Proteomics aims at the analysis of all the protein content of a biological system1. This involves the identification and study of protein structure, functions, post-translational modifications and interactions.2 Post-translational modifications of proteins are vital in biological functions. Phosphorylation plays a key role in cell signaling,3 for instance, while glycosylation has important roles in molecular transport.4 Phosphorylation occurs on 30% of mammalian proteins,5 while glycosylation occurs on 50% of proteins.6 Aberrant phosphorylation has been found to be responsible for signaling abnormalities in various cancers,7 while glycosylation is found to play roles in cancer metastasis.8 Understanding these changes through proteomic studies may play a role in biomarker discovery. However, biological samples are complex and highly diverse protein mixtures. Protein expression is a system which is dynamic, dependent on regulation and environmental signaling. While MS can identify the peptides generated by digestion of the proteins, it can only identify a limited number of peptides in a period of time and is also limited by ion suppression effects in complex mixtures.9 For accurate analysis and quantification it is important to separate peptides well using chromatography before MS analysis. Enrichment for peptides with posttranslational modifications may also increase their rates of identification.
The physiochemical properties of amino acid residues vary. Chromatography can separate peptides by exploiting differences in charge, as in ion exchange chromatography; in polarity, as in hydrophilic interaction chromatography (HILIC); in hydrophobicity, by RPLC; and in size, using size exclusion chromatography. Chromatographic separation plays an important role in both the top-down and bottom-up proteomic approaches. In the top-down approach, liquid chromatography methods are used to separate whole proteins before MS analysis.10 Liquid chromatography serves as a better alternative to twodimensional gel electrophoresis,11 a traditional method that is limited by issues such as poor reproducibility, a laborintensive workflow and a narrow dynamic range.12 The bottom-up approach, in contrast, involves the enzymatic digestion of proteins into peptide fragments before chromatographic separation and MS analysis.13 By reducing the sample complexity, chromatography increases the probability of detecting peptides of low abundance whose identification is hindered by the ion suppression effects from peptides of high abundance that compete for ionization. This results in the under-detection of peptides of low abundance. Multidimensional liquid chromatography (MDLC) methods have been developed in order to improve separation and resolution.14 Orthogonality is achieved by utilizing methods that separate peptides by different physicochemical properties.15 This in turn improves peak capacity and resolution.14
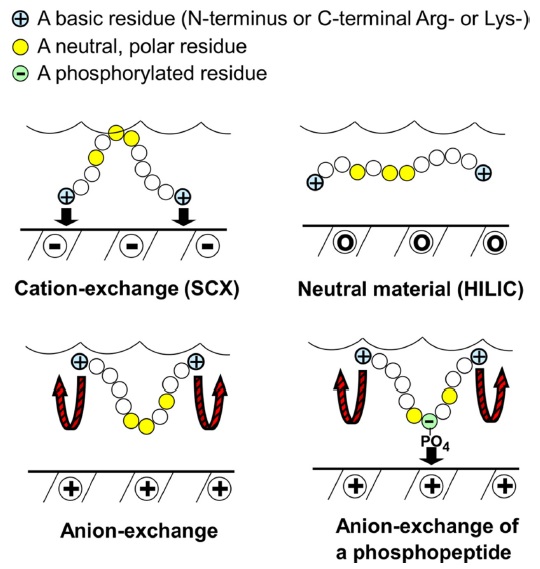
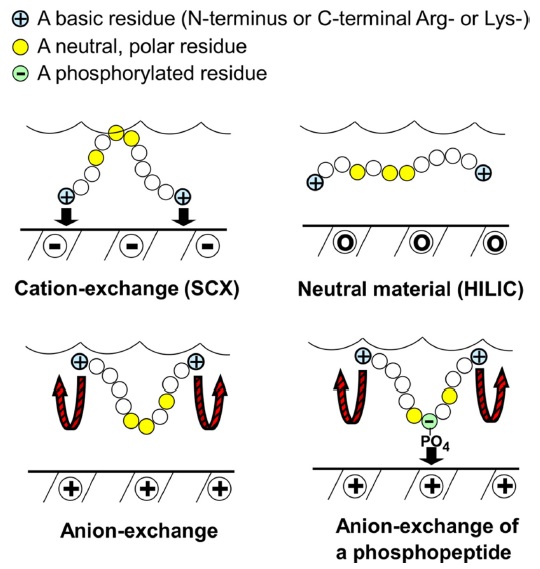
The mechanism for HILIC is hypothesized to involve a stagnant layer of water interacting with the polar stationary phase, with the analyte partitioning between it and the dynamic mobile phase. In HILIC, a neutral stationary phase is used and peptides are separated based on hydrophilic interaction. The more polar an analyte, the greater its partitioning into the stagnant aqueous phase and the longer the retention time (Figure 1). In order to elute the analytes, a gradient of decreasing concentrations of organic solvent is used in order to make the mobile phase more polar. The use of HILIC requires the use of a gradient to separate basic analytes, because of their strong hydrophilic interaction.16
As a recently introduced mixed-mode chromatographic method, ERLIC combines hydrophilic interaction and electrostatic repulsion. The two different forces hydrophilic interaction and electrostatic repulsion will act during separation. Peptides of the same net charge as the stationary phase will still be retained if hydrophilic interaction is strong enough. In the case of peptides, this usually involves an anion-exchange column (Figure 1).A high concentration of organic solvent would allow hydrophilic interaction to retain compounds which are of the same net charge as the stationary phase because electrostatic repulsion and hydrophilic interaction are independent properties. Electrostatic repulsion may also be harnessed to repel strongly retained compounds.17 The ERLIC combination is shown to resolve certain issues associated with peptide retention. For very acidic peptides that elute late as a result of strong electrostatic attraction to the column, a low pH may be used to uncharge aspartate and glutamate residues, leaving them neutral. For very basic peptides that elute late because of strong hydrophilic interaction, the use of electrostatic repulsion would allow isocratic elution of peptides and peptides differing widely in charge within a similar time frame, with a mechanism being likened to that of an immobilized salt gradient. For basic peptides that elute early because of electrostatic repulsion, hydrophilic interaction can be used to increase their retention by using a high concentration of organic solvent.17
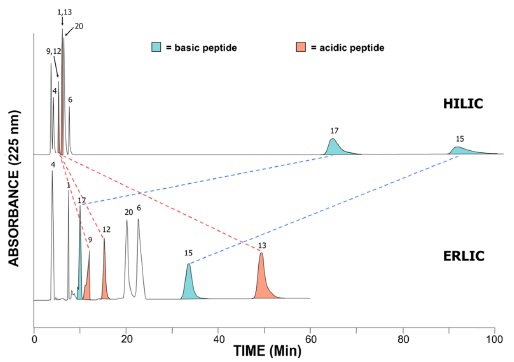
Hence, the ERLIC method allows the isocratic resolution of peptide mixtures that would require gradients in other methods, such as HILIC (Figure 2). This is because the basic residues which bind strongly in HILIC would have their retention In HILIC, basic peptides are retained the longest and there is poor isocratic separation of earlyeluting peptides. ERLIC affords resolution within 60 minutes of these peptide standards (Figure 2). Selectively antagonized by utilizing the combination of an anionexchange column and hydrophilic interaction,17 ERLIC also allows for the enrichment of phosphopeptides. At pH 2, phosphates retain negative charge, while the carboxylgroups of aspartate and glutamate remain uncharged. Peptides with one or more phosphate groups will then be selectively retained through a combination of the electrostatic attraction by the phosphate groups and their appreciable hydrophilic interaction. Unmodified peptides with basic residues (cf. the N-terminus and the C-terminal Arg or Lys of tryptic peptides) will experience only electrostatic repulsion and elute in the void volume if the hydrophilic interaction is not too high. A salt gradient is required when more than one phosphate group is present.17
ERLIC was originally introduced by Alpert to separate negatively-charged analytes such as nucleotides and to enrich acidic peptides such as phosphopeptides and sialic acid-containing glycopeptides.17 Hao
This review aims to provide an overview of the uses of ERLIC for proteomics applications. The enrichment of phosphopeptides is demonstrated, as is the analysis of other post-translational modifications. The fractionation by ERLIC of tryptic peptides in general is shown to be suitable for the first dimension of MDLC-MS/MS. In addition, ERLIC can be used to study peptide deamidation, which is difficult to accomplish with RPLC (Table 1).

The use of Electrostatic Repulsion-Hydrophilic Interaction Chromatography (ERLIC) for Proteomic applications
>
General considerations in ERLIC
For tryptic peptides, an anion-exchange column such as PolyWAX LP or an amino column has been used successfully. At pH 2.0, peptides with a strongly acidic group such as phosphopeptides are the best retained. As pH increases, carboxyl groups become ionized and acidic peptides become better retained, up to a maximum pH of 5.0. Above that pH, the charge density of a WAX (weak anion-exchange) column decreases.17 In a range of 65-70% ACN concentration, electrostatic repulsion is stronger than hydrophilic interaction for typical tryptic peptides, most of which flow through the column unretained or elute within a short and similar timeframe, but phosphopeptides elute later. Beyond 70%, unmodified peptides become retained because of the increase in hydrophilic interaction. The use of Na-MePO3 promotes the selective retention of phosphopeptides and has been used in the investigation of ERLIC for phosphopeptide enrichment and separation. As Na-MePO3 concentration increases, the retention of basic peptides increases while retention of acidic peptides decreases, because the presence of salt blocks electrostatic effects, both attractive and repulsive. This implies that the appropriate combination of ACN concentration and salt concentration could be used to elute peptides within an appropriate timeframe. Elution of multi-phosphorylated peptides requires a gradient of increasing salt and decreasing ACN concentration. TEAP was chosen for such gradients both because of its good solubility at high levels of ACN and because it promotes retention of phosphopeptides less well than does Na-MePO3. When TEAP was used, basic peptides were well retained while acidic ones were not. This was presumably because the presence of a second negative charge in the phosphate counterion allowed basic residues to be attracted and retained, while acidic amino acids are repelled. Using a concentration low enough to reduce the shielding effects of the salt may allow the acidic amino acids to elute later and basic amino acids to elute earlier, and hence within the same time frame. However, when methylphosphonate is used as the counterion, only a single negative charge is present, which allows acidic peptides to bind to the column.17
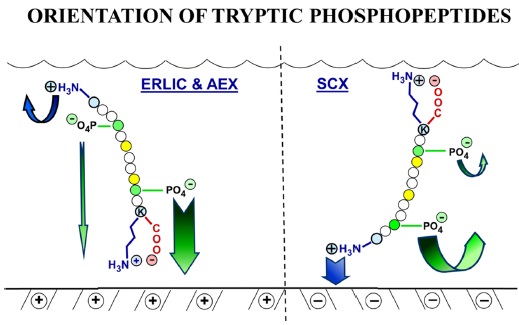
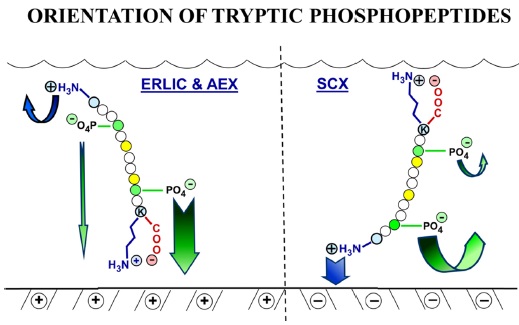
The selectivity of ion exchange chromatography is affected by the orientation of peptides. It was discovered that when ERLIC is performed using a weak anion exchange column, tryptic peptides are oriented such that the C-terminus faces the stationary phase. This could be because a zwitterion can form between the C-terminal carboxyl group and the C-terminal Arg or Lys residue, resulting in less electrostatic repulsion from the column than is experienced by the N-terminus (Figure 3). The position of phosphate groups in the peptide has the potential to affect retention time. When a phosphate group is closer to the C-terminal end, the retention time of a tryptic peptide would be longer on a weak anion exchange column than would be the case if the phosphate group is farther away. When peptides have more than one phosphate group, then the net charge seems to determine the peptide retention.23
ERLIC for Tryptic peptide fractionation
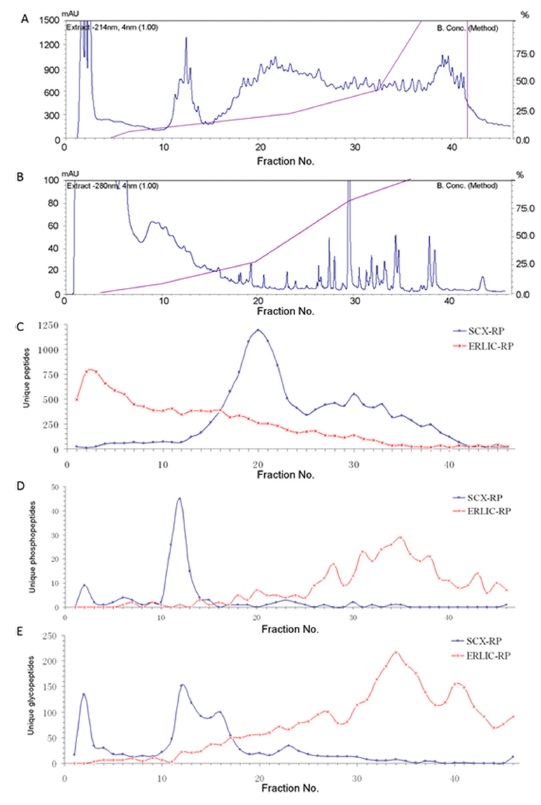
ERLIC-RPLC was evaluated in comparison to SCXRPLC, a common MDLC combination, using a tryptic digest of rat kidney tissue. The first dimensional fractionation was performed in a 4.6 mm × 200 mm PolyWAX and PolySULFOETHYL columns. For ERLIC fractionation, a gradient was used starting with a high concentration of ACN followed by a decrease in organic solvent and pH: 100% buffer A (90% ACN/0.1% acetic acid, pH 3.6) for 10 min, 0-8% buffer B (30% ACN/0.1% formic acid [FA], pH 3.0) for 20 min, 8-27% buffer B for 30 min, 27-45% buffer B for 10 min, 45-81% buffer B for 20 min, and 81-100% buffer B for 20 min followed by 30 min at 100% buffer B at a flow rate of 0.5 mL/min.
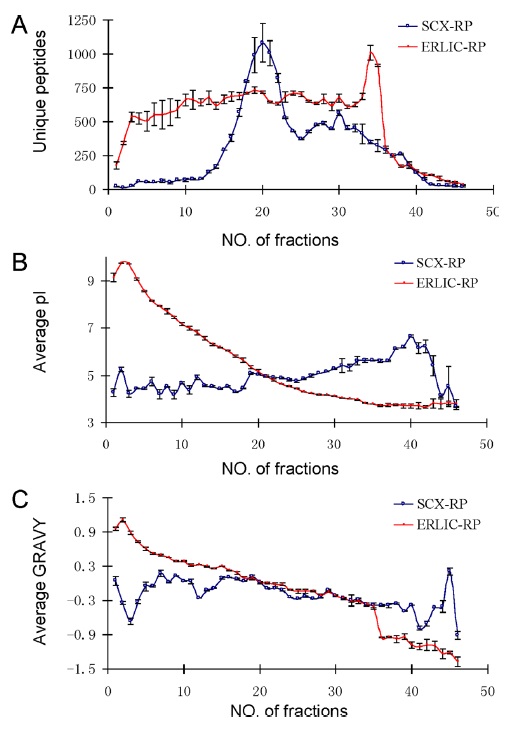
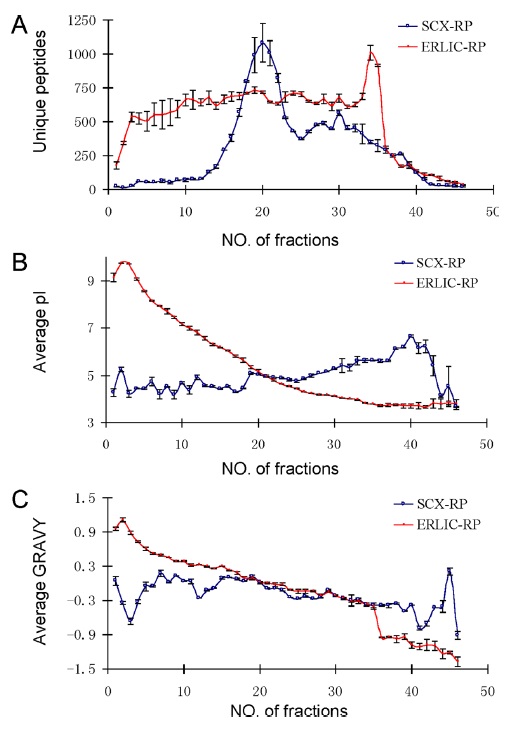
Figure 4 compares the separation of peptides using ERLIC-RPLC and SCX-RPLC. The number of unique peptides in each fraction was found to be distributed much more evenly in ERLIC than in SCX. In addition, the separation of peptides in ERLIC is based on decreasing isoelectric point and decreasing average GRAVY values. (increasing polarity). GRAVY value is an indicator of the hydrophobicity of a peptide, and hence describes polarity of the peptide. No such trend was observed for SCX chromatography. The ERLIC-RPLC method identified 36% more proteins and 64% more unique peptides compared to SCX-RP. Moreover, the ERLIC method was able to identify 120% more basic and hydrophobic peptides than SCX. This may be applicable to identification of peptides in membrane-rich organelles.18
ERLIC was subsequently evaluated in comparison with SCX in the fractionation of iTRAQ- labeled peptides in rat liver tissue.19 It was observed that iTRAQ-labelled peptides had decreased solubility in 90% ACN/0.1% acetic acid, and decreasing ACN concentration to 70% did not completely solubilize them. In this protocol, volatile ammonium salts and a lower ACN concentration had to be used in order to improve solubility, hence the choice of 10 mM NH4-formate in 85% ACN/1% FA as mobile phase A. A desalting step was not required but vacuum drying had to be performed above 45℃ to eliminate the volatile ammonium salts. Again, ERLIC affords a more even distribution of peptides among the collected fractions, and it also separates iTRAQ-labelled peptides based on decreasing isoelectric point and increasing polarity. This trend is not observed in SCX fractionation. This observation is in agreement with Hao
ERLIC has been evaluated in comparison with other methods. A study was done to evaluate fractionation for phosphopeptides in HeLa cells using HILIC, SCX and ERLIC with volatile (NH4-formate) and non-volatile (NaMePO3 and TEAP) salts following IMAC enrichment for phosphopeptides. This was followed by RPLC-MS/MS.24 The four methods together allowed the identification of 9069 unique phosphopeptides. The four sets of unique phosphopeptides exhibited little overlap; each method identified phosphopeptides not identified by the others. This suggests the possibility of combining methods to maximize the identification of phosphopeptides. The SCX method demonstrated good orthogonality to RPLC and provided good resolution in phosphopeptide separation. However, it did not perform as well compared to ERLIC methods in the separation between monophosphorylated peptides and multiphosphorylated peptides, and the workflow required a desalting step. HILIC demonstrated lesser resolution and orthogonality to RP, but the workflow did not require the desalting step. HILIC also had the lowest number of identifications of diphosphopeptides. ERLIC with the non-volatile buffers was the best in separation of multiphosphorylated and monophosphorylated peptides, in agreement with the principles of the ERLIC method. It also demonstrated good orthogonality to RP. However, there was worse resolution in phosphopeptide separation and the fractions required desalting. ERLIC with volatile buffers did not require a desalting step.24
de Jong
The use of SCX, ERLIC, RPLC at pH 2.5 with a C-18 material, RPLC at pH 9.7 with a C-18 material, and a pentafluorophenylpropyl (F5) column (elution similar to low-pH RPLC) were evaluated as alternatives for the first dimension of tryptic peptide fractionation of human serum proteins.20 The second dimension involved a nanoChip-LC (RP C18, pH 2.6) and Q TOF-MS/MS. The highest number of peptides was identified by ERLIC, while an equal number of proteins were identified by ERLIC and high-pH RPLC. The orthogonality of the different fractionation methods was evaluated using Pearson correlation. An ERLIC- high pH RP- low pH nanoChip-LC-MS/MS sequence was implemented for the profiling of serum proteins, and this 3-dimensional method identified 208 proteins and 1088 peptides.20
Following validation of ERLIC for use on iTRAQ labelled proteins,19 several proteomic studies have been done. The use of ERLIC for iTRAQ labelled peptides has been applied to identify the components of proteins secreted by
ERLIC for Post-Translational Modification Analysis
>
ERLIC for the enrichment of phosphopeptides
ERLIC was evaluated in comparison to SCX-IMAC in the enrichment of phosphopeptides. The gradient used was 10 mM sodium methylphosphonate (Na-MePO3) with 70% acetonitrile, pH 2.0 (Solvent A) and 200 mM triethylamine phosphate (TEAP) with 60% acetonitrile, pH 2.0 (Solvent B). This involved decreasing the ACN concentration and replacing Na-MePO3 with TEAP as a salt to elute phosphopeptides. ERLIC was found to enrich for more total phosphorylation sites and more multi-phosphorylated peptides than SCX-IMAC in EGF-treated A431 cells. SCX-IMAC enriched for more unique phosphopeptides and mono-phosphorylated peptides than did ERLIC. Motif analysis showed that ERLIC enriched for more acidophilic motifs while SCX-IMAC enriched for more basophilic ones. The ERLIC method did not detect any basophilic motifs. The ERLIC workflow is also more efficient, utilizing a single step as compared to SCX-IMAC, and hence has shorter processing time. Each approach could not adequately cover the entire phosphoproteome. There was little overlap (12%) between the two phosphopeptide sets in the number of unique phosphopeptides identified by both methods, suggesting that they could be used in combination to increase phosphoproteome coverage.30
ERLIC and IMAC were used to study phosphorylation changes in chicken embryonic fibroblast peptides that were infected with Marek’s Disease virus (MDV). This approach identified approximately 50 times more unique phosphopeptides than did ERLIC or IMAC alone. 54.6% of unique total peptides identified were unique phosphopeptides. The ERLIC method alone identified the most unique total peptides, but only 1.2% of the total peptides were unique phosphopeptides. This demonstrates the importance of the IMAC step for further enrichment. These methods resulted in 581 unique phosphopeptides being detected in MDV-infected and mock-infected cells. Among 279 unique phosphopeptides in MDV- infected cells, 32 phosphopeptides were associated with the MDV MD11/75C virus. In contrast, 150 phosphopeptides were detected that were unique to mockinfected cells.31
Proteomic approaches have been applied to study the temporal regulation of the DNA damage response.32 Phosphopeptide enrichment was performed using ERLIC and TiO2 affinity chromatography on SILAC-labelled cells, followed by LTQ-Orbitrap MS to identify and classify double strand break-induced phosphorylations. 5204 phosphorylation sites were identified, of which 594 sites in 204 proteins were found to have their abundance altered by more than 2-fold. This study identified novel phosphorylation sites that are regulated by DNA damage and provided greater understanding of temporal signaling events following DNA damage.32
Guo
>
ERLIC for the enrichment of glycopeptides
ERLIC was used for the enrichment of glycopeptides for platelet glycoproteins. Human platelets were selectively purified and plasma membranes enriched using aqueous two-phase partitioning with PEG and dextran. ERLIC was then performed using a PolyWAX LP column. The gradient involved a buffer A containing 20 mM methylphosphonic acid adjusted to pH 2.0 using NaOH in 70% acetonitrile and buffer B containing 200 mM triethylamine phosphate in 60% acetonitrile. The conditions involved are the same as those used for phosphopeptide enrichment. Subsequently, PNGase F treatment and MS were performed. Peptides with deamidation at Asn residues were presumed to have been N-linked glycopeptides. The mechanism for ERLIC enrichment is speculated to involve electrostatic attraction with sialic acid residues (which retain some charge at pH 2.0) plus hydrophilic interaction. Peak broadening was observed in the elution of glycopeptides, which could indicate glycan isoform separation. This was subsequently verified by the authors as real and not artifactual. 125 glycosylation sites on 66 proteins were discovered, providing an increase in the existing knowledge of glycosylation sites on platelets.34
In one study, Jacalin lectin affinity chromatography was used to isolate glycoproteins with O-linked glycans from bovine serum, followed by tryptic digestion and ERLIC chromatography. The O-linked glycopeptides in the ERLIC fractions were then isolated with the Jacalin column, followed by partial deglycosylation with neuraminidase and beta-galactosidase, after which higher-energy collisiondissociation (HCD) and electron-transfer dissociation (ETD) MS analyses were performed. Alternatively, the intact glycoproteins were fractionated via mixed-bed ionexchange, followed by digestion and Jacalin enrichment of the O-linked glycopeptides. ERLIC fractionation without subsequent affinity enrichment did not adequately select for sialylated glycopeptides. Enrichment was considered successful since 65% of the peptides identified in the protein level approach and 76% of peptides in the peptide level approach were glycopeptides. Both methods combined yielded a total 126 O-glycosylation sites in 51 different proteins, with 76 novel O-glycosylation sites and 35 newly identified proteins. The ERLIC and affinity purification approach yielded slightly more glycoproteins and glycosylated sites compared to the mixed bed IEX chromatography. The study was limited to the selection of GalNAcá1 peptides without C6 modification through the use of Jacalin, as well as by the loss of structural information as a result of the partial deglycosylation steps.35
ERLIC for the simultaneous categorization of post-translational modifications
There are several potential advantages in the simultaneous categorization of post-translational modifications (PTMs). Both phosphorylation and glycosylation may occur as a result of a single signaling event, and being able to analyze this provides important information. It has been noted that cross-talk between signaling from different PTMs may regulate cellular processes.36 The simultaneous categorization of PTMs also avoids bias introduced by inter-experimental variation.
ERLIC has been used to simultaneously characterize the phosphoproteome and glycoproteome of mouse brain membrane. A comparison was also done between ERLIC and the hydrazide chemistry method in the enrichment of glycopeptides. Using ERLIC, 544 glycoproteins and 922 glycosylation sites were identified. In contrast, the hydrazide chemistry method only identified 192 glycoproteins and 345 glycosylation sites. 272 glycosylation sites and 174 glycoproteins could be identified with both methods. The combined results from both methods led to the identification of 995 glycosylation sites and 562 glycoproteins. 96% of the glycosylation sites were considered new. Also, 13.7% of the glycoproteins was found to be associated with neurological disorders. In addition, the ERLIC method allowed the identification of 383 phosphoproteins and 915 phosphorylation sites. Phosphopeptides were found to elute earlier while glycopeptides eluted later and over a wider range of organic solvent, suggesting successful fractionation and enrichment of the two categories of post-translational modification.37 The gradient was chosen accordingly, from buffer A (10 mM Na-MePO3, pH 2.0, with 70% acetonitrile) to buffer B (200 mM TEAP, pH 2.0, with 25% acetonitrile).
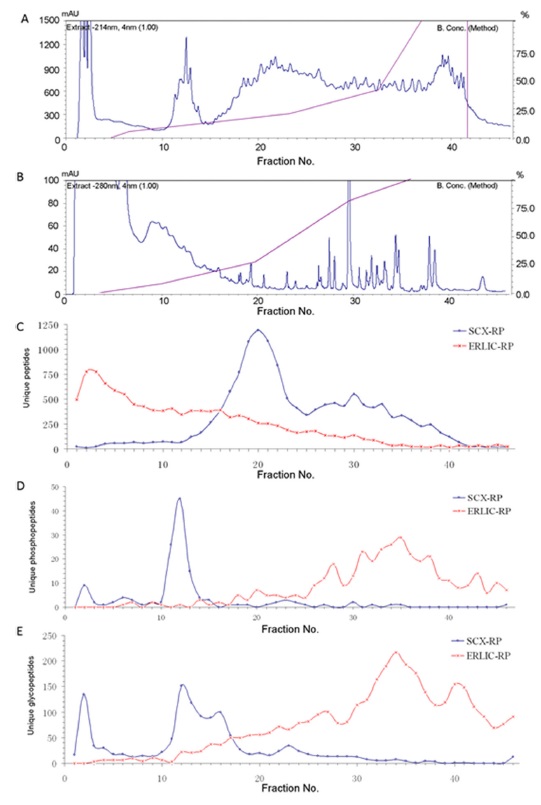
ERLIC has allowed the simultaneous characterization of the proteome, phosphoproteome and glycoproteome in rat kidney tissue in a single run.38 This is done by facilitating the retention of unmodified peptides by the ERLIC column during the enrichment of phosphopeptides and glycopeptides. Different SCX and ERLIC gradients were evaluated, followed by PNGase F treatment and LC-MS/MS. In ERLIC, peptides can be retained by starting with a concentration of organic solvent high enough to cause a modest degree of retention of unmodified peptides despite their electrostatic repulsion by the column. This concentration is then decreased to decrease the hydrophilic interaction. Phosphopeptides and glycopeptides elute later than unmodified peptides because of their hydrophilic interaction and attraction to the column. In SCX, however, phosphopeptides and glycopeptides tend to elute early (Figure 5). It was found that an ERLIC method involving 80% ACN, 0.1% FA as buffer A and 10% ACN, 2% FA as buffer B resulted in the most identifications of proteins, phosphoproteins, phosphorylation sites, glycoproteins and glycosylation sites compared to other SCX and ERLIC gradients tested. The choice of 80% ACN was a compromise between the two protocols for good retention of all unmodified peptides and the enrichment for modified ones. The protocol by Zhang
Tran
>
ERLIC for the study of peptide deamidation
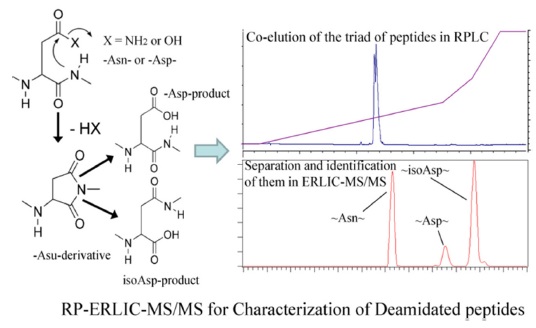
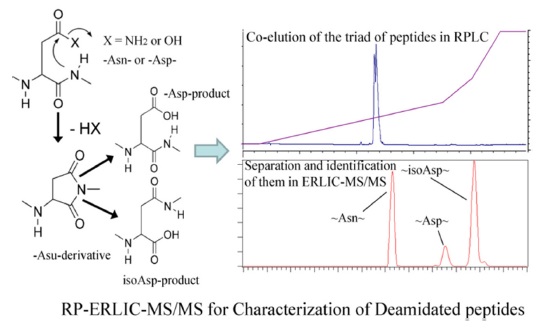
Some asparagine residues (N) can undergo deamidation nonenzymatically, forming a succinimide ring intermediate that hydrolyzes to produce isoaspartate (isoD) or
RPC-ERLIC can also be used to study peptide deamidation, either
This paper reviews the uses of ERLIC for proteomics applications. ERLIC has proved to be effective if not superior for tryptic peptide fractionation, and has been applied successfully with various biological samples such as serum, milk, and tissue extracts. Peptides are fractionated according to isoelectric point and polarity. It also allows the enrichment for phosphopeptides, and has also been utilized to enrich for other PTMs such as glycosylated and deamidated peptides. In addition, the simultaneous analysis of the proteome and post translational modifications has been achieved. ERLIC has been shown to perform as well or better than methods currently in widespread use for various applications.
ERLIC is expected to be utilized further in studies of PTMs in order to identify novel modified sites as potential biomarkers for diseases.39 The increase in hydrophobic and basic peptides identified by ERLIC-RP suggests potential applications for analyzing membrane proteins.18 ERLIC parameters may be further optimized for use with labelling strategies such as SILAC, ICAT and TMT to allow an increase in the sensitivity of quantification.19,31
![Comparison of HILIC and ERLIC in the resolution of peptide standards. (HILIC was performed using a PolyHYDROXYETHYL A column [5-μM, 300-A], with a mobile phase of 20 mM sodium methylphosphonate (Na-MePO3), pH 2.0, with 63% ACN. Flow rate: 1.0 mL/min. ERLIC was performed using a PolyWAX LP column [same dimensions], with the same mobile phase except containing 70% ACN. Flow rate: 1.0 mL/min.). Taken from Reference 17.](http://oak.go.kr/repository/journal/17350/E1MPSV_2014_v5n4_95_f002.jpg)