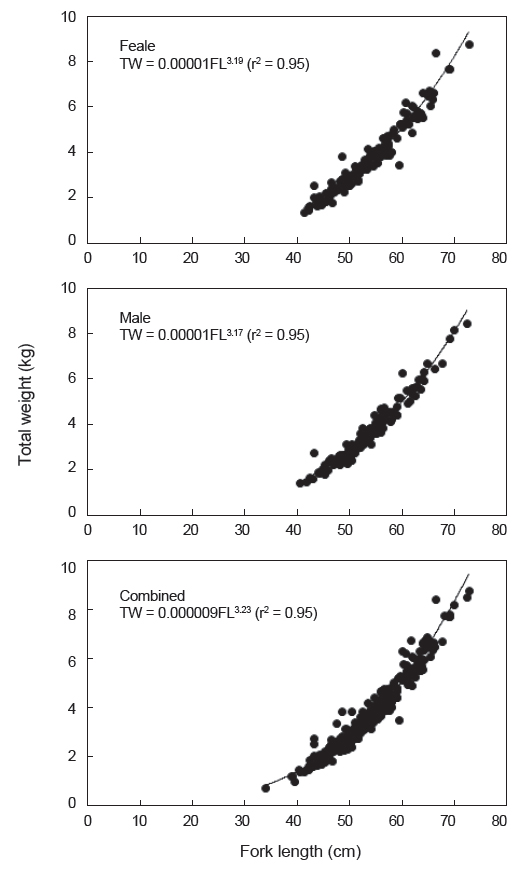
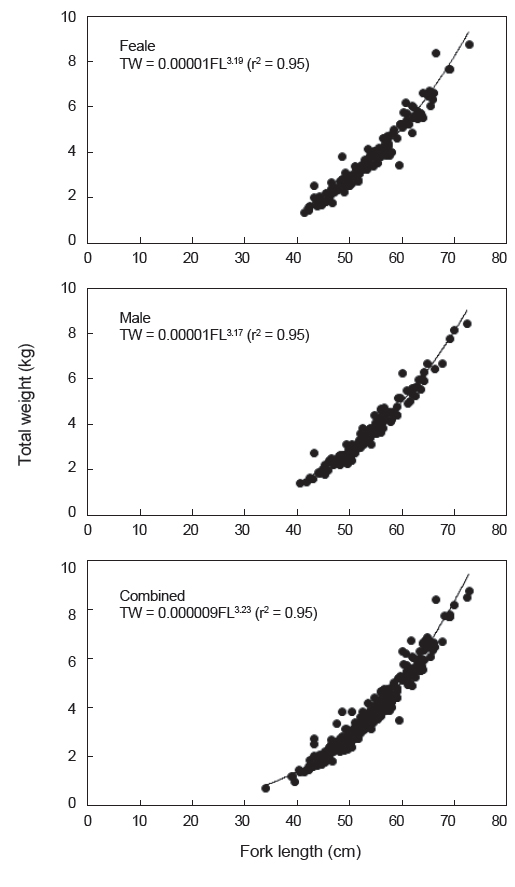
가랑이체장(FL)과 체중(TW)간의 상대성장식은 암컷은 TW=0.00001FL3.19(r2=0.95), 수컷은 TW=0.00001FL3.17(r2=0.95)이었으며, 암·수 전체에 대한 상대성장식은 TW=0.000009FL3.23(r2=0.95)이었다(Fig. 8).
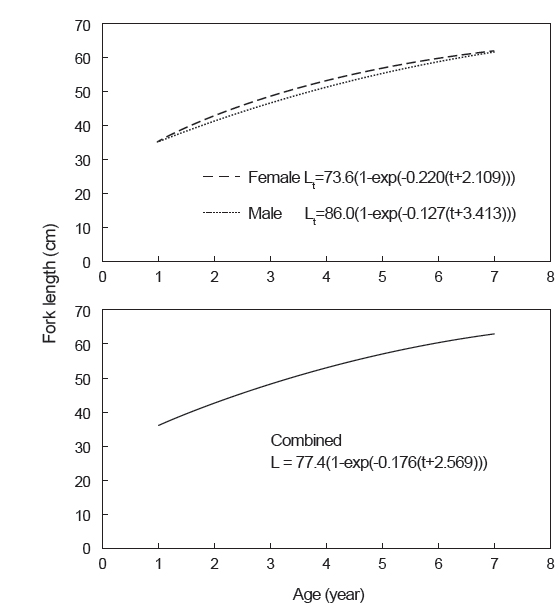
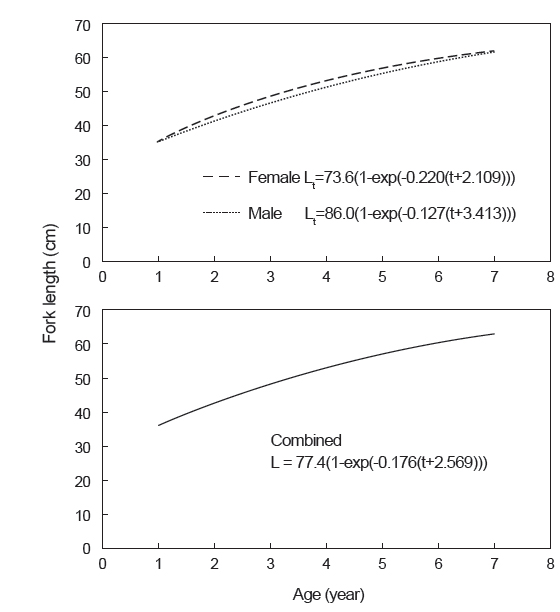
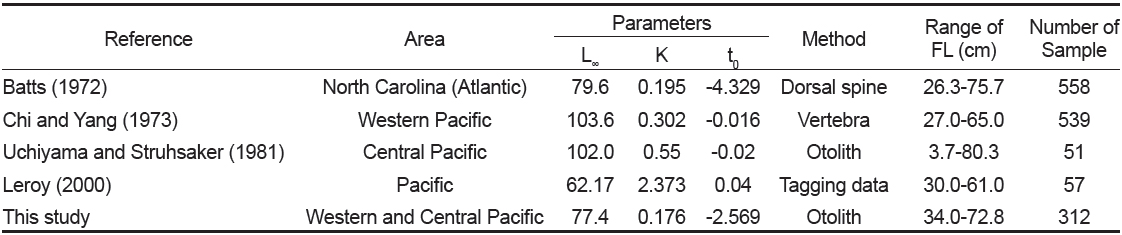
따라서 연령별 평균 가랑이체장으로부터 추정된 가다랑어의 von Bertalanffy 성장식은 Fig. 9와 같다. 암·수 전체에 대한 성장 매개변수는 이론적 최대 가랑이체장(L∞)이 77.4 cm, 성장계수(K)는 0.176/year, 체장이 0일 때의 이론적 연령(t0)은 −2.569 years이었으며, 암수간에는 큰 차이가 없었다. 그리고 이론적 최대체중(W∞)은 11.4 kg으로 추정되었다.
본 연구에서는 중서부태평양 수역의 가다랑어에 대한 연령과 성장을 구명하고자 연령형질로써 이석을 사용하여 분석하였다. 가다랑어의 연령과 성장에 관한 선행 연구결과를 살펴보면, Shabotiniets (1968)은 연령형질로 7 개체의 척추골과 등지느러미의 단면을 사용하여 3세는 40-45 cm, 4세는 40-60 cm라고 보고하였는데, 특히 4세의 가랑이체장 범위가 넓은 분포를 나타내었다. 그리고 Aikawa and Kato (1938), Yokota et al. (1961)은 연령형질로 척추골을 사용하였는데, Aikawa and Kato (1938)은 1세의 가랑이체장이 27-37 cm, 4세가 55-64 cm, 그리고 Yokota et al. (1961)은 1세가 37 cm, 5세는 85 cm라고 보고하여, 1세는 이석을 이용한 본 연구결과와 유사하였으나 4-5세는 척추골에 의해 추정된 가랑이체장이 크고, Shabotiniets (1968)과 마찬가지로 넓은 체장 범위를 보였다.
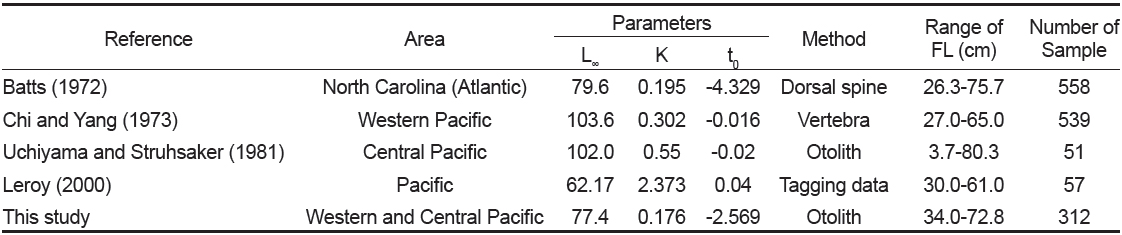
본 연구에서 추정된 von Bertalanffy 성장 매개변수는 이론적 최대체장(L∞)이 77.4 cm, 성장계수(K)는 0.176/year, 체장이 0일 때의 이론적 연령(t0)은 −2.569 years이었다. Batts (1972)는 등지느러미의 단면을 사용하여 L∞는 79.6 cm, K는 0.195/year, t0는 −4.329 years로 추정하였고, Chi and Yang (1973)은 척추골을 사용하여 L∞는 103.6 cm, K는 0.302/year, t0는 −0.016 years, Uchiyama and Struhsaker (1981)는 L∞는 102.0 cm, K는 0.55/year, t0는 −0.02 years, Leroy (2000)는 표지방류 자료를 사용하여 L∞는 62.2 cm, K는 2.373/year, t0는 0.04 years로 추정하였다(Table 5). 따라서 이석을 이용한 본 연구 결과는 등지느러미의 단면을 이용한 Batts (1972)의 결과와 유사하였으나, 동일한 이석을 이용한 Uchiyama and Struhsaker (1981)의 결과와는 차이를 보였다. 그러나 본 연구에서 사용된 표본은 상업적 어선인 선망어업에 의해 수집되었기 때문에, 상업적 가치가 없는 소형개체가 채집되지 않았고, Uchiyama and Struhsaker (1981)는 분석에 사용된 표본수가 너무 적다는 문제점을 안고 있다.
Comparison of the von Bertalanffy’s growth parameters of Katsuwonus pelamis estimated in this study with those of previous studies
연역지수(MI)는 일반적으로 어류의 윤문형성시기를 판단하는 지표로 이용되고 윤문형성시기는 산란시기와 관련이 있다(Zhang et al., 2010). 본 연구 결과에 의하면, 가다랑어 연역지수의 월 변화가 뚜렷한 경향을 나타내지 않았다. 그 원인은 일반적으로 가다랑어는 열대(적도) 해역에서는 특정 시기와 무관하게 연중 산란하고, 아열대 해역에서는 주로 봄부터 가을 사이에 산란하는 것으로 알려져 있는데(Collette and Nauen, 1983;Matsumoto et al., 1984) 이러한 산란 특성과 관련이 있는 것으로 보인다. 가다랑어의 산란에 대해 좀 더 살펴보면, 동부태평양의 경우 15°N-20°S 해역에서는 10월에서 3월 사이에 작은 성어들이 산란을 하고(Naganuma, 1979), 60 cm 이상 되는 큰 성어들은 15°N의 이북에서 산란을 한다고 한다(Shomura, 1966). 한편, 하와이 해역에서는 4월말에서 5월초, 그리고 7월말에서 8월초 사이에 1년에 최소 2번은 산란한다고 하였으며(Matsumoto et al., 1984), 남태평양에서는 11월에서 12월 사이에 산란이 일어나고, 산란기간 동안 반복적으로 산란이 일어날 가능성이 있다고 한다(Hunter et al., 1986). 이렇듯 가다랑어는 서식지에 따라 산란시기가 다르고, Fujino et al. (1981)은 가다랑어는 4개의 계군(인도양, 대서양, 서부태평양, 중동부 태평양)이 존재한다고 하였다. 따라서 본 연구를 수행한 해역에는 두 개 이상의 계군이 존재할 가능성이 있으며, 연령과 성장을 포함한 가다랑어의 명확한 생태학적 특성 구명을 위해서는 향후 계군분석 연구가 수행되어야 할 것으로 생각된다.
본 연구에서는 이석을 이용하여 가다랑어의 연령과 성장을 분석하였으며, 이러한 자료는 가다랑어의 자원생태학적 특성치 추정 및 자원평가에 중요한 정보로 활용될 수 있을 것이다. 그러나 앞에서 언급한 바와 같이, 가다랑어의 연령과 성장에 있어서 연구자들간에 차이를 보이고 있어, 이들의 생태학적 특성에 대한 정확한 구명을 위해서는 계군분석과 각 계군별 산란 및 회유 특성에 관한 체계적인 조사·연구가 필요한데, 가다랑어는 고도 회유성 어종이므로 여러 기관(또는 국가)간의 협업을 통해 수행하는 것이 보다 효율적일 것으로 생각된다.