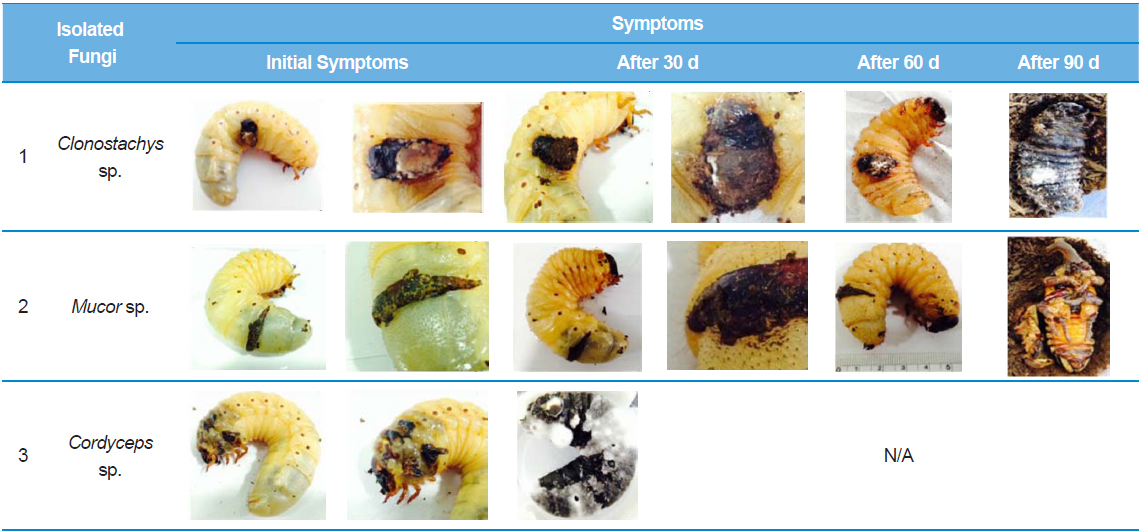
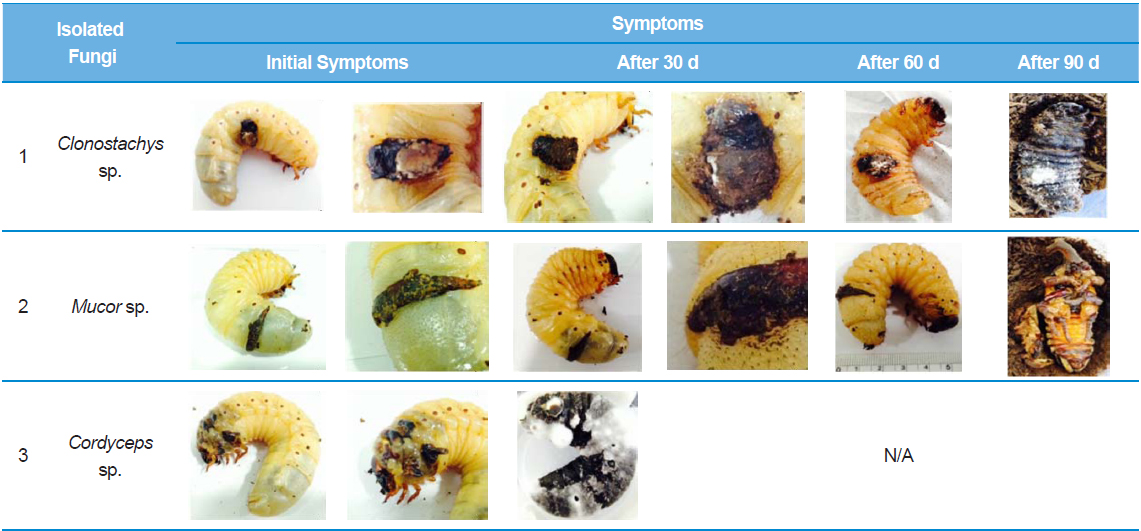
Recently in Korea, there has been an increase in farms breeding insects for use as food, medicine, animal feed, or pets. In particular, there has been an increase in the number of farms mass breeding rhinoceros beetles (Allomyrina dichotoma) for use as medicine in China and South Korea, as they have anti-oxidant and anti-hepatofibrotic properties (Kim 2005; Koyama et al. 2006; Sagisaka et al. 2004; Suh et al. 2010). However, insect breeding requires sufficient space for growth and regulation of the number of insects being bred. Hence, with the industrialization of insect breeding, there is a need for continuous monitoring and prevention of diseases and pathogens (Papp et al. 2014). In the present study, we investigated the process by which mass breeding of insects results in overcrowding, which leads to larval cannibalism and subsequent secondary fungal infection in A. dichotoma. Opportunistic fungal infections, such as those caused by entomopathogenic fungi, arise inherently in plants and even during hibernation in insects (Assaf et al., 2011), and such fungal infections are the major cause of fatalities in hibernating insects (Kubatova and Dvorak 2005). In the present study, the main cause of infection seems to be saprobic fungi in the sawdust that infects the epidermis. However, secondary infection caused by Cordyceps sp. on the wounds in the larval epidermis lead to death of larvae within one month. Cordyceps sp. are primary entomopathogens, with the fungal hyphae entering the mouth or epidermal spiracles of the insect, growing throughout the whole tissue, and replacing it with fungal mycelium, while the cuticle supports the fungal stroma, which cover the entire epidermis (Dworecka-Kaszak 2014; Hodge et al. 1996). The medicinal value of Cordyceps sp. has been revealed by various previous studies(Cho et al. 2006; Dworecka-Kaszak 2014).
Clonostachys sp. are endophytic fungi. Endophytic fungi infect plants, but the plants do not show any symptoms. Endophytes are used by the plant for defense against other phytopathogens, nematodes, and insects. They also interact with other phytopathogenic bacteria and viruses to protect the plant (Tan et al. 2012; Zabalgogeazcoa 2008). According to (Muvea et al. 2014), Clonostachys rosea(Ascomycota: Hypocreales) acts as an entomopathogenic fungi and reduces the survival of Thrips tabaci. This effect promotes plant growth and inhibits plant pathogens. T. tabaci laid the lowest number of eggs on C. rosea inoculated onions.
Phytoprotective endophytic fungi that inherently infect plants are directly or indirectly pathogenic in insects. This phenomenon was also confirmed in the present study, where endophytic fungi that infected A. dichotoma did not cause death within one month like Cordyceps sp. and other primary entomopathogenic fungi, but caused death of the larvae before pupation. Mucor sp. are also entomopathogenic fungi that cause mortality in hibernating insects. In the present study, larvae infected by Mucor sp. did pupate, but ultimately died. However, Mucor sp. are generally not considered entomopathogenic (Kubatova and Dvorak 2005). Moreover, there are no studies that confirm Clonostachys sp. as entomopathogenic fungi.
Among the stage-3 A. dichotoma larvae raised at a farm in Chungcheongnam-do, South Korea, we received larvae with black spots. The A. dichotoma farm used a mass breeding method, with 50–60 stage 3 larvae placed in a 420 × 240 × 230-mm breeding cage, and the temperature controlled at 25–29°C. However, the A. dichotoma at this breeding farm were procured from other regional farms, so the not all insects were raised at this farm from their egg stage. The stage-3 infected A. dichotoma larvae with black spots larger than 0.5 × 0.5 cm were placed in transparent, circular plastic containers with a 9-cm diameter and 7-cm height, at a temperature of 25°C and 60% humidity. Each container contained 1–2 larvae that were observed for 90 d as they pupated and matured.
We obtained spores from the black spots on the larval A. dichotoma epidermis, and transferred them to potato dextrose agar (PDA) (Difco, USA) plates. The spores were cultured for 10 d in a 24°C incubator, and then individual spores were isolated. We purified genomic DNA (gDNA) from the isolated fungi using the soil genomic DNA purification kit (MO Bio, USA).
In order to identify the isolated strains using molecular biology, we performed PCR amplification of the internal transcribed spacer (ITS1-5.8S-ITS2) using ITS1 (5'-TCCGTAGGTGAACCTGCGG-3') and ITS4 (5'-TCCTCCGCTTATTGATATGC-3') primers. We performed electrophoresis of the products of PCR amplification using a 1% agarose gel, and then purified the samples using a gel extraction kit (Qiagen, Germany). The purified PCR products were ligated to pGEM-T vectors (Promega, USA). Base sequence analysis was outsourced to Macrogen Co. (Seoul, Korea), and the results were compared to gene sequences using the National Center for Biotechnology Information (NCBI) Basic Local Alignment Search Tool (BLAST). We composed a catalogue of the fungi isolated from each A. dichotoma larva.
For the morphological observations of the Clonostachys sp. lines isolated from A. dichotoma larvae that were unable to pupate, we mounted them on a 200-mesh Ni grid and used an H-7000 Hitachi microscope (Hitachi, Japan) to observe the specimens by scanning electron microscopy (SEM) at 10 kV and 7.5 kV.
Among the A. dichotoma larvae with black spots infected by Clonostachys sp., those larvae that could not pupate showed an increase in spot size caused by fungal settlement. The larval epidermis became hard and dull, before turning yellowish, gradually becoming brown-yellow, and then turning black. The black spots on the infected A. dichotoma larvae did not show a considerable change in black spot size, but the spot thickness increased by approximately 0.3 cm.
We first isolated and identified fungi from larvae infected by Clonostachys sp. These larvae did not die, despite a rapid rate of fungal proliferation, but could not pupate. To identify the selected strains using molecular biology, we amplified the ITS region and ligated it into T-vectors, before determining the base sequence. This showed 100% homology with Clonostachys sp. 18SrRNA. It also showed 98% homology with the 18SrRNA partial sequence of C. rosea , which is an entomopathogenic fungus in Oncometopia tucumana and Sonesimia grossa (Hemiptera: Cicadellidae) (Toledo et al. 2006). However, larvae infected by Mucor sp. or Cordyceps sp. did not show a change in the size of the black spots or lesions. Mucor sp., in addition to Acremonium, Aphanocladium, Engyodontium, and Morierella are not considered to be entomopathogens in all insect species. Larvae infected by Mucor sp. showed no change in the size of the black spots, and were able to pupate, but ultimately died as pupae. According to (Kubatova and Dvorak 2005), when Mucor hiemalis Wehmer is infected to an insect’s wounds, M. hiemalis penetrates and infects the wounds of the insect. These fungi can also be seen as opportunistic entomopathogens. Larvae infected by Cordyceps sp. died within 30 d of appearance of black spots, and although the change in spot size was relatively small compared to other fungi, the time to death was faster, demonstrating fatality due to a primary entomopathogenic fungus. A. dichotoma larvae infected by Clonostachys sp. generally died without pupating compared to other larvae, but the time to death was approximately 3-fold longer than larvae infected by entomopathogenic Cordyceps sp. Approximately 70% of A. dichotoma larvae with black spots due to lesions pupated, but they all died as pupae, and were unable to eclose as adults. The black spots on the larvae were possibly the result of cannibalistic wounds from overcrowded breeding conditions. The black spots appeared to be dead cells, and the size of the spots did not increase. For more precise research on these black spots, confirmation of the course of disease progression is required, by evaluating the extent of the lesions and fungal infections of the epidermis through histological staining (Gotoh et al. 2008), as well as by investigating fungal infections of epidermal cells and hemolymph (Ortiz-Urquiza and O. Keyhani 2014; Zhao et al. 2014). Based on the above results, overcrowded breeding of A. dichotoma increases the risk of secondary fungal infection through cannibalistic lesions, with a 90% larval mortality rate. In the preliminary bioassay, to test the pathogenicity of Clonostachys sp., we used 2-stages of A. dichotoma larvae and sprayed 1 × 107 fungal spores 3 times at 1-h intervals, and after 3 wk, the mortality in non-wounded, infected larvae was 33%. For this mortality rate to have validity, a bioassay needs to be performed under identical conditions to determine whether the death of pupae is the result of secondary fungal infection, or physical deformity resulting from cannibalistic wounds. In addition, because various entomopathogenic microorganisms and Acarina, inherent to sawdust (Navarro-Campos et al. 2012), lie dormant in A. dichotoma, other microorganisms or chemicals (Schwarz et al. 2012) are required to prevent this from causing disease in insects. Thus, overcrowded breeding of A. dichotoma larvae should be avoided, and a breeding environment suited to A. dichotoma ecology needs to be determined.