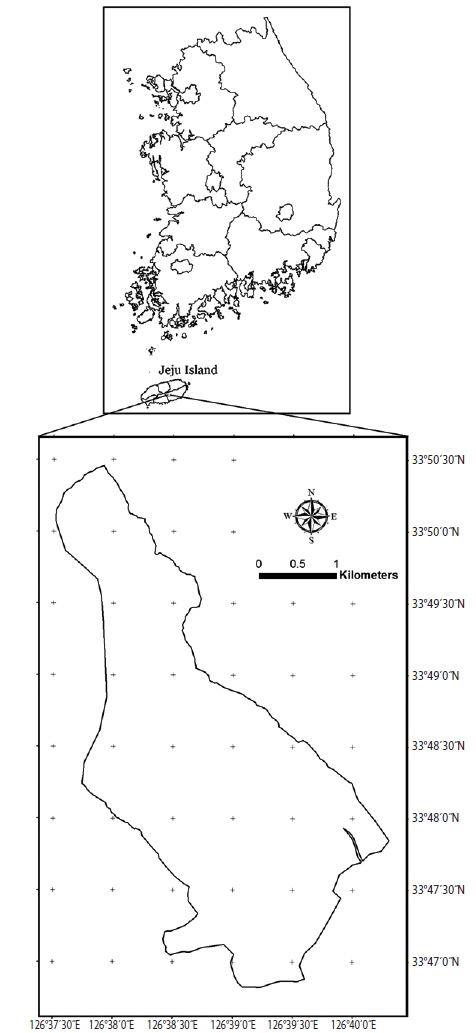
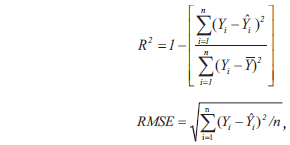Jeju Island is located in the southernmost part of Korea between 33°11′27″−33°33′50″ N and 126°08′43″−126°58′20″ E (Lee et al. 2009). The highest mountain in South Korea, Mount Halla, can be found in this island (highest peak: 1,950 m above sea level). This island has a total land area of 184,840 ha (1 ha = 104 m2) and approximately 88,874 ha (48%) of this island has forest cover (Korea Forest Service 2012). A forest (approximately 1,203 ha) is located in the Hannam experimental forest, southeast of Mount Halla. This forest has diverse and essential roles because it is considered as timber production forest, recreational forest, water conservation forest, environmental conservation forest, and seed orchard (Korea Forest Service 2014). However, there is still insufficient study on the biomass of this forest, specifically for the
Forest ecosystems are very important as they are considered as a major sink for carbon and have the potential to release this carbon in cases of deforestation and degradation (Lim et al. 2013). According to Dixon et al. (1994), approximately 80% of all aboveground carbon (C) and 40% of belowground C were stored in the forest biomass. Based on the United Nations Framework Convention on Climate Change (UNFCCC), different countries are mandated to accurately determine the biomass and carbon stocks available in their forests and regularly report their forest resources status (Levy et al. 2004, Basuki et al. 2009, Kim et al. 2011). Biomass estimations in forests have become a major research interest because of the significant role of forests in global climate change (Watson et al. 2000, Lehtonen et al. 2004, Tobin and Nieuwenhuis 2007, Bollandsas et al. 2009, Teobaldelli et al. 2009, Li et al. 2010). Therefore, accurately estimating the biomass of forests is a critical step in the estimation of carbon stored in the forest (Xiao and Ceulemans 2004, Fehrmann et al. 2008, Chung et al. 2009, Hosoda and Iehara 2010).
The 5th National Forest Inventory (NFI) in Korea was conducted from 2006 to 2010 by the Korea Forest Service to determine the state of the forests, but one of the disadvantages of the forest inventory data was the lack of direct biomass measurements (Lehtonen et al. 2004). Teobaldelli et al. (2009) suggested the allometric equations as one of the methods that can be applied to determine the biomass of the forests by using the data from the NFI. Various authors (Fang and Wang 2001, Tobin and Nieuwenhuis 2007, Lim et al. 2013) also suggested using stem densities and biomass expansion factors (BEF), which convert stem volume to total tree biomass including branches and leaves (Li et al. 2010). BEF can be applied in the NFI data to estimate the biomass in the forests. Such interpretations of forestry data and retrospective calculation of biomass are widely accepted as the Intergovernmental Panel on Climate Change (IPCC) guidelines (IPCC 2003) recommended stem density and BEF could be used to accurately estimate biomass and carbon stocks of a forest (Korea Forest Research Institute 2010). However, allometric equations for
In 1920, the
Plots with a size of 30 m by 30 m were established in the
The relationship of DBH to the different biomass components such as stem, stem wood, stem bark, needles, branches, and root and to the total biomass was analyzed. The model form used to develop the biomass equations was presented below:
where
This model form was used in this study as it was recommended by the Korea Forest Research Institute (2006b) for Korean tree species and was previously used to develop biomass equations for the different Korean tree species (Park and Moon 1994, Kim et al. 1995, Son and Kim 1998, Son et al. 2001, Seo et al. 2013). To correct the bias in log-transformed allometric equations, Sprugel correction factors (Sprugel 1983) were used (Son and Kim 1998, Son et al. 2001, Annighofer et al. 2012) and was calculated as:
where CF represents correction factor and SEE is standard error of estimate.
The collected data were statistically analyzed by using SAS 9.1 (SAS Institute Inc. 2004). The coefficient of determination (
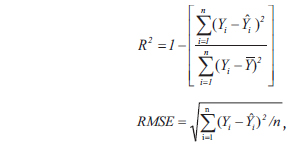
where
Stem density was calculated by dividing the dry weight of the stem (biomass) by its total volume (g cm-3) (IPCC 2006). BEF was determined by dividing the biomass of the upper part of a tree (such as branches and foliage) by the stem biomass (stem wood and bark) (Hosoda and Iehara 2010, Korea Forest Research Institute 2010).
>
Biomass estimation and allometric equation
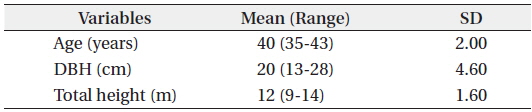
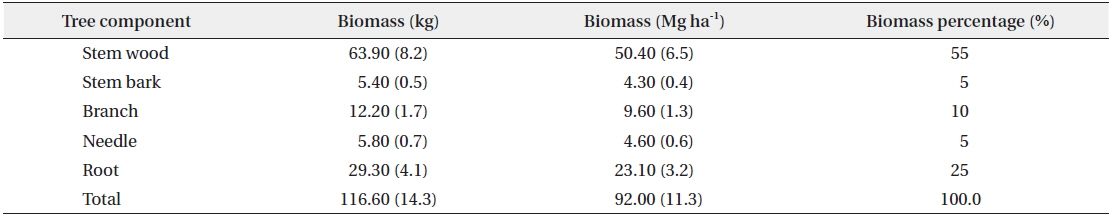
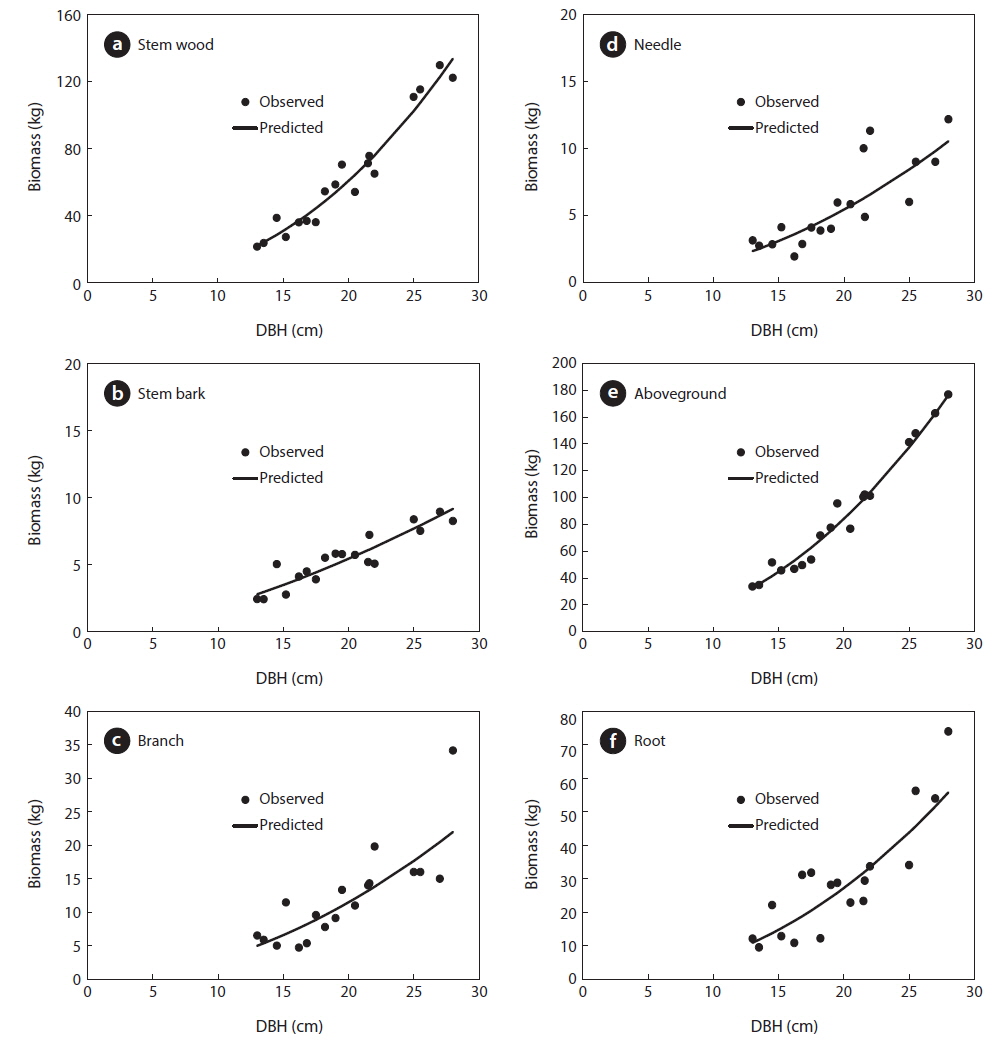
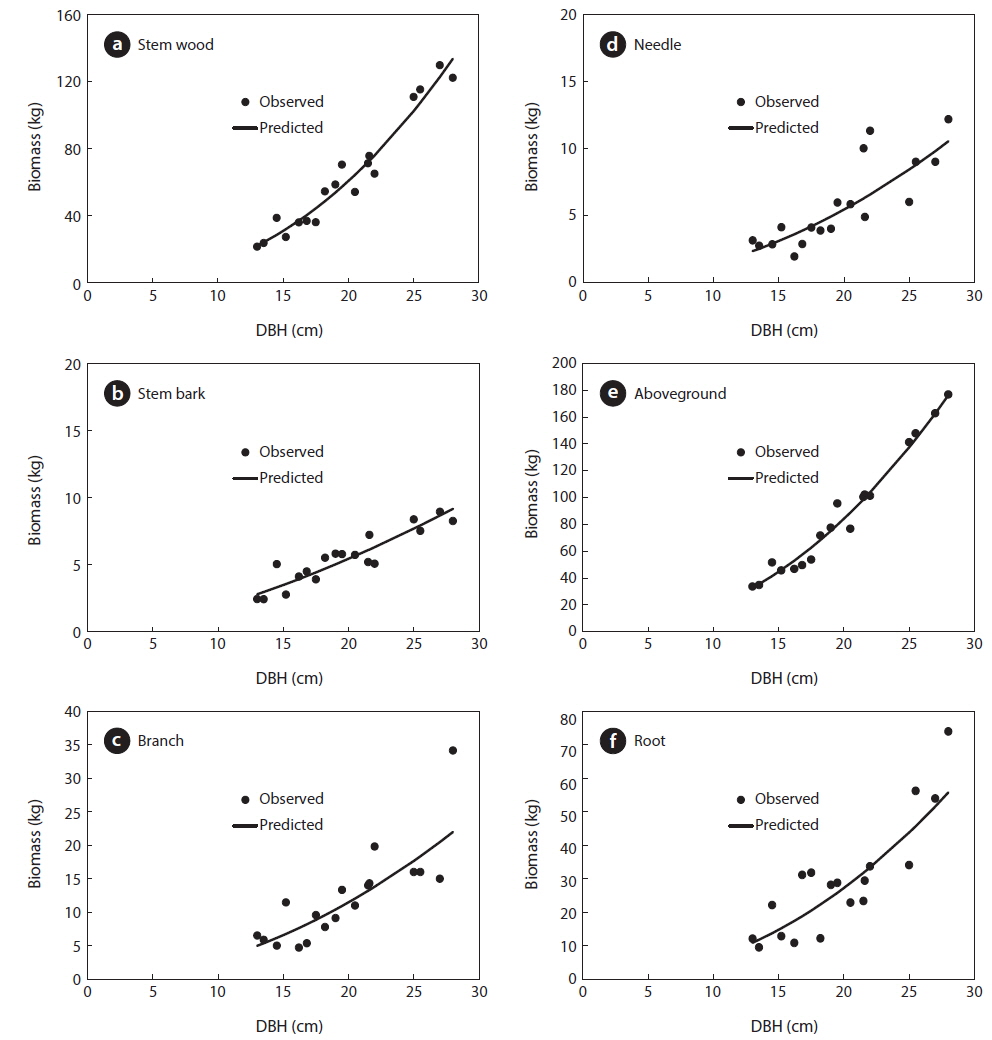
Results of the data collection showed that the mean DBH was 20 cm ranging from 13 to 28 cm. For the total height, the mean was 12 m ranging from 9 to 14 m and the mean age was 40 years ranging from 35 to 43 years (Table 1). Results also showed that the mean total biomass was 116.60 kg, with the stem wood having the highest mean biomass with 63.90 kg followed by root (29.30 kg), branch (12.30 kg), needle (5.80 kg), and stem bark (5.40 kg), respectively (Table 2). On the stand level, the biomass density of the
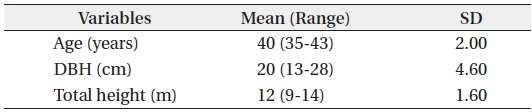
Summary of the observed statistics of the harvested Cryptomeria japonica trees in Hannam experimental forest, Jeju Island, Korea
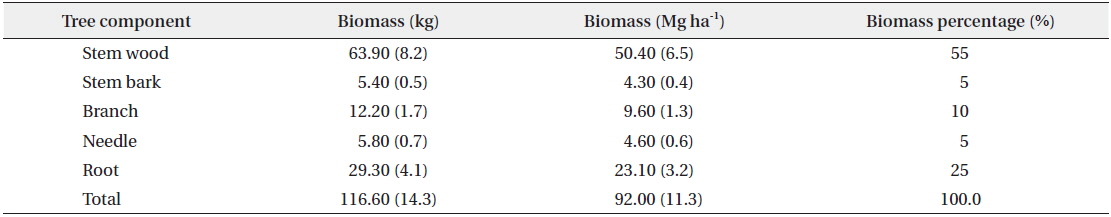
Tree component biomass and the biomass distribution ratio of Cryptomeria japonica in Hannam experimental forest, Jeju Island, Korea
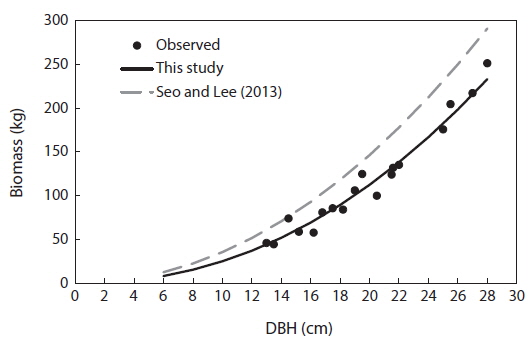
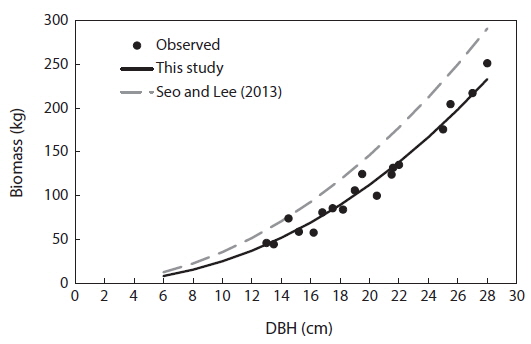
For the biomass distribution, the stem wood accounted for the highest biomass percentage of 55%; followed by the root, 25%; branch, 10%; needle, 5%; and stem bark, 5% (Table 2). This result showed that excluding secondary tree biomass components like needles, branches, and roots was not recommended as it would significantly underestimate the total biomass and carbon storage potential of a forest (Peichl and Arain 2007, Seo et al. 2013). In comparison to the study of Seo and Lee (2013), the biomass distribution of the younger
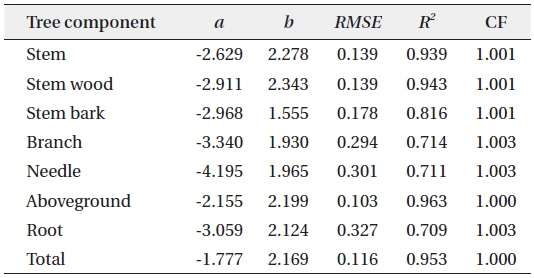
Allometric equations that estimated the different biomass components and total biomass of
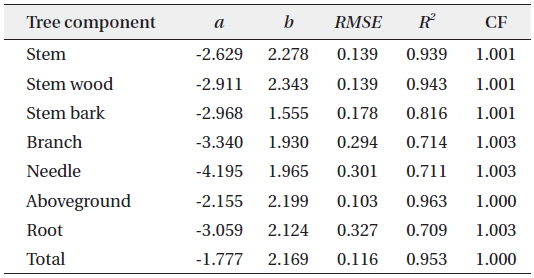
Allometric equations for the different tree biomass component of Cryptomeria japonica in Hannam experimental forest, Jeju Island, Korea
Total height was not used as a predictor variable in this study because in most forest inventories, such as the 5th NFI of Korea, only representative trees were measured for total height, due to the difficulty and high cost, while all the DBH of the trees in a plot were collected (Seo et al. 2013). Furthermore, Montagu et al. (2005) stated that DBH was considered as the best predictor variable in order to obtain a stable relationship. Various studies reported that DBH was sufficient to accurately estimate the tree biomass because of the difficulty to accurately measure the total height of the trees (Lim et al. 2013) and that adding total height as second variable did not significantly improve the biomass estimations of the models (Annighofer et al. 2012).
>
Stem density and biomass expansion factor
This study did not only estimate the biomass and develop allometric equations but it also determined the stem density, BEF, and the ratio of the root biomass to aboveground biomass (R) of the
The estimated aboveground BEF of the
The R of
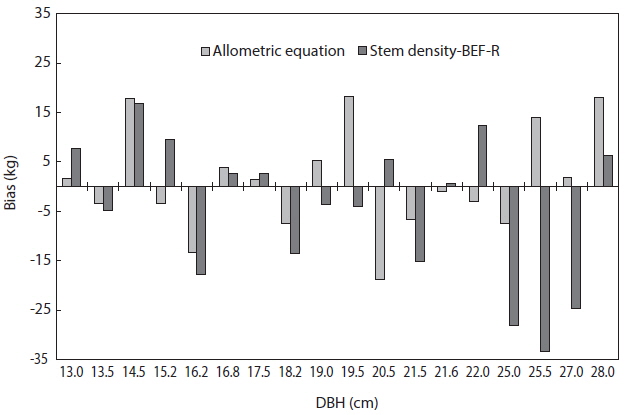
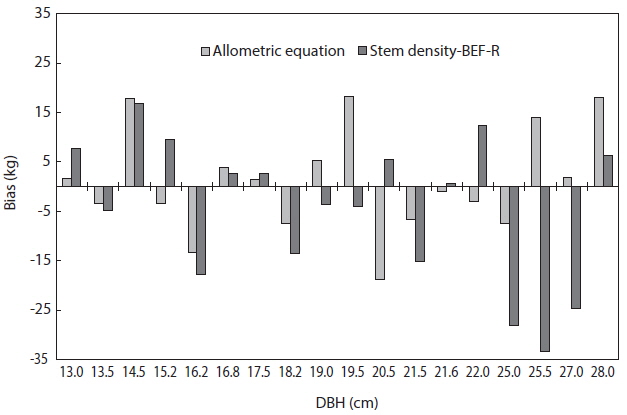
The total biomass that were predicted using both the allometric equation (method 1) and the stem density, BEF and R (method 2) were compared to the observed biomass of the 18 trees. The total biomass using method 2 was calculated as:
Total biomass = tree volume × stem density × BEF × R.
The biases of the two methods were determined as shown in Fig. 4. This comparison showed that the method 1 had a better predictive capability as compared to the method 2. A negative value in the bias indicates over prediction of total biomass while a positive value indicates under prediction of total biomass. The mean bias was also determined for the two methods. Method 1 had a mean bias of 1 kg while method 2 had a mean bias of -4.5 kg. Thus, the more suitable method to estimate the total biomass of
Allometric equation using DBH as predicting variable was developed, and the stem density, BEF, and R were determined to accurately estimate the biomass of