현재, 해양에서 어족생물의 현존 자원량 평가, 어초에 대한 자원조성효과의 평가, 물리환경 및 지층구조의 탐사 등과 같은 각종 해양계측분야에서 chirp (compressed high intensity radar pulse) 기술이 널리 활용되고 있다. 그 중에서도 특히, 어업생물자원의 계측과 관련하여 chirp 기술을 채택한 소너(sonar) 및 어군탐지기가 널리 보급되고 있는데, 그 이유 중의 하나는 해중이나 해저 부근에 군집하고 있는 어족생물 상호간의 거리분해능 개선, SNR (signal to nose ratio, 신호대 잡음비)의 향상, 또한, 낮은 출력으로 높은 신호이득을 얻을 수 있다는 장점 때문이다(Airmar, 2013). 종래의 어군탐지기에서는 거리분해능이 전적으로 송신신호의 펄스폭에 의해 결정되었지만, chirp 어군탐지기에서는 펄스폭과 관계없이 chirp 신호의 주파수 대역폭에 의해 결정되기 때문에 주파수 대역을 증대시킬수록 어족생물 상호간의 거리분해능을 향상시킬 수 있다. 일반적으로 해중으로 방사되는 음향신호의 출력은 첨두출력(peak power)과 송신펄스 폭의 곱으로서 정의되기 때문에 종래의 어군탐지기에서는 어류의 탐지능력을 향상시키기 위하여 첨두출력을 증대시키거나, 또는 펄스폭을 증대시키는 방법이 널리 이용되어왔다. 이 경우, 첨두출력을 증대시키면 장치의 규모가 커지고, 펄스폭을 증대시키면 거리분해능이 저하하는 상충문제(trade-off)가 발생한다. 그러나, chirp 어군탐지기에서는 거리분해능이 펄스폭과 독립적이기 때문에 펄스신호의 첨두출력을 낮추더라도 그에 비례하는 만큼, 펄스폭을 증대시키면 종래의 어군탐지기와 동일한 탐지성능을 얻을 수 있기 때문에 낮은 출력으로 높은 신호이득을 얻을 수 있는 장점이 있다. 이와 같이 긴 펄스폭을 갖는 음향신호가 해중의 어족생물로부터 산란될 때, 넓은 주파수 범위의 chirp 산란신호를 펄스압축처리, 즉 matched filter를 통과시켜 처리하면 각종 해중 잡음이 제거된 고분해능의 echogram을 얻을 수 있다(Chu and Stanton, 1998). 이에 부가하여 chirp 어군탐지기에서는 종래의 어군탐지기와 비교하여 매우 넓은 주파수 대역의 펄스신호를 해중으로 송신하기 때문에 대상어류로부터 더 다양한 주파수 의존적인 어종식별정보를 얻을 수 있다(Stanton and Chu, 2008; Stanton et al., 2010; Stanton et al., 2012).
따라서, 본 연구에서는 어족생물의 형상, 이석(otolith)을 포함하는 내부조직, 부레의 유무 등과 같은 생태학적 차이에 따른 음향산란신호의 변동특성을 정량적으로 분석, 고찰하기 위한 목적으로, 9개의 어종을 대상으로 95-220 kHz의 주파수 대역에서 측정한 chirp 산란신호의 진폭 및 주파수 스펙트럼(spread spectrum)의 자세각 의존성을 분석하고, 또한, 펄스압축기술을 통해 추출된 chirp echo 응답패턴을 바탕으로 어종식별과 관련된 어체 내부에 대한 산란요소(scattering feature)의 분포패턴 등을 분석, 고찰하였다.
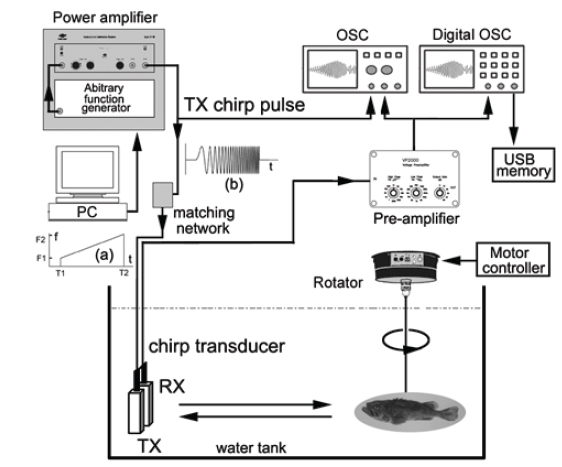
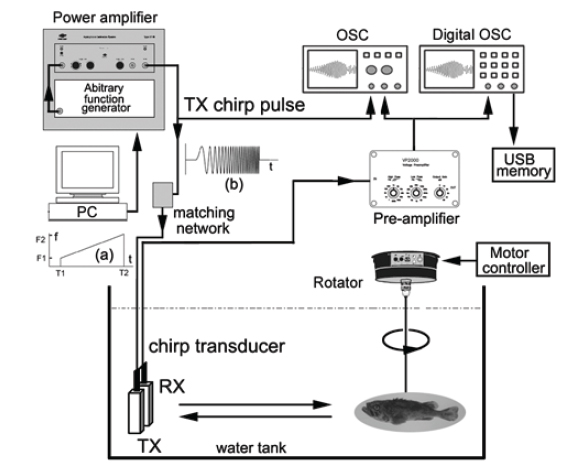
실험어류에 의한 광대역 음향산란신호를 계측하기 위한 chirp 어군탐지 시스템의 구성 모식도는 Fig. 1과 같다. Fig. 1에서 어류에 의한 chirp 산란신호의 계측은 먼저 PC에서 일정한 펄스폭(0.3 ms)과 펄스반복주기(0.02 s)를 갖는 선형주파수 변조(linear frequency modulation) 신호, 즉, chirp 신호를 발생시켜 이것을 RS232C 인터페이스를 통해 임의파형발생기(33120A, HP, USA)의 메모리에 전송하여 저장한 후, 필요에 따라 이 chirp 신호를 호출하여 사용하였다. 본 연구에서 이용한 up-chirp 펄스신호
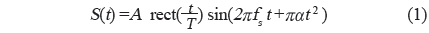
여기서,
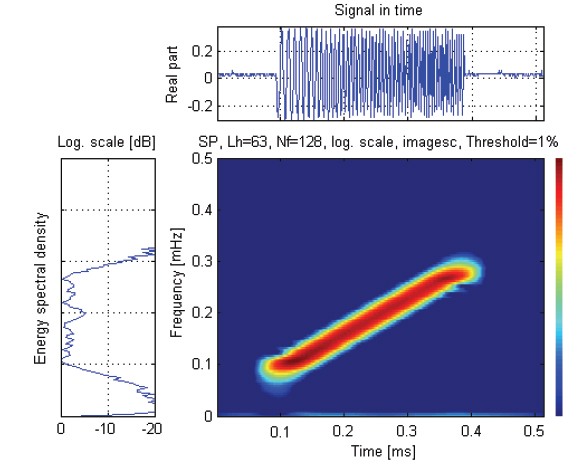
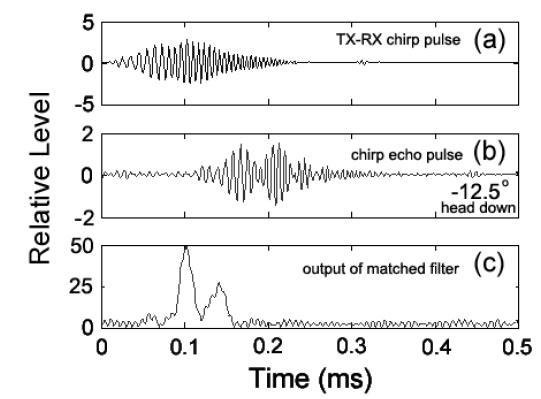
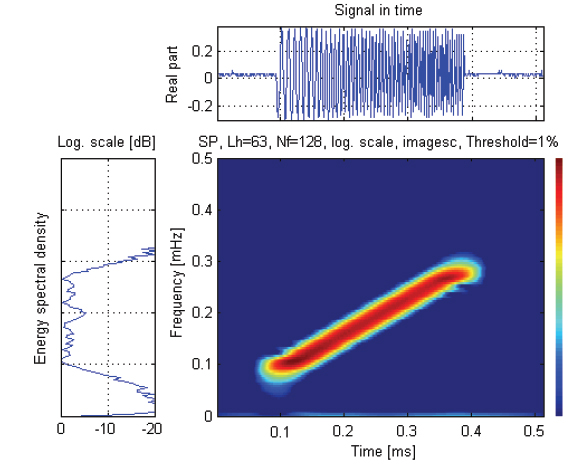
실험은 임의파형발생기의 메모리에 저장되어 있는 up-chirp 펄스를 호출하여 출력전압과 주파수 대역을 각각 400 mV, 90-280 kHz로 조정하여 전력증폭기(2713, B&K, Denmark)를 거쳐 40 dB로 증폭한 후, 이 신호를 정합회로(matching network)를 경유하여 광대역 초음파 변환기(R203, Airmar, USA)에 공급하였는데, 그 chirp 송신 펄스신호의 시간주파수 응답특성은 Fig. 2와 같다.
Fig. 2에서 어류에 의한 광대역 산란신호는 chirp 펄스의 송신에 사용한 것과 동일한 또 하나의 광대역 음향변환기를 사용하여 수신하였는데, 이들 송신과 수신용 음향변환기는 투명 아크릴 수조(L×B×D, 1.8×1.2×1.2 m)의 한쪽 벽면에 나란히 설치하였다. chirp 산란신호의 계측은 0.02%의 MS-222 (sigma chemical, Canada)로서 마취시킨 활어 개체어를 송∙수신 광대역 음향변환기의 음축상 약 1.2 m 거리에 나이론 monofilament (Ø=0.3 mm)를 사용하여 현수시키고, 어체의 등쪽이 진동면과 수평이 되도록 깊이와 자세를 정밀하게 조정하면서 수행하였다. 즉, 회전장치(turn table)를 제어하여 ±45° pitch 범위(+: head up, -: head down)에 대한 몸체(fish body)의 경사상태와 신호변동의 연속성 등을 확인하면서 +45°, 0°, −45°의 순서로 pitch 각 변화에 따른 산란신호의 변동을 약 1° 간격으로 측정하였다. 이 때, 수신 음향변환기를 통해 측정된 chirp 산란 신호는 대역필터와 전치증폭기(VP2000, RESON, Denmark)를 통해 필터링 및 증폭한 후, 디지털 오실로스코프(DS1530, EZ, Korea) 및 스펙트럼 분석기(LSA-30, LIG Nex1, Korea)의 USB 인터페이스를 통해 chirp 산란신호의 시간응답파형과 주파수 스펙트럼 등을 수록하였다.
본 연구에서 사용한 실험어류는 부레를 갖는 부세(
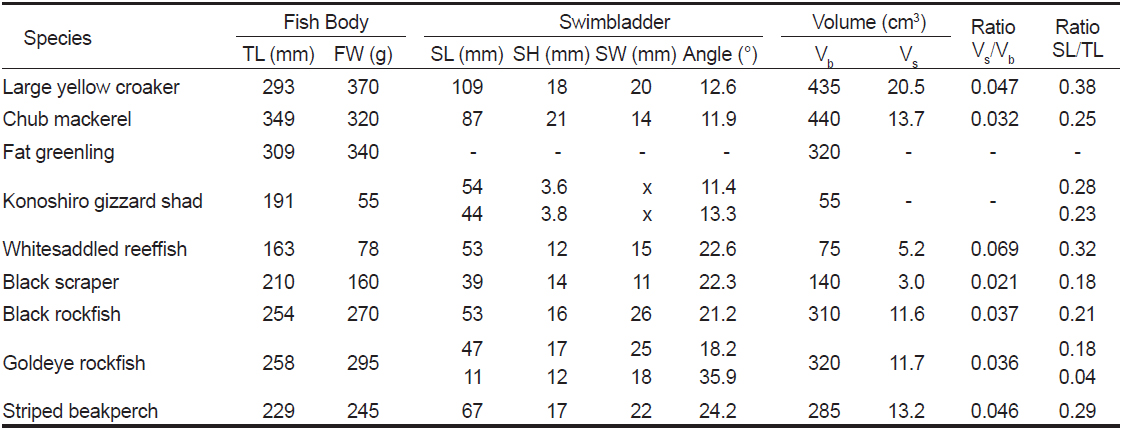
[Table 1.] Morphometric measurements of swimbladders for 9 fish species obtained from X-ray images
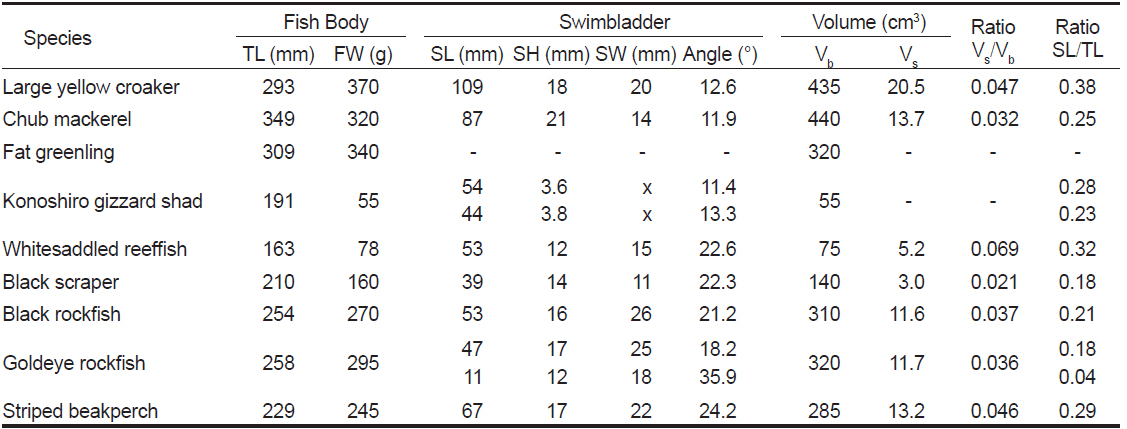
Morphometric measurements of swimbladders for 9 fish species obtained from X-ray images
한편, 불볼락의 부레의 형상은 2개의 가스 체임버를 갖는 이중 체임버 (double chamber)형이므로, 그 부레체적은 전부(anterior chamber)와 후부(posterior chamber)의 각각에 대하여 a, b 및 c 값을 구하여 산출하였다.
>
chirp 산란신호의 펄스압축 및 압축파형의 평활화
펄스압축은 chirp 소너 시스템이나 chirp 레이더 시스템 등에서 탐지물표의 거리 분해능이나 SNR을 증대시키기 위해 이용하는 신호처리기술이다. 본 연구에서 이용한 up-chirp 신호
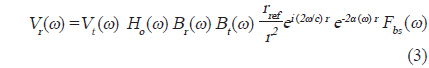
여기서,
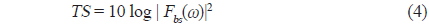
에 의해 구할 수 있다(Reeder et al., 2004). (3)식에서 어군탐지기의 송∙수신 모드에 대한 시스템 응답특성과 개체어 위치에 대한 수중에서의 왕복(two-way) 전반손실을 각각
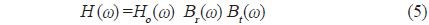
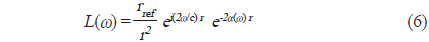
이 되므로,
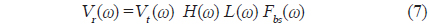
의해 구할 수 있다. 어류가 존재하지 않는 상태에서 송파기와 수파기를 서로 마주보도록
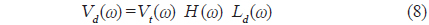
이 된다. 따라서, 개체어에 대한 산란신호
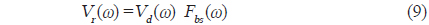
와 같이 나타낼 수 있다. 이 때, 시간영역에 대한 matched filtering 신호처리는 개체어의 산란신호
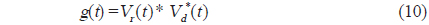
에 의해 구할 수 있고(Chu and Stanton, 1998), 여기서, *은 컨벌루션을 의미한다. 일반적으로 어떤 신호
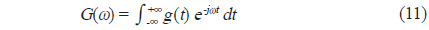
에 의해 정의되므로, 주파수 영역에서 matched filter를 통과한 어류에 의한 산란신호
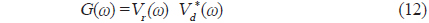
에 의해 구할 수 있다(Maurice, 2007). 이 식으로부터 matched filter 출력, 즉, 압축펄스신호 y는
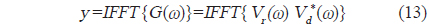
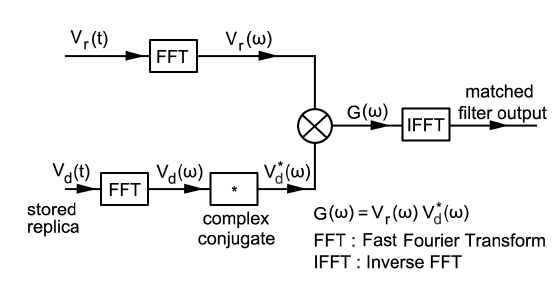
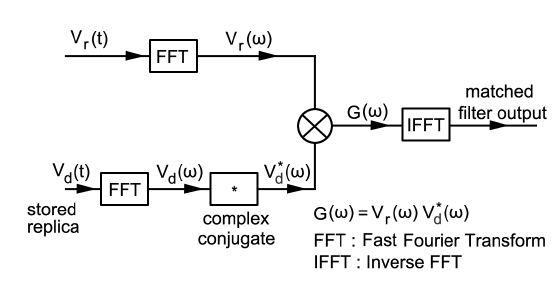
에 의해 구할 수 있는데(Wang and Yao, 2009), 이들 matched filtering 신호처리에 대한 모식도는 Fig. 3과 같다(Ahmed et al., 2014).
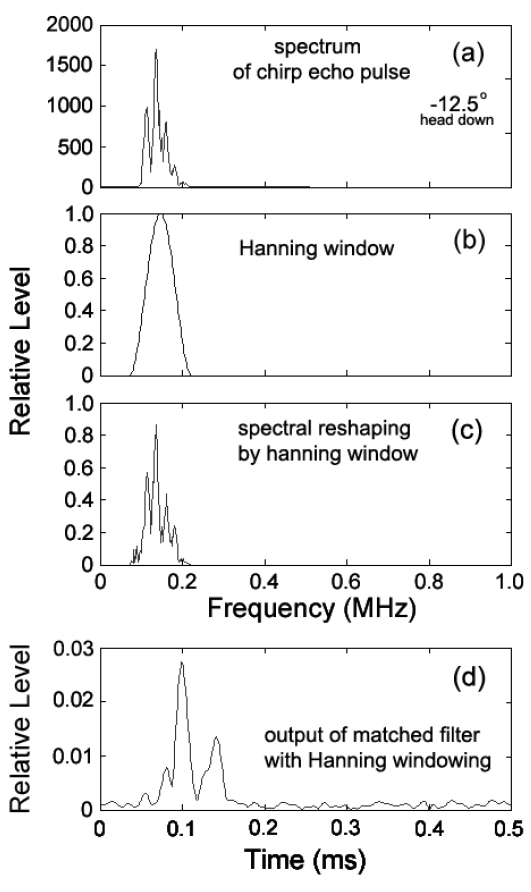
한편, 이와 같은 matched filter의 출력, 즉 압축된 echo 펄스에는 많은 측엽(side lobe)들이 함께 나타나기 때문에 이들 측엽레벨을 억제시키고, 또한, 어군탐지기 자체에 기인하는 응답성분을 제거하기 위하여 역 필터링(inverse filtering) 처리와 윈도우(window) 함수
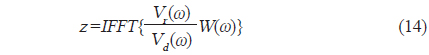
에 의해 구할 수 있는데(Lord, 2000), 본 연구에서는 윈도우 함수로서 Hanning 윈도우를 이용하였다.
>
실험어류의 X-ray 영상 및 부레의 형상학적 특징
본 연구에서 실험에 사용한 9개 어종에 대한 X-ray 영상은 Fig. 4와 같다. Fig. 4에서 a~i의 X-ray 영상은 각각 부세(a), 고등어(b), 쥐노래미(c), 전어(d), 자리돔(e), 말쥐치(f), 조피볼락(g), 불볼락(h), 돌돔(i)에 대한 영상이다. 본 연구에서는 Fig. 4의 X-ray 영상을 토대로 어류에 의한 산란신호의 자세각 및 주파수 의존성을 조사, 분석하였다. 이 때, 어류의 자세 변화가 산란신호의 변동에 미치는 영향을 각 어종별로 비교, 고찰하기 위하여 X-ray 영상부터 각 어종에 대한 부레의 경사각, 형상, 크기, 체적 등에 대한 파라미터를 산출하였는데, 그 결과는 Table 1과 같다. Table 1에서 부레의 길이와 높이는 각 개체어의 측면 X-ray 영상으로부터, 또한, 부레의 폭은 복부 방향에 대한 X-ray 영상으로부터 구하였다. 여기서, 부레의 각 부 치수를 구할 때, 등쪽 방향에 대한 X-ray 영상을 사용하지 않은 이유는 척추골 등의 내부 어체 조직에 대한 영상과 부레의 영상이 서로 중첩되어 부레만의 선명한 영상을 얻는데 어려움이 있었기 때문이다.
Fig. 4 및 Table 1에서 알 수 있는 바와 같이 쥐노래미의 X-ray 영상(Fig. 4c)에서는 부레를 관찰할 수 없지만, 나머지 8개 어종(부세, 고등어, 전어, 자리돔, 말쥐치, 조피볼락, 불볼락, 돌돔)의 X-ray 영상에는 다양한 형태와 크기의 부레가 선명하게 나타나 있다.
Fig. 4에서 불볼락과 전어는 두 개의 독립적인 부레(double chambered swimbladder)를 갖고 있는 반면, 나머지 어류는 단지 하나의 잘 발달된 부레(single chambered swimbladder)를 갖고 있다. 불볼락은 체장 방향으로 앞쪽에는 큰 체임버(chamber)가, 뒷쪽에는 작은 체임버가 서로 붙어 있지만, 전어는 체고 방향으로 상부에는 길고 가느다란 체임버가, 하부에는 상부에 비해 그 길이가 짧은 체임버가 서로 상하로 분리되어 존재한다. 부세는 유선형의 긴 부레(길이 109 mm)가 척추골 하부에 12.6°의 경사각을 유지하면서 분포하는데, 몸체에 대한 부레의 체적비는 4.7%, 길이비는 38%이었다. 북해 연안에서 어획되는 고등어에는 부레가 존재하지 않지만, 우리나라 주변해역에서 어획되는 고등어에는 Fig. 4에서와 같이 반달모양의 부레(길이 87 mm)가 11.9° 경사되어 복부 부근까지 팽창되어 있는데, 몸체에 대한 부레의 체적비와 길이비는 각각 3.2%, 25%이었다. 전어의 경우에는 부레의 복부 X-ray 사진으로부터 정확한 부레의 형상을 추출하기 어려워 체적비를 산출하지 않았고, 다만, 길이비는 상부의 경우, 28%, 하부는 23%이었다. 말쥐치는 다른 어류에 비해 두개골 조직이 매우 견고하면서 상부의 단면적이 상대적으로 큰 형태학적 특성을 갖는다. 또한, 부레는 척추골 하부에 매우 잘 발달된 형태로 존재하면서 두개골 조직과는 상당한 거리를 두고 분포하고 있다. 이와 같이 음향 임피던스가 서로 다른 두 이질적인 산란요소가 일정한 거리를 두고 서로 분리되어 있으면, 이들 요소로부터 음파가 산란될 때, 두 요소에 의한 산란신호의 위상이 서로 보강 및 상쇄간섭을 일으켜 매우 규칙적인 음향산란패턴을 나타낸다(Jaffe, 2006; Jaffe and Roberts, 2011). 말쥐치의 몸체에 대한 부레의 체적비는 2.1%, 길이비는 18%이었다. 한편, 조피볼락과 불볼락은 몸체에 대한 부레의 체적비가 각각 3.7%와 3.6%로서 거의 같았고, 공통적으로 매우 견고한 두개골 조직을 갖고 있지만, 조피볼락의 경우에는 부레가 두개골 조직과 서로 인접하여 분포하고 있는 반면, 불볼락은 부레가 두개골 조직과 상당한 거리를 두고 분포하고 있음을 알 수 있었다. 이 때문에 말쥐치에서와 같이 이들 두 요소에 의한 산란신호는 서로 보강 및 상쇄간섭을 일으켜 음향산란패턴을 주기적으로 변동시킬 것으로 판단된다. 한편, 돌돔의 부레는 하나의 체임버를 갖고 있지만, 전부의 부레 공간은 13.6°, 후부의 공간은 39.4° 기울어져 분포하고(전체의 경사각 24.2°), 그 체적비는 4.6%이었다.
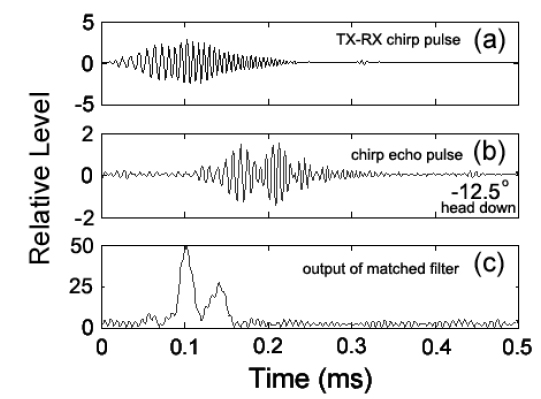
불볼락에 대한 chirp 산란신호를 대상으로 펄스압축처리, 즉 matched filter를 통과시킨 결과는 Fig. 5와 같다. Fig. 5에서 (a)는 실험수조에 어류가 존재하지 않는 상태에서 측정한 chirp 펄스의 직접파 신호로서, 본 연구에서는 이 신호를 기준신호(replica)로 이용하여 각 어종의 개체어로부터 얻어진 chirp 산란신호의 펄스압축처리를 수행하였다. (b)는 불볼락의 head-down −12.5° 자세각에서 측정한 chirp 산란신호의 파형이고, (c)는 matched filter를 통과시킨 펄스압축된 검파파형이다. Fig. 5(c)에서 알 수 있는 바와 같이 펄스압축파형은 Fig. 5(b)의 산란파형과 비교하여 공간거리 분해능과 SNR이 매우 향상되었지만, matched filter의 출력신호에는 많은 측엽들이 나타나고 있는데, 이것은 chirp 어군탐지기의 echogram에 허상을 유발시키는 요인이 되므로 제거할 필요가 있다. 따라서, 본 연구에서는 matched filter의 출력, 즉 압축된 echo 응답신호에 나타나는 측엽 레벨을 억제시킬 목적으로 역 필터링 처리를 통해 스펙트럼 패턴을 성형하였는데, 그 결과는 Fig. 6과 같다.
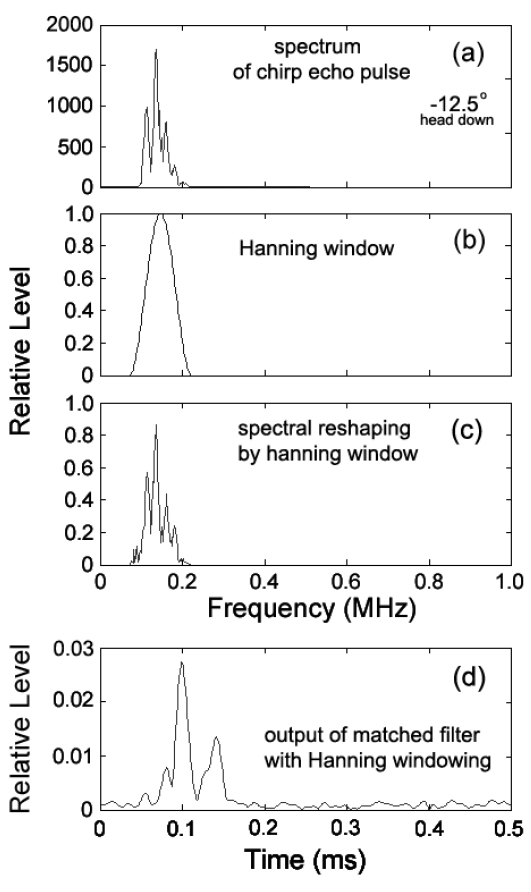
Fig. 6에서 (a)는 불볼락의 head-down −12.5° 자세각에서 측정한 chirp 산란 신호의 주파수 스펙트럼이고, (b)는 본 연구에서 가중함수로서 이용한 Hanning 윈도우이다. (c)는 (a)의 chirp 산란신호의 주파수 스펙트럼에 (b)의 가중함수를 적용하여 성형한 chirp 산란신호의 새로운 주파수 스펙트럼이다. 한편, (d)는 (c)의 Hanning 윈도우에 의해 성형된 echo 스펙트럼을 다시 Fourier 역변환하여 얻은 새로운 펄스압축파형의 검파파형이다. Fig. 5(c)에 대한 기존의 펄스압축파형과 Fig. 6(d)의 본 연구에서 추출한 펄스압축파형을 서로 비교해 볼 때, 측엽 레벨이 대폭 억제되어 인접하는 산란신호 상호간의 분해 및 식별이 매우 용이하게 되었음을 알 수 있다. chirp 어군탐지기에서 거리분해능은 chirp 펄스신호의 지속시간(폭)에 의해 결정되기 때문에 본 연구에서 사용하고 있는 up-chirp 펄스의 대역폭(
>
펄스압축기법에 의한 광대역 음향산란패턴의 식별 및 분석
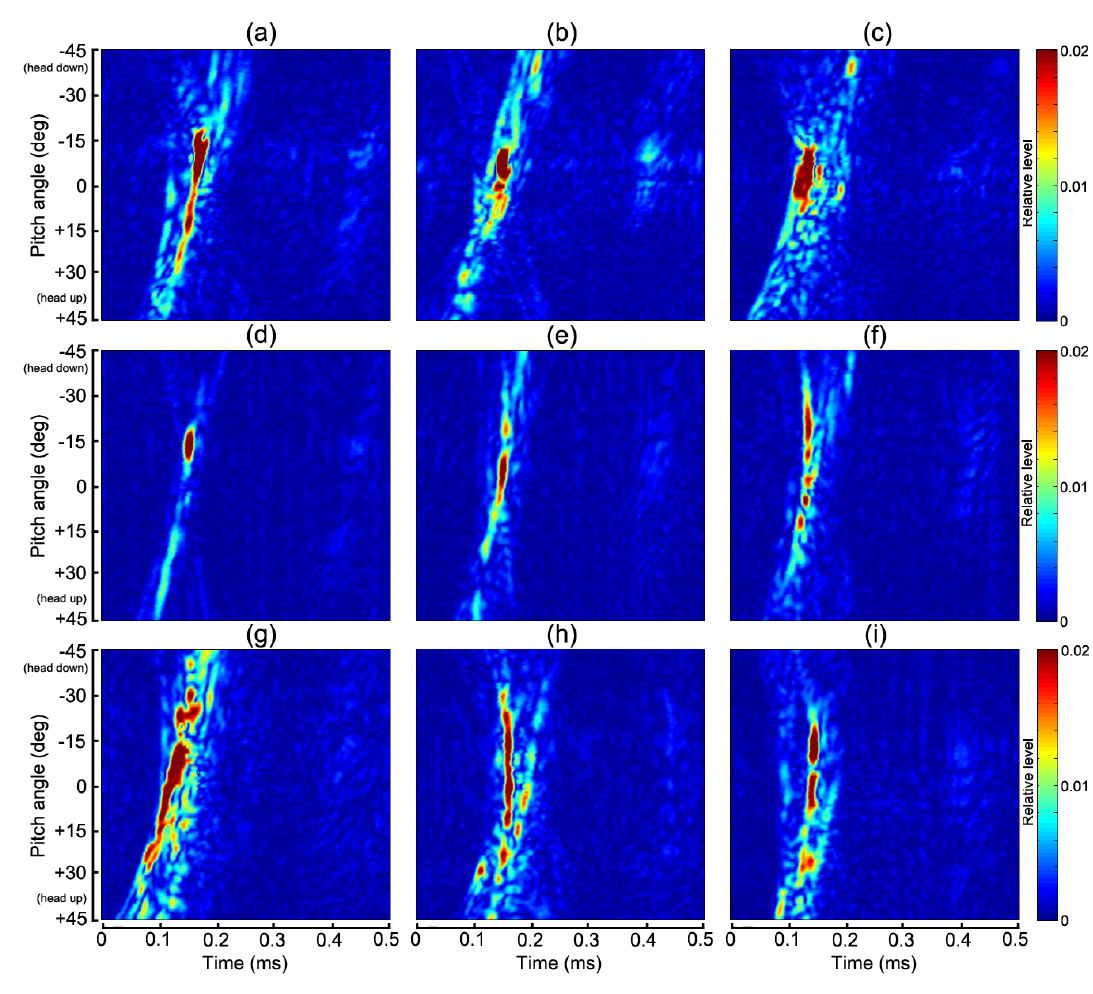
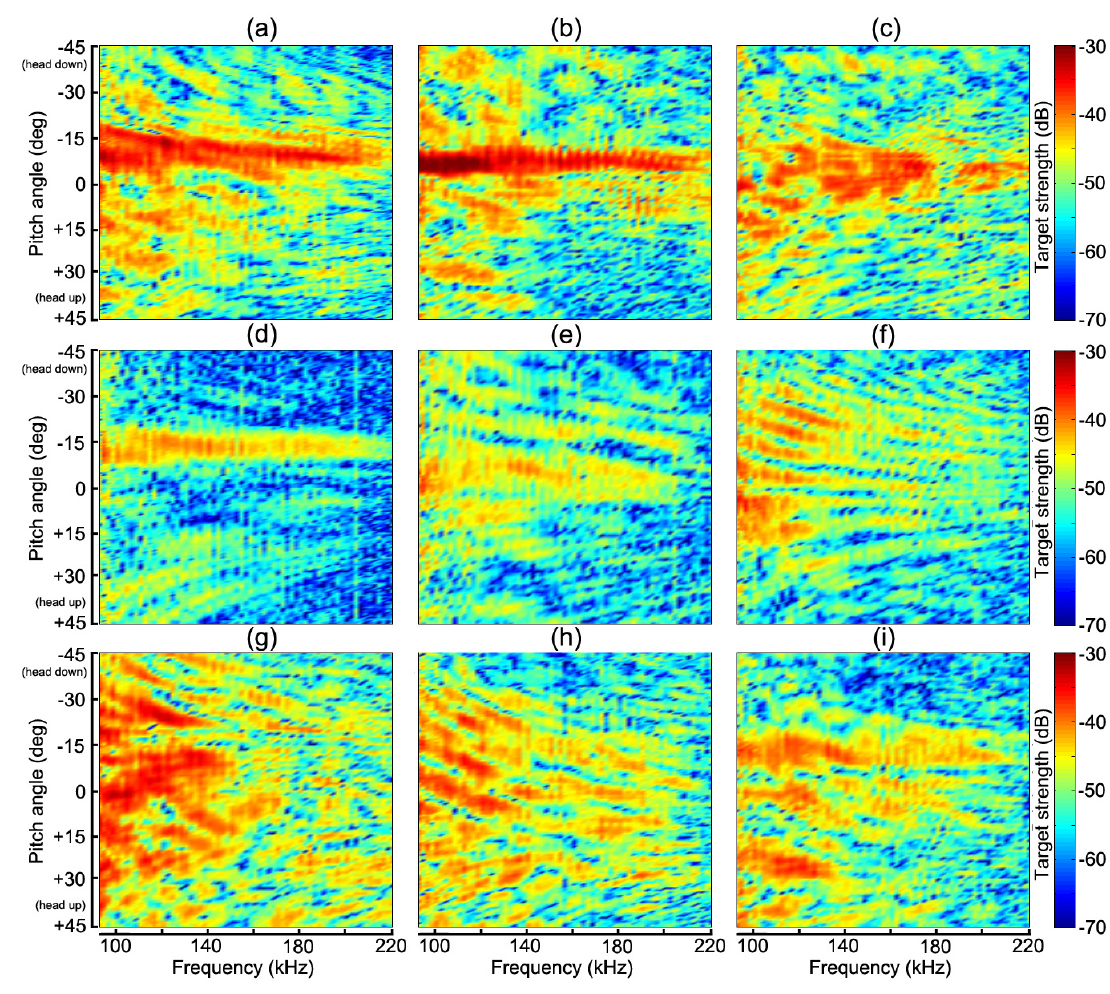
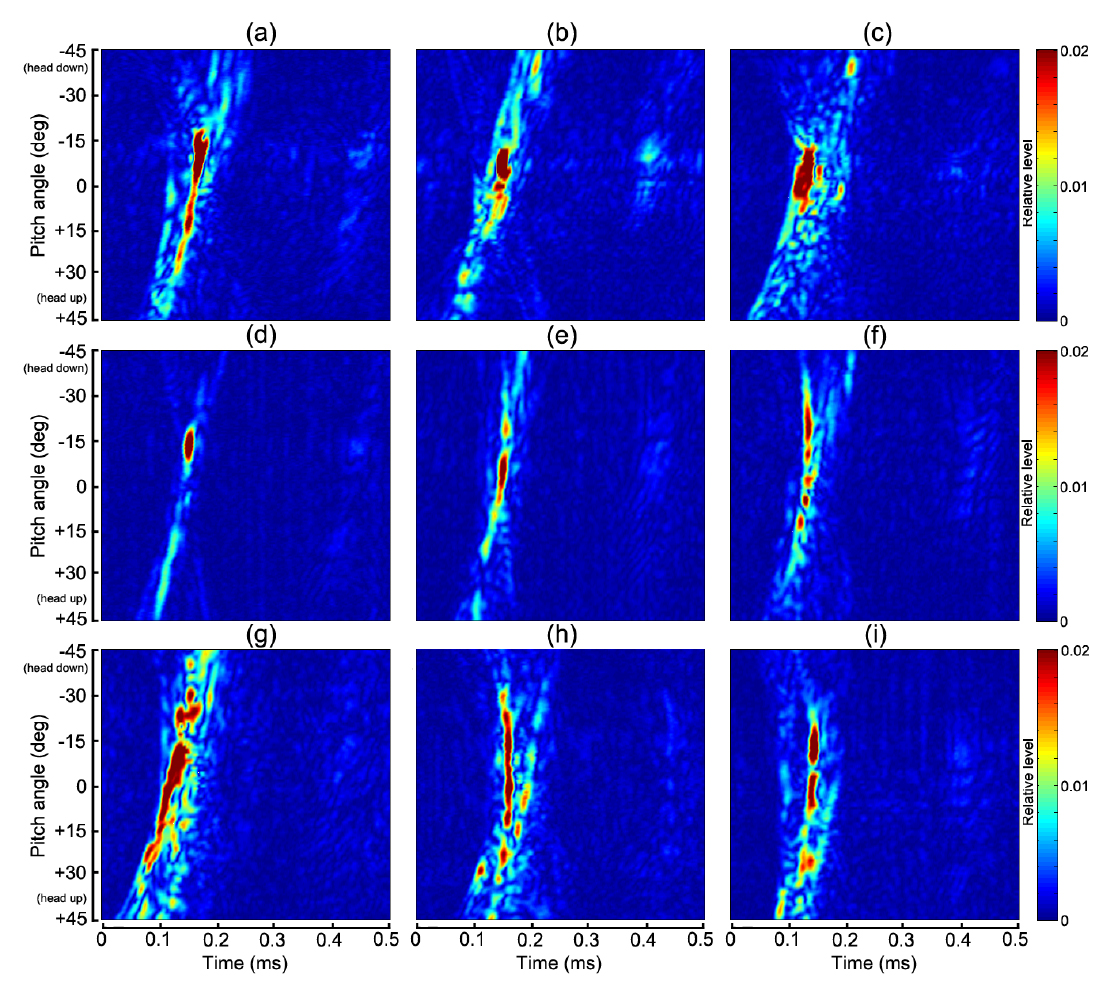
Fig. 4 및 Table 1에서 알 수 있는 바와 같이 쥐노래미를 제외한 모든 어종은 부레를 갖고 있지만, 부레의 형상, 기울기, 몸체에 대한 체적비, 길이비 및 부레 위치 등은 각각 서로 다른 패턴을 나타내고 있다. 생물학적으로 이들 부레의 가장 중요한 기능 중의 하나는 부력조절이지만, 음향학적으로는 주변 어체 조직과의 음향 임피던스의 차이가 매우 크기 때문에 음파를 강하게 산란시키는 산란체로서의 기능을 발휘한다(Lovik and Hovem, 1979; Foote, 1980; Fassler et al., 2009). 본 연구에서는 이 점에 주목하여 Fig. 6에 나타낸 펄스압축기법을 각 개체어의 산란 신호에 적용하여 어종식별정보를 추출하였는데, 각 어종에 대한 산란신호의 진폭과 주파수 스펙트럼에 대한 자세각 의존성, 또한, 반사강도와 압축펄스응답에 대한 자세각 및 주파수 의존성을 각각 분석한 결과는 각각 Fig. 7, Fig. 8, Fig. 9 및 Fig. 10과 같다.
Fig. 7에서 echo 응답특성은 0~1.2 V의 컬러패턴으로, Fig. 8 의 주파수 스펙트럼은 −15~+30 dB의 상대진폭으로, Fig. 9의 반사강도는 −70~ −30 dB의 컬러 패턴으로, 또한, 압축펄스 응답레벨은 0~0.02 범위에 대한 무차원 상대진폭의 컬러패턴으로 각각 나타내었다. Fig. 4에 나타낸 각 어종별 부레의 X-ray 영상은 마취시킨 실험어류를 동물병원으로 이송하여 공기중(대기압 상태)에서 촬영한 것이므로 수심의 변화에 따른 부레체적의 변화는 전혀 고려되어 있지 않다. Fig. 4과 Table 1에 나타낸 각 어종별 개체어의 부레는 그 형상과 체적이 서로 다르기 때문에 음파에 대한 산란응답특성 및 공진특성도 서로 다르고, 또한, 체적이 감소하면 공진 주파수도 증가할 것으로 판단된다. 특히, 군집을 이루고 있는 경우에는 각 개체어의 부레들이 서로 연성공진(coupling resonance)을 나타내어 단일 부레에 의한 공진 주파수 보다 더 낮은 쪽으로 공진 주파수가 이동하는 경향이 있다(Nero et al., 2007). 이 때문에 최근에는 부레의 공진특성을 조사하여 어종을 식별하려는 연구가 시도되고 있지만(Stanton et al., 2010; Stanton et al., 2012), 본 연구의 Fig. 7~Fig. 10에 나타낸 결과들은 이들 문제에 대한 고려 없이, 단지 대기압 상태에서 측정한 chirp 산란신호의 진폭, 주파수 스펙트럼, 반사강도 및 압축펄스신호의 자세각 및 주파수 의존성에 대해서만 분석, 고찰하였다.
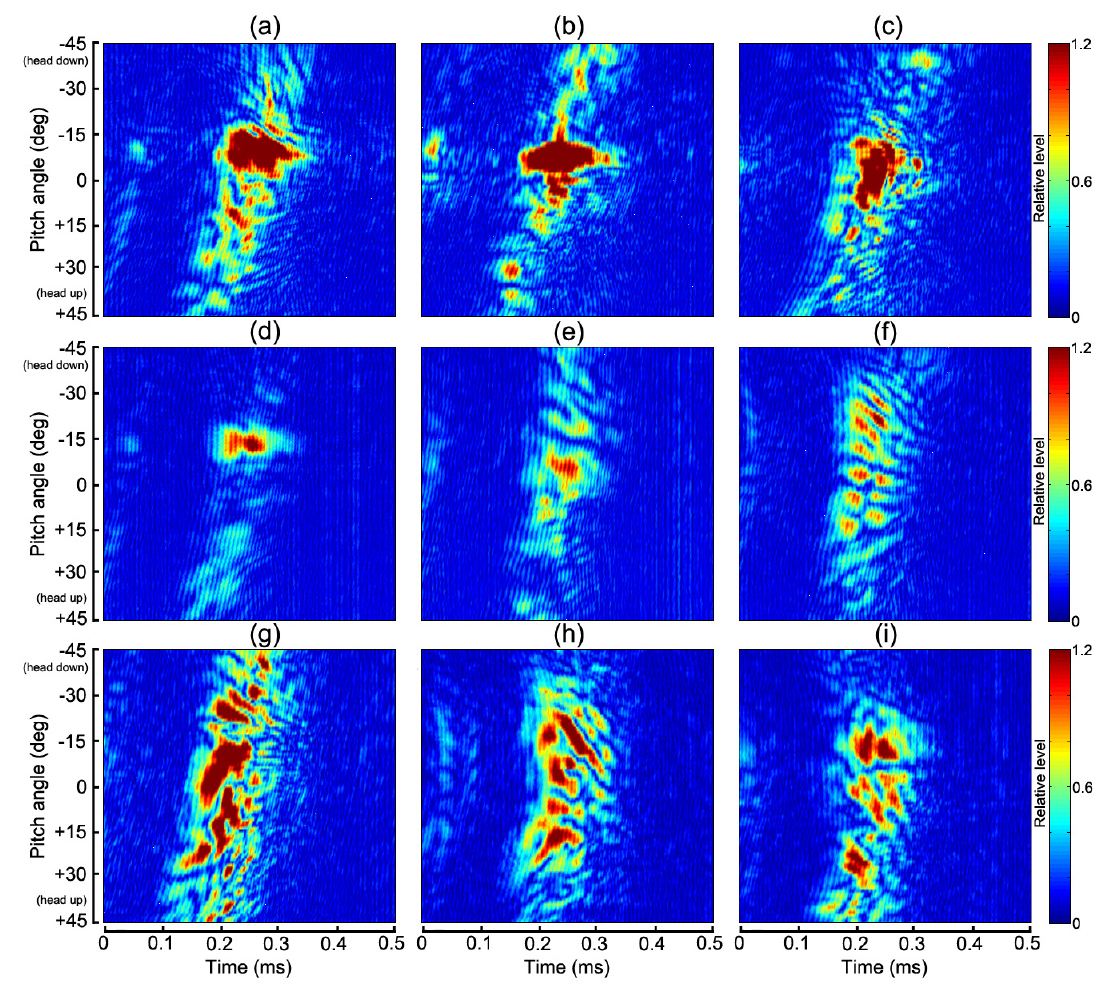
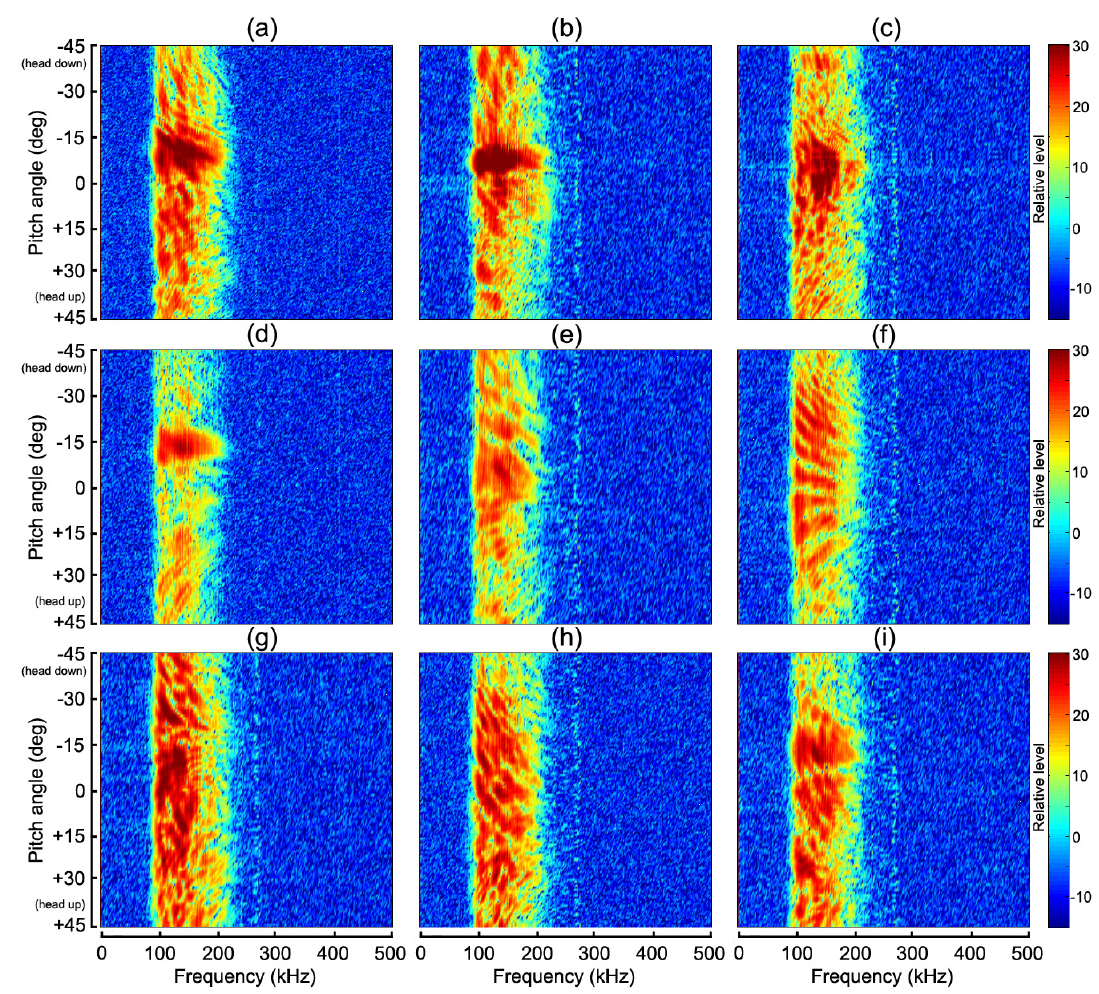
Fig. 7~Fig. 10에서 (a)~(i)의 2차원 echogram은 각각 부세(a), 고등어(b), 쥐노래미(c), 전어(d), 자리돔(e), 말쥐치(f), 조피볼락(g), 불볼락(h), 돌돔(i)에 대한 결과이다. 본 연구에서는 Fig. 9과 Fig. 10에 나타낸 반사강도와 압축펄스응답의 자세각 및 주파수 의존성을 바탕으로, 그 응답특성의 유사성이 인정되는 어종 그룹을 4개 그룹, 즉, 부레가 없는 쥐노래미, 부레를 갖는 어종으로서 부세와 고등어 그룹, 전어, 자리돔 및 말쥐치 그룹, 조피볼락, 불볼락 및 돌돔 그룹 등으로 구분하여 분석 및 고찰하였다.
Fig. 7~Fig. 8에서 부레가 존재하지 않는 쥐노래미의 경우, 산란진폭의 시간응답은 자세각 0°를 중심으로 −15°~+10°의 위치각과 0.2~0.3 ms의 시간구간 범위에 집중되어 나타났다. 또한, 주파수 스펙트럼 역시 시간응답특성과 거의 동일한 위치각 범위의 120-180 kHz의 주파수 구간에서 강한 응답특성을 나타내었다. 이로부터 쥐노래미의 산란신호에 대한 진폭 및 주파수 응답특성은 수직입사방향, 즉, 등쪽 부근의 좁은 입사각 범위에 제한되어 나타나고 있음을 알 수 있었다. 이와 같은 응답특성은 쥐노래미의 두개골, 척추 뼈, 육질조직 및 이석 등과 같은 어체 내부조직 상호간의 밀도 차(density contrast) 및 음속 차(sound speed contrast)가 매우 미소하기 때문에 산란성분의 중첩이 수직입사 방향에 집중되어 나타나는 것으로 판단된다. 또한, Fig. 9(c)와 Fig. 10(c)에서 쥐노래미의 반사강도와 압축펄스신호는 산란진폭의 시간 및 주파수 응답특성과 거의 유사하게 자세각 0°를 중심으로 −15°~+10° 범위의 120-180 kHz의 주파수 영역에서 강한 응답특성을 나타내었고, 부레가 존재하지 않는 관계로 이들 어체 내부의 산란요소에 의한 echo 생성의 기여도는 분해하여 식별할 수 없었다.
Fig. 4와 Table 1에서 부세와 고등어의 부레는 척추골 아래쪽으로 휘어져 길이 방향으로 길게 뻗어 있는데, 몸체에 대한 부레의 체적비율은 각각 4.7%와 3.2%, 길이비율은 각각 38%와 25%, 부레의 자세각은 각각 12.6°와 11.9°이었다. 어체 내부 조직에 의한 echo 형성에 가장 중요한 요소가 되는 부레 형상은 부세의 경우는 유선형, 고등어는 반달형이었다. 특히, Fig. 4의 X-ray 영상에서 부세의 이석은 부레와 일직선상의 두개골 내부에 존재하고 있지만, 고등어에서는 분명하게 식별되지 않았다. 이들 두 어류의 부레는 음향 임피던스가 서로 다른 이석기관을 포함하는 두개골 조직과 함께 산란신호를 생성하는 주요 음향 산란요소이다. 이 때문에 Fig. 7과 Fig. 8에서 알 수 있는 바와 같이 부세의 경우, 산란진폭과 주파수 스펙트럼의 자세각 의존성은 약 −12.6°의 자세각을 중심으로 −7°~ −15°의 위치각 범위의 95-220 kHz 주파수 구간에서 강한 응답특성을 나타내었다. 고등어의 경우에는 부레의 경사각에 상당하는 약 −11.9°를 중심으로 −5°~ −13°의 위치각 범위에서, 또한, 주파수 스펙트럼은 시간 응답특성과 거의 같은 위치각 범위의 95-220 kHz 주파수 구간에서 강한 응답특성을 나타냈다. Fig. 7~Fig. 9에서 산란신호의 진폭, 주파수 스펙트럼 및 반사강도에 대한 자세각 의존성은 고등어의 경우, 부세에 비해 더 좁은 자세각 범위와 더 넓은 주파수 범위에 걸쳐 집중되어 나타나고 있음을 알 수 있었다. 이것은 Fig. 4에서 알 수 있는 바와 같이 고등어의 부레가 반달 모양으로서 음파가 부레 경사면에 수직 입사할 때, 부레의 위쪽과 아래쪽 사이의 공간간격이 부세에 비해 약 17% 정도 더 넓어 이들 두 임피던스의 불연속 경계면에서 생성되는 산란신호의 간섭이 더 긴 시간 동안 지속되었기 때문이다. 반면, 부세에 대한 산란신호의 자세각 의존성은 고등어와 반대로 더 넓은 자세각 범위와 더 좁은 주파수 범위에 강하게 집중되어 있음을 알 수 있었다. 이것은 부레의 위쪽과 아래쪽의 두 불연속 경계면에 대한 공간간격이 고등어 보다 좁아 산란신호의 간섭지속시간이 짧았고, 또한, 부레의 길이가 고등어 보다 길어 자세각의 변화에 대한 응답이 더 넓은 범위에 걸쳐 나타났기 때문이다. 한편, Fig. 10의 펄스압축에 대한 부세의 응답신호는 head-up 약 45°부터 미약하게 나타나기 시작한 후, head-up 약 30° 부근에서 응답레벨이 강해져 거의 직선적으로 연속되다가 부레에 음파가 수직입사하는 head-down 약 −12.6° 부근에서 가장 강한 응답특성은 나타내었다. 이 때, head-up +30° 부근에서부터 부레의 응답신호 전방에 또 다른 산란성분이 출현하여 자세각 0° 근방까지 거의 직선적으로 나타나는데, 이것은 이석기관을 포함하는 두개골 조직에 의한 산란응답신호로 판단된다(Nesse et al., 2009). 고등어의 압축펄스응답은 head-up 약 +15°의 자세각부터 강한 응답특성이 나타나기 시작하여 보강 및 상쇄간섭을 거친 후, 부레에 음파가 수직입사하는 head-down 약 −11.9° 부근에서 가장 강한 응답특성은 나타내었다. 고등어에 있어서도 두개골 조직에 의한 산란성분은 부세에서와 같이 head-up +30° 부근에서부터 미약하게 출현하지만, 부레에 의한 응답신호 바로 전방에 매우 근접하여 나타나고 있는 것으로부터 고등어에 대한 두 산란요소의 중심간격은 부세의 경우보다 더 작은 것으로 판단된다. 특히, 이들 어종에 있어서는 음파가 부레 경사면에 수직으로 입사하는 약 ±4° 자세각의 좁은 구간에서 echo 응답이 매우 강하고, 또한 집중적으로 나타나지만, 그 자세각으로부터 head-up, 또는 head-down 방향으로 자세가 점차 기울어지면 어체 근육에 기인하는 감쇠(damping) 성분이 echo 생성을 억제시켜 echo 응답이 급속히 약화되는 패턴을 나타내었다.
Fig. 4의 X-ray 영상으로부터 전어의 부레는 척추골과 완전히 분리되어 2개의 층상 구조를 하고 있는데, 상부의 부레 공간은 길이와 폭이 각각 54 mm, 3.6 mm이고, 경사각은 11.4°, 또한, 하부의 부레공간은 길이와 폭이 각각 44 mm, 3.8 mm이고, 경사각이 13.3°이었다. Fig. 7과 Fig. 8에서 전어에 대한 산란신호의 진폭 및 주파수 스펙트럼 응답은 head-up 약 45° 부근부터 미약하게 출현하기 시작하여 head-down 약 −11°~ −16°의 좁은 구간에서 강한 응답특성을 나타내었다. 또한, Fig. 9의 반사강도의 자세각 및 주파수 의존성 패턴에 있어서는 Fig. 7과 Fig. 8에서와 거의 유사하게 자세각 −11°~ −16° 범위의 95-220 kHz의 주파수 영역에서 강한 응답특성을 나타내었다. 한편, 자세각 20°~45°의 좁은 구간에서는 미약하지만 저주파 영역으로부터 고주파 영역을 향해 반사강도가 방사선 모양으로 휘어지는 패턴을 나타내었으나, 이와 관련된 echo 응답은 Fig. 10의 펄스압축응답패턴에서는 관찰할 수 없었다. 또한, 전어에 있어서도 부세와 고등에서와 같이 음파가 부레 경사면에 수직으로 입사하는 약 ±2.5° 자세각의 매우 좁은 구간에서 echo 응답이 매우 강하고, 또한 집중적으로 나타났고, 그 자세각 범위로부터 벗어나면 어체 근육에 기인하는 감쇠성분이 echo 생성을 강하게 억제시켜 echo 응답이 급속히 약화되는 패턴을 나타내었다.
자리돔의 부레는 척추골을 따라 앞쪽에서는 10.5°, 뒤쪽에서는 22.6° 기울어져 마치 가지 모양으로 휘어져 존재하고, 그 연장선상의 두부 쪽에는 안구 바로 뒤쪽에 작은 이석이 분포하고 있다. 이들 이석을 포함하는 두개골 조직과 앞쪽 부레 부분은 매우 인접하여 분포하고 있는 관계로 하나의 산란요소로 작용하고, 뒤쪽에 큰 각도로 기울어져 존재하는 부레 영역은 또 다른 음향산란요소로 작용한다. Fig. 7 및 Fig. 8에서 산란신호의 진폭 응답은 head-up 약 20° 부근부터 점차 강하게 출현하기 시작하여 head-down 약 −5°~ −12°의 좁은 구간에서 강한 응답을 나타내었고, 주파수 스펙트럼 응답은 자세각 +5°~ −15°의 구간의 95-200 kHz 주파수 영역에서 강한 응답특성을 나타내었다. 또한, Fig. 10의 펄스압축파형에서 앞쪽의 10.5°기울어져 있는 부레 영역과 이석을 포함하는 두개골 조직에 의한 산란성분이 head-up 약 20° 부근부터 보강과 상쇄간섭을 반복하면서 출현하기 시작하였고, 바로 후방에 매우 미약하지만 뒤쪽의 부레 영역에 의한 echo 응답이 나타났다. 이들 두 산란요소에 의한 응답신호는 자세각 약 5° 부근에서 서로 중첩되어 하나의 응답으로 결합되었고, 앞쪽 부레 영역에 수직 입사하는 head-down 약 −10.5° 부근에서 가장 강한 응답을 나타내었다. 그 후, 자세각 −15° 부근에서 상쇄간섭이 나타나 산란성분이 일시적으로 소멸되었다가 뒤쪽의 부레 영역에 수직 입사하는 자세각 −22.6° 부근에서 다시 강한 echo 응답특성을 나타내었다. head-down 약 −20° 이후부터는 다시 두개골 조직과 앞쪽 부레의 산란요소에 기인하는 산란성분이 매우 미약하게 출현하였다. 이 때문에 Fig. 9에서 알 수 있는 바와 같이 앞쪽 부레 공간에 수직으로 입사하는 자세각 −10.5°를 기준으로 좌∙우측으로 고주파 영역으로부터 저주파 영역으로 이동해 감에 따라 반사강도 패턴이 방사선 모양으로 휘어져 분포하는 패턴을 나타내었다. 이것은 부레의 앞쪽 공간과 이석을 포함하는 두개골 조직이 독립된 하나의 산란 요소로, 또한 후부의 부레공간이 또 다른 독립된 산란요소로 작용하여 두 요소에 의한 산란성분이 자세각 −10.5°를 중심으로 서로 상쇄 및 보강간섭을 일으킨 결과라고 판단된다.
한편, 말쥐치의 부레는 활 모양으로 휘어진 척추골을 따라 −22.3° 경사져 존재하고, 부레의 체적중심은 두개골 조직의 체적중심, 특히 주둥이 앞단과 거의 일직선상에 존재한다. 또한, 안구와 등쪽의 가시(spine) 등을 포함하는 두개골 조직과 부레는 그 산란중심간격이 다른 어류에 비해 상대적으로 더 커서 산란신호의 간섭현상이 강하게 발생하였는데, 특히, 말쥐치의 두개골 조직은 다른 어류보다 뼈 조직이 견고하고 커서 음향산란체로서 중요한 역할을 하였다. Fig. 10의 압축펄스파형에서 알 수 있는 바와 같이 head-up 13.6° 부근의 자세각부터 두개골 조직에 의한 echo 응답이 매우 미약하게 나타났고, 그 뒤쪽에 부레에 의한 echo 응답이 자세각 0° 부근까지 보강과 상쇄 간섭이 반복되면서 나타나고 있다. 또한, 자세각 0° 부근에서는 이들 산란요소에 의한 echo 응답이 일시적으로 중첩되었다가 완만하게 휘어져 head-down −30° 부근까지 강한 echo 응답이 지속되었다. 반면, 두개골 조직에 의한 echo 응답은 0°부터 head-down으로 자세가 변화되면서부터 부레에 의한 echo 응답으로부터 점차 벗어나는 변동패턴을 나타내었다. 즉, 두개골 조직과 부레에 의한 echo 응답특성이 자세각 0°를 기준으로 상하로 교차하면서 서로 다른 궤적패턴을 나타내었는데, 이것은 서로 인접하고 있는 두 개의 음향산란요소에 의한 echo 변동 패턴과 매우 유사하다. 따라서, 말쥐치의 경우에는 어체 내부에 두 개의 우세한 음향산란요소, 즉, 두개골 조직과 부레가 존재하지만, 두개골 조직의 산란강도는 부레에 비해 매우 미약한 패턴을 나타내었다. 이와 같은 현상은 Fig. 7의 산란진폭의 자세각 의존성에서 관찰할 수 있는 바와 같이 자세각 0°를 중심으로 echo 응답이 head-down과 head-up 방향으로 거의 좌우 대칭적인 방사선 패턴으로 휘어져 나타나는 특성과 밀접한 관계가 있다. 또한, Fig. 8의 주파수 스펙트럼의 자세각 의존성 역시 자세각 0°를 기준으로 고주파수 영역으로부터 저주파수 영역으로 이동함에 따라 echo 스펙트럼 응답이 거의 좌우 대칭적인 패턴으로, 또한, 거의 방사선 모양으로 휘어져 나타났다. 이들 두 패턴의 공통적 특성은 말쥐치의 부레와 두부 조직에 의한 산란신호 성분이 서로 보강 및 상쇄 간섭을 일으켜 시간과 주파수 영역에서 규칙적으로 변동하고 있는 점이다. Fig. 9의 반사강도의 자세각 및 주파수 의존성 패턴에서도 이들 부레와 두부 조직에 의한 산란성분이 서로 보강 및 상쇄간섭을 일으킨 결과, echo 응답이 고주파 영역에서 저주파 영역으로 이동함에 따라 자세각 0°를 기준으로 거의 대칭적으로 휘어지는 변동특성을 나타내었는데, 가장 강한 응답은 95-130 kHz의 저주파 범위에서 나타났다. 또한, Fig. 9에서 반사강도의 자세각 및 주파수 의존성은 고주파 영역에서는 echo 응답의 peak 및 null의 간격이 좁고, 주파수가 저주파 영역으로 이동함에 따라 그 간격이 점차 넓어지는 경향을 나타내었다. 이와 같은 현상은 음향 임피던스가 서로 다른 두개골 조직과 부레 사이에 발생하는 산란신호의 간섭이 저주파수 보다는 고주파수 영역에서 매우 복잡하게 일어나고 있음을 의미한다. 또한, Fig. 10에서와 같이 반사강도의 peak와 null의 위치를 정량적으로 추적하면 역으로 부레의 크기를 추정할 수 있다(Jaffe, 2006). 따라서, 어류의 공진산란을 측정하기 위해 10 kHz 이하의 저주파수 영역에서 산란신호를 수록 및 분석하는 경우에는 어체 내부조직 사이의 음향학적 간섭이나 어류의 형상에 의한 산란신호의 변동보다는 부레 자체의 공진에 의한 음향산란이 중요한 문제가 될 것으로 판단된다(Stanton et al., 2010; Stanton et al., 2012).
Fig. 4의 X-ray 영상으로부터 조피볼락의 부레는 척추골을 따라 −21.2° 경사져 완만한 “ㄱ” 자 모양으로 휘어져 존재하고, 그 연장선상의 두부 쪽에는 안구 바로 뒤쪽에 이석이 분포하고 있다. 이들 이석을 포함하는 두개골 조직과 부레는 매우 인접하여 분포하고 있는 관계로 Fig. 7의 산란진폭의 자세각 의존성에서 알 수 있는 바와 같이 이들 두 요소에 의한 산란신호는 자세각의 변화에 따라 매우 복잡한 보강 및 상쇄 간섭을 반복하면서 불규칙하게 변동하였다. 또한, Fig. 8의 산란신호에 대한 주파수 스펙트럼의 자세각 의존성에 있어서도 Fig. 7에서와 유사한 변동 패턴을 나타내었고, 95-150 kHz의 저주파 영역에서 강한 응답특성을 나타내었다. 한편, Fig. 10의 산란신호의 펄스압축파형에 대한 자세각 의존성에 있어서는 이석을 포함하는 두개골 조직의 음향산란중심과 부레의 음향산란중심이 서로 인접하고 있는 관계로 head-up 약 30° 부근에서부터 이들 두 요소에 의한 산란신호가 약 13° 부근까지 중첩되어 나타났다. 그 후, 15° 부근에서는 상쇄간섭에 의해 산란신호 성분이 일시적으로 소멸하였다가 음파가 부레에 수직으로 입사하는 −21.2° 부근에서는 매우 강한 echo 응답을 나타내었다. 또한, 조피볼락에 있어서는 다른 어류에서와 달리 부레에 의한 echo 응답이 발생한 이후에 불규칙한 응답신호가 전 자세각 범위에 걸쳐 나타났는데, 이것은 실험 당시 마취된 어류의 미소한 움직임에 기인하는 것으로 판단된다. 한편, Fig. 9의 반사강도의 자세각 및 주파수 의존성에 있어서는 head-down −15° 부근을 기준으로 −45° 까지의 범위에서는 고주파 영역에서 저주파 영역으로 이동함에 따라 반사강도가 방사선 모양으로 휘어져 변동하는 패턴을 나타내고 있으나, −15°에서 45°의 자세각 범위에서는 매우 불규칙한 변동 특성을 나타내었다.
한편, 불볼락의 부레는 X-ray 영상으로부터 볼 때, 두 개의 부레 영역이 서로 분리되어 있는데, 앞쪽 영역은 척추골 하부를 따라 완만하게 휘어져 큰 가스 체임버를 갖고 있는 반면, 뒤쪽 영역은 앞쪽 영역의 바로 뒤쪽에 붙어있으면서 작은 가스 체임버를 갖는다. 앞쪽 부레 영역에 대한 가스 체임버의 음향중심은 두개골 조직과 거의 일직선상에 위치한다. 그러나, 실제, head-up상태로부터 head-down의 자세로 이동해 갈 때, head-up 약 25°부터는 앞쪽 부레영역의 위쪽 경계면으로부터 음향산란이 먼저 발생하고, 그 다음에 두개골 조직에 의한 음향산란이 잇따라 발생하는 산란특성을 나타내었다. 이 때문에 Fig. 10에서 알 수 있는 바와 같이 head-up 약 30° 부근에서 두부에 의한 echo 응답이 먼저 나타나고, 약 25° 부근에서 부레와 두부조직의 산란성분이 서로 보강간섭을 일으켜 강한 응답특성을 나타내었다. 그 후, 그 자세각이 head-down으로 점차 이동해 감에 따라 부레에 의한 강한 echo 응답이 먼저 나타나고, 뒤이어서 두개골 조직에 의한 echo 응답이 일정한 간격을 두고 나타났다. head-up 15°부터는 매우 강한 부레의 echo 응답이 완만하게 휘어져 head-down 약 −30°까지 연속적으로 나타났다. 한편, 두부 조직에 의한 echo 응답패턴 역시 head-down 약 −30°까지 완만하게 휘어져 있는데, 이와 같은 현상은 말쥐치에서와 같이 두개골 조직과 부레의 산란중심 사이의 간격이 크기 때문인 것으로 판단된다. 또한, Fig. 8의 주파수 스펙트럼에 대한 자세각 의존성에 있어서는 부레와 두개골 조직의 echo 응답이 서로 분리되는 −30°~20° 자세각 구간에서 고주파 영역으로부터 저주파 영역으로 이동함에 따라 스펙트럼의 응답패턴이 head-down 방향으로 휘어져 나타나는 경향을 보였다. 그러나, Fig. 8의 echo 진폭에 대한 자세각 의존성 패턴에 있어서는 보강 및 상쇄 간섭이 불규칙하게 나타나 이와 같은 현상을 관찰할 수 없었다. 반면, Fig. 9의 반사강도의 자세각 및 주파수 의존성 패턴에 있어서는 부레와 두부조직이 서로 보강간섭을 일으켜 강한 echo 응답을 나타내었는데, 그 반사 강도 패턴은 head-up 약 25°의 자세각을 기준으로 고주파 영역으로부터 주파수가 점차 낮아짐에 따라 head-down과 head-up 방향으로 방사선 형태로 휘어져 나타났다.
한편, 돌돔의 부레는 머리에서 꼬리 쪽을 향하여 활 모양으로 휘어져 척추골 아래쪽에 위치하고 있는데, 부레의 앞쪽 영역의 경사각은 13.6°, 뒤쪽의 영역은 39.4°로 경사져 마치 표주박 형상을 나타내었다. Fig. 10의 압축펄스파형에서 어체가 head-up 자세로부터 점차 head-down 자세로 이동해 감에 따라 부레에 의한 echo 응답이 먼저 나타나고, 그 다음 두개골 조직에 의한 echo 응답이 뒤이어서 나타났다. 즉, Fig. 10에서 head-up 약 30°에서부터 시작되는 활 모양의 부레에 의한 강한 산란성분은 −25°까지 연속적으로 나타났고, 바로 뒤쪽에 두개골 조직에 의한 미약한 성분이 약 0° 부근까지 직선적으로 나타났다. Fig. 4의 X-ray 영상에서 알 수 있는 바와 같이 실험 당시 돌돔은 마취된 상태에서도 등지느러미가 강하게 돌출된 상태로 있었기 때문에 이것에 의한 산란성분이 부레에 의한 echo 응답의 전방에 미약하게 출현하였다. 이와 같이 돌돔의 경우에는 등지느러미, 부레, 두개골 조직 등에 의한 echo 응답이 서로 분리되어 나타나고 있는 것으로부터 이들 요소는 서로 독립적으로 echo 생성에 기여하고 있는 것으로 판단된다. Fig. 9의 반사강도의 자세각 및 주파수 의존성 패턴에서 가장 강한 응답은 앞쪽 부레 영역에 수직 입사하는 head-down 약 13.6° 부근의 전 주파수 영역에서 나타났다.

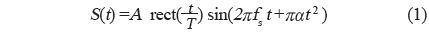
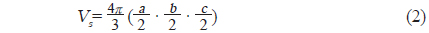
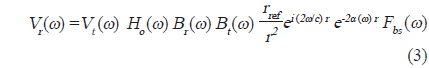
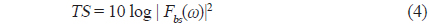
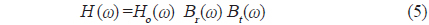
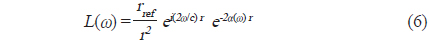
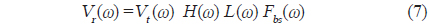
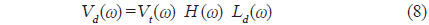
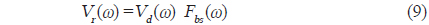
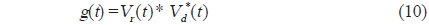

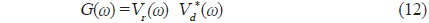
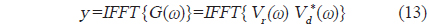
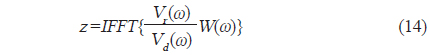
![Morphotypes observed on X-ray photographs of nine fish species. Photographs highlight the swimbladder and mineralized structures such as skeleton, rocker bone and otoliths. (a) large yellow croaker Larimichthys crocea, (b) chub mackerel Scomber japonicus, (c) fat greenling Hexagrammos otakii, (d) konoshiro gizzard shad Konosirus punctatus, (e) whitesaddled reeffish Chromis notata notata, (f) black scraper Thamnaconus modesutus [K], (g) black rockfish Sebastes schlegeli, (h) goldeye rockfish Sebastes thompsoni, (i) striped beakperch Oplegnathus fasciatus.](http://oak.go.kr/repository/journal/16273/KSSHBC_2015_v48n2_207_f004.jpg)
![Echograms of uncompressed broadband echo signals as a function of pitch angle for nine fish species. Variations in echograms are due to the shape and inclination of swimbladders and mineralized structures such as skeleton, bone and otoliths. (a) large yellow croaker Larimichthys crocea, (b) chub mackerel Scomber japonicus, (c) fat greenling Hexagrammos otakii, (d) konoshiro gizzard shad Konosirus punctatus, (e) whitesaddled reeffish Chromis notata notata, (f) black scraper Thamnaconus modesutus [K], (g) black rockfish Sebastes schlegeli, (h) goldeye rockfish Sebastes thompsoni, (i) striped beakperch Oplegnathus fasciatus.](http://oak.go.kr/repository/journal/16273/KSSHBC_2015_v48n2_207_f007.jpg)
![Frequency spectrum of broadband echo signals as a function of pitch angle for nine fish species. Changes in the spectrum patterns among nine fish species are due to the complex constructive and destructive interference occurring within the fish. This interference patterns and complexity contain the biological information of interest. (a) large yellow croaker Larimichthys crocea, (b) chub mackerel Scomber japonicus, (c) fat greenling Hexagrammos otakii, (d) konoshiro gizzard shad Konosirus punctatus, (e) whitesaddled reeffish Chromis notata notata, (f) black scraper Thamnaconus modesutus [K], (g) black rockfish Sebastes schlegeli, (h) goldeye rockfish Sebastes thompsoni, (i) striped beakperch Oplegnathus fasciatus.](http://oak.go.kr/repository/journal/16273/KSSHBC_2015_v48n2_207_f008.jpg)
![Frequency dependence on target strength (TS) as a function of pitch angle for nine fish species. Changes in the TS patterns among nine fish contain more clearly resolved information of interest related to fish classification. (a) large yellow croaker Larimichthys crocea, (b) chub mackerel Scomber japonicus, (c) fat greenling Hexagrammos otakii, (d) konoshiro gizzard shad Konosirus punctatus, (e) whitesaddled reeffish Chromis notata notata, (f) black scraper Thamnaconus modesutus [K], (g) black rockfish Sebastes schlegeli, (h) goldeye rockfish Sebastes thompsoni, (i) striped beakperch Oplegnathus fasciatus.](http://oak.go.kr/repository/journal/16273/KSSHBC_2015_v48n2_207_f009.jpg)