Average temperatures have risen globally over the past century, with the most pronounced and rapid changes at high altitudes (IPCC 2007, 2013). Within these zones, treeline positions are widely thought to be temperature sensitive and potentially responsive to a warming climate (Lenoir et al. 2008, Harsch et al. 2009, Kreyling et al. 2010, Kullman 2010a). For this reason, the dynamics of the upper altitudinal or latitudinal treeline have been studied around the globe with the aim of detecting change, understanding responses to temperature variation, and evaluating the threat to alpine biota in response to treeline movement (Kong 1999, 2000, Walther 2003, Holtmeier and Broll 2005, Ihm et al. 2007, 2012, Case and Duncan 2014, Hagedorn et al. 2014, Smith-McKenna et al. 2014). Harsch et al. (2009) reported a global dataset of 166 sites for which treeline dynamics had been recorded since AD 1900: advance was recorded at 52% of sites with only 1% reporting treeline recession. Recent treeline expansion has been reported for many locations around the world: Russia (MacDonald et al. 2008), Asia (Zhang et al. 2001), Europe (Kullman 2001), India (Singh et al. 2012), North America (Szeicz and MacDonald 1995) and New Zealand (Wardle and Coleman 1992). An assessment of treelines for the Swiss Alps (Gehrig-Fasel et al. 2007, Díaz-Varela et al. 2010) found a decadal increment of 32 m of mean altitudinal increment for a 12-year period. The record of temporal treeline dynamics on a slope in the Austrian central Alps (Wallentin et al. 2008) indicated a decadal advance of 28 m for the maximum elevation of the treeline and 17 m for the mean elevation in the period 1954-2006. Feeley et al. (2011) in a study of elevational shifts of the 38 Andean tree genera reported a mean migration rate of 2.5 m yr−1.
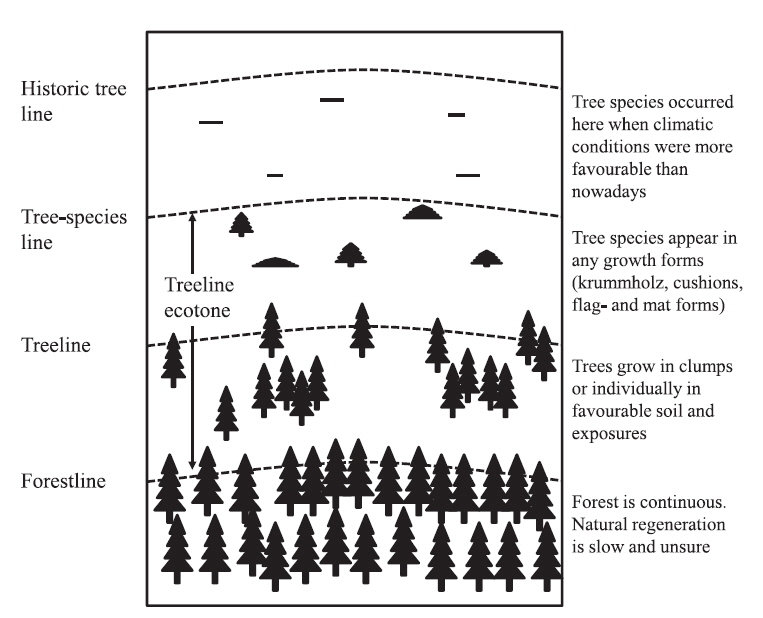
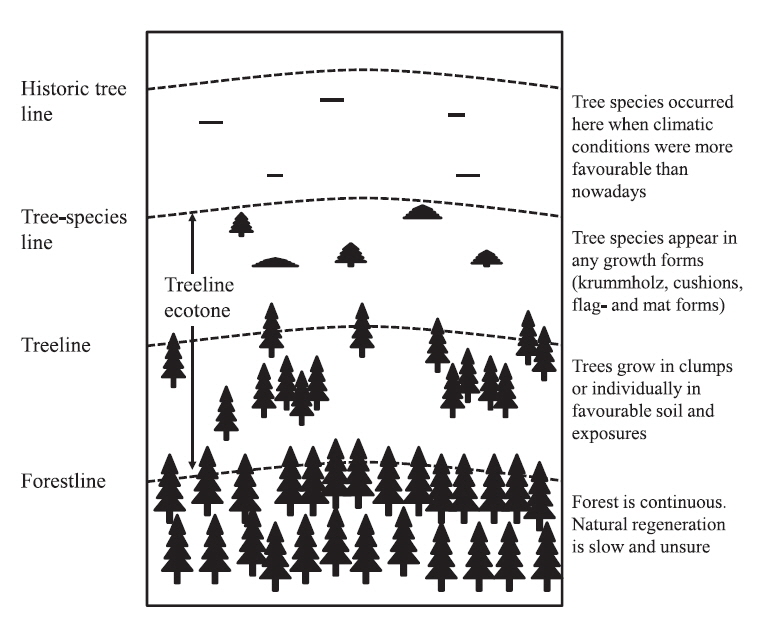
The tree limit of upright tree growth is defined as the line connecting the uppermost upright trees with a minimum height of 2 m (Holtmeier 2003). This definition of tree limits captures three important vegetation boundaries: “treeline” (the point where trees disappear), the “forestline” (the upper limit of closed-canopy forest) (Wieser et al. 2009, Harsch and Bader 2011), and the “treeline ecotone” (the transition zone from the uppermost closed forest to treeless subalpine and alpine vegetation)(Fig. 1) (Körner and Paulsen 2004, Autio 2006, Fajardo et al. 2011, Kim 2012, Körner 2012). Four primary treeline forms have been distinguished (Kong and Watts 1993, Chang et al. 1998, Cuevas 2000, Moen et al. 2004, Harsch et al. 2009, Harsch and Bader 2011, Green and Venn 2012): diffuse, abrupt, island and krummholz. Treeline form indicates the relative dependence of tree performance on various aspects of the external climate and other environmental factors (especially summer warmth versus winter stressors) and on internal feedbacks, thus allowing inferences on the type as well as the strength of climate-change responses (Harsch and Bader 2011). For example, diffuse treelines may be more responsive to warming because they are more strongly growth limited. In contrast, abrupt or krummholz treelines may be more strongly influenced by stress factors associated with winter conditions that lead to plant damage and limit survival. Krummholz form, characterized by a stunted habit, is commonly attributed to damage associated with factors such as wind abrasion, snow and ice damage. Hence, advance in krummholz and abrupt treelines may occur only when winter warming is sufficient to ameliorate other constraints, or when temperatures increase sufficiently to compensate for those constraints.
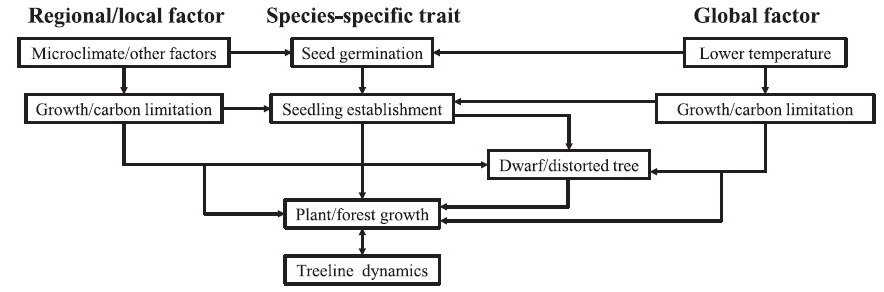
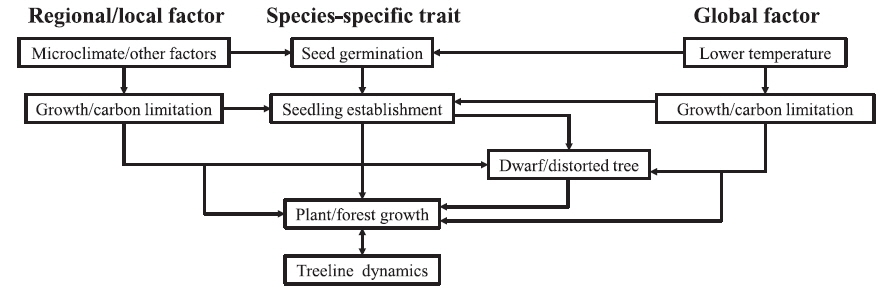
A range of regional and local scale factors will affect the overall impact of global scale climate drivers (Grabherr et al. 1994, Körner 1998, Jobbagy and Jackson 2000, Cullen et al. 2001, Dullinger et al. 2004, Walther et al. 2005). Assessment of the response of the treeline to environmental variability at regional and local scales is complex (Batllori et al. 2009) and requires a solid organizational framework. We summarized 12 categories of abiotic and biotic factors as 4 groups of global, regional, and local factors, speciesspecific traits, and positive feedbacks to differentiate the effects of each factor on the treeline positions (Fig. 2 and Table 1). Of prime importance are global factors related to climate. The thermal limitation of either carbon uptake (photosynthesis) or carbon investment (growth) can be explained by limitations dictated by a globally common isotherm: (1) growth limitation (Tranquillini 1979, Körner 1998, Shi et al. 2006) and (2) carbon limitation (Schulze et al. 1967, Stevens and Fox 1991). Secondarily, there are six regional and local factors related to treeline dynamics: (1) frost stress (Körner 1998, Körner and Paulsen 2004), (2) topography (Brown 1994a, 1994b, Leonelli et al. 2011), (3) water stress (Grace 1989, Richardson and Friedland 2009), (4) snow (Walsh et al. 1994, Gottfried et al. 2011), (5) wind (Holtmeier and Broll 2007, Richardson and Friedland 2009),(6) fire (Shankman and Daly 1988, Stueve et al. 2009), and (7) disturbance (Gehrig-Fasel et al. 2007, Tomback and Resler 2007). Thirdly, species-specific traits deal with the effects of treeline species on treeline positions including reproduction (Sveinbjörnsson et al. 1996, Körner 1998) and life history traits (Szeicz and Macdonald 1995, Motta and Nola 2001). Finally, there are positive feedbacks in structuring the dynamics of treelines (Wilson and Agnew 1992, Malanson et al. 2011).

Global, regional, and local factors, species-specific traits, and positive feedbacks determining treeline positions in high altitude environments
The most likely explanations for the formation of alpine treelines emphasize the temperature-factor characteristic of higher altitudes such as growing season length and summer temperatures (Table 1 and Fig. 2). Any common treeline theory needs to account for higher- and lower-latitude conditions, but the traditional view of temperature-controlled tree growth at upper elevations is largely based on high-latitude treelines (Handa et al. 2005, Shi et al. 2008). The treelines of the world’s mountains seem to follow a common isotherm, but this is only supported by indirect evidence. Actually, high altitude climatic treelines are associated with a seasonal mean ground temperature of 6.7-13.0°C (Cogbill and White 1991, Körner and Paulsen 2004). This temperature-treeline relationship emphasizes the length and mean temperature of the growing season (Hättenschwiler and Körner 1995, Körner 1998, Sveinbjörnsson 2000, Daniels and Veblen 2003, Hoch and Körner 2003, Lloyd and Fastie 2003, Camarero and Gutierrez 2004, Wang et al. 2006). Air and soil temperature has recently been identified as the most important factor in controlling the treeline positions across the world (Körner and Paulsen 2004, Leonelli et al. 2011). Seasonal mean temperatures are higher (7-8°C) in the temperate and Mediterranean zone treelines, and are lower in equatorial treelines (5-6°C) and in the subarctic and boreal zone (6-7°C). While air temperatures are higher than soil temperatures in warm periods, and are lower than soil temperatures in cold periods, the daily means of air and soil temperatures are almost the same at 6-7°C, a physics driven coincidence with the global mean temperature at treelines (Körner and Paulsen 2004). This translates to low soil temperatures that reduce soil nutrient availability by lowering microbial soil mineralization rates and nitrogen fixation and by reducing nutrient uptake capacity and reducing root growth and thus their exploitable soil volume (Moen et al. 2008). Two hypotheses have been put forward to link temperature, plant metabolism and treeline positions: (1) the growth (i.e., sink) limitation hypothesis (Körner 1998), a sink limitation through thermal limitation of meristematic activity with direct consequences for structural growth, and (2) the carbon (i.e., source) limitation hypothesis (Stevens and Fox 1991), a source limitation through insufficient net carbon acquisition due to low daytime temperatures.
The commonly accepted hypothesis is that growth at a treeline is limited by temperature (Hoch and Körner 2003, Shi et al. 2006, Susiluoto et al. 2007, Shi et al. 2008, Hoch and Körner 2009). Körner (1998) argued that the development of new tissues in treeline trees are less likely to be limited by the supply of photosynthetic assimilates, but rather by the rate at which the products can be utilized. This is known as the growth limitation hypothesis. In a comparison with different treeline
Carbon limitation because of a shortage of photoassimilates has long been regarded as the key to explain the upper altitudinal or latitudinal treeline on a global scale (Schulze et al. 1967, Stevens and Fox 1991). The carbon limitation hypothesis draws on the carbon balance of photosynthesis and respiration (Körner 2003). Körner (1998) hypothesized that carbon gain is restricted by the growing season lengths, whereby low temperatures and short growing periods limit photosynthesis to the point that it is exceeded by respiratory demand. To date, there has been little evidence in support of the carbon limitation hypothesis, although no direct manipulative test has been carried out. A recent study did not find evidence for carbon processing (sink) limitations, but rather photosynthetic carbon gain (source) limitations in Himalayan trees at the treeline (Li et al. 2008). Also, significant reductions in photosynthetic carbon gain in seedlings just older than the pre-establishment life stage (>3 years old) have been reported (Johnson et al. 2004), although limitations in carbon gain versus processing have not been differentiated. Meanwhile, the photosynthetic carbon uptake in treeline trees does not appear to be highly sensitive to temperature (Shi et al. 2008). Furthermore, studies of the mobile carbon pool, measured as non-structural carbonates and lipids, in trees across an altitudinal and latitudinal transect at the treeline have shown that the accumulated carbon reserves are not lower at high elevations compared to low elevations (Hoch and Körner 2003, Körner 2003, Handa et al. 2005).
The second suite of factors determining treeline positions operate at regional and local scales (Walsh et al. 1994, Daniels and Veblen 2003, Körner and Paulsen 2004, Malanson et al. 2007, Leonelli et al. 2011). All these factors interact with and, in some cases, supersede the influence of climate to explain treeline positions (Table 1 and Fig. 2). Thus, research on climate impacts on treelines at regional and local scales needs to identify multiple potential sources of variation in the structure and dynamics of treelines (Daniels and Veblen 2003).
Frost, frost desiccation or phototoxic effects may contribute to treeline formation (Körner 1998, Berdanier 2010). Frost damage does not threaten tree survival in the temperate zone treelines, but may lead to distorted growth by causing injury damages (Tranquillini 1979, Körner 2012, Rixen et al. 2012). Frost desiccation occurs during late winter when the soil is frozen but skies are clear and solar radiation is high. As exposed frosted foliage warms in direct sun, a strong vapor pressure deficit is created, evapotranspiration from the leaf is high, and desiccation occurs (Sakai 1970, Richardson and Friedland 2009). Frost desiccation is mainly observed in young trees and diminishes with age and size of branches or trees (Körner 1998). Although this frost desiccation may be a problem for young trees above the treeline in some parts of the temperate zone, this factor does not appear to be widespread for established trees.
The role of topographical factors in controlling future treeline positions is manifest in the interaction of climate with elevation, aspect and soil properties at higher altitudes (Brown 1994a, 1994b, Gottfried et al. 1999, Walsh et al. 2003, Butler et al. 2007, Bader and Ruijten 2008, Leonelli et al. 2009, 2011, Scherrer and Körner 2011) (Table 1 and Fig. 2). Mountains are characteristically conical in shape, and climate change impact scenarios usually assume that a smaller surface area will be available as species shift to higher elevations. However, as the frequency distribution of additional physiographic factors (e.g., slope angle) changes with increasing elevation (e.g., fewer gentle slopes available at higher elevation), upslope migrating species will encounter increasingly unsuitable conditions (Pauli et al. 1996, Guisan and Theurillat 2000). The masselevation effect describes variation in the treeline based on mountain size and location and was introduced to account for the observed tendency for temperature-related factors such as treeline and snowline to occur at higher elevations in the warmer and drier continental climate of the inner regions than on their outer margins (Odland 2009, Leonelli et al. 2011). Under warmer temperature conditions, treeline shifts are therefore expected to be more evident in the inner regions since the treelines are more likely to shift upward into the alpine environments (Leonelli et al. 2011). Resler (2006) highlights the importance of surface geomorphic features, specifically terrace risers, increasing favorable local site conditions, largely by protecting seedlings from wind in the study of the role of surface geomorphic features in tree establishment at the alpine treeline in Glacier National Park, Montana. The sheltering effect of surface features enables initial seedling establishment, and in some cases survival, above current treeline locations, thereby initiating a positive feedback effect that encourages subsequent tree establishment. Geomorphic features are therefore important in linking scales of patterns and positions at the alpine treeline.
The effects of water stress limiting the altitude of a treeline has been reported (Marchand and Chabot 1978, Cochrane and Slatyer 1988, Rada et al. 1996, Cairns and Malanson 1998, Smith et al. 2003). Decreasing moisture contents and osmotic potential values at wind-exposed treeline during the winter months can be explained by the fact that at such sites the soil remains frozen to a depth of 1 m. The soil thaws near the surface only at the end of April, but later in May it thaws out to a greater depth (Aulitzky 1961, Baig and Tranquillini 1980). Plants wintering on such sites are unable to absorb soil moisture, thus their survival depends upon tissue water reserves and drought resistance.
The proportion of trees suffering serious snow load damage has been reported to be 15-76% at the treeline (Walsh et al. 1994, Autio and Colpaert 2005, Bebi et al. 2009), so the damage of this kind is a key factor controlling the treeline positions. Growing season length and moisture can be affected by snow cover. In some places, snow will suppress the treeline by shortening the growing season and in others, snow encourages an upward expansion of the treeline along the elevation gradient by providing more water (Walsh et al. 1994). But these factors alone cannot explain why snow-free ridges at high elevations do not have trees, nor why some tropical mountains with little or no annual snowfall still have treelines (Hättenschwiler and Smith 1999, Richardson and Friedland 2009, Barbeito et al. 2012). Snowload also plays an important role in the protection of young trees from frost and wind damage in treelines (Holtmeier 2003, Holtmeier and Broll 2007, Devi et al. 2008). In this case, snow accumulation around trees might induce an insulation and protection from injurious climatic effects, which may allow establishing seedlings to grow into successful trees. In contrast, long-lying snow may curtail the growing season and increase snow fungus infection (
Fire can affect local or regional treelines in several ways. Firstly, fire can cause the depression of altitudinal limits of treelines through the physical destruction of stands of trees (Wilson and Agnew 1992, Noble 1993). Under this scenario, particularly severe fires propagated in flammable treeless alpine environments or the treeline forests themselves can cause the mortality of large populations of trees at the treeline. For example, fires in the Colorado Front Range (Shankman and Daly 1988) and the Cascades National Park (Hemstrom and Franklin 1982, Stueve et al. 2009) of the western USA, severely burned extensive areas of high-elevation forest, effectively lowering the existing treeline. Secondly, the destruction of seedlings established upslope by fire may prevent the advance of treelines. This mechanism is exacerbated by the slow growth rates of seedlings at higher altitudes such that they cannot escape the fire trap (Murphy and Bowman 2012). Thirdly, the removal of upslope vegetation cover may reduce competition pressure and provide substrate for seedling establishment and treeline advance (Noble 1980, Green 2009). The impacts of fire on treelines are particularly relevant to regions with a climate and vegetation conducive to fire such as the Rocky Mountains of North America, Patagonia in South America and the Australian Alps. Anthropogenic climate change has the capacity to alter fire regimes fundamentally through changing seasonal patterns of temperature, wind and precipitation, especially the occurrence of extreme weather events (Scott et al. 2014). Clearly, any consideration of the effect of global climate change on treelines will have to factor in a possible upregulation of fire in flammable alpine environments and their effect on tree populations and seedling recruitment.
Natural disturbance regimes related to herbivory, fungal pathogens, grazing and human disturbance such as land use changes and fragmentation are significant factors determining treeline dynamics (Cullen et al. 2001, Daniels and Veblen 2003, Batllori and Gutiérrez 2008, Brown 2010, Leonelli et al. 2011) (Table 1 and Fig. 2). Losses of a dominant treeline species by invasive forest pathogen may result in changes in vegetation patterns at the treeline (Harvell et al. 2002, Tomback and Resler 2007). Herbivores can limit the treeline below its potential at the landscape scale (Cairns and Moen 2004, Speed et al. 2010, Herrero et al. 2011). The presence of large numbers of small trees above the current treeline at a site in northern Sweden that experiences limited reindeer populations (
A possible explanation for treeline changes is land use change (Bolli et al. 2007, Gehrig-Fasel et al. 2007, Macek et al. 2009, Chauchard et al. 2010). Most upward shift activities were found to occur below the potential regional treeline (Gehrig-Fasel et al. 2007). Only 4% of the upward shifts were identified to rise above the potential regional treeline, thus indicating climate change. Land abandonment was the most dominant driver for the establishment of new forest areas, even at the treeline ecotone.
Seedling establishment and subsequent growth are necessary for the formation of new forests at higher altitudes, and both appear to be particularly challenging for tree species in the upper treeline ecotone (Black and Bliss 1980, Daly and Shankman 1985, Butler et al. 1994, Cho 1994, Hättenschwiler and Smith 1999, Germino et al. 2002, Kullman 2002, Smith et al. 2003, 2009, Gworek et al. 2007, Batllori et al. 2009, Kullman 2010b) (Table 1 and Fig. 2). Regeneration success determines whether the treeline shifts or remains static in response to environmental changes. Seed development, dispersal, germination, and seedling establishment are all limited by cold temperatures at the treeline (Körner 1998, Hättenschwiler and Smith 1999, Danby and Hik 2007, Holtmeier and Broll 2007, Dang et al. 2009). Some strategies such as extensive natural layering among
The life history traits of tree populations at treelines have been closely linked to differences amongst species in relation to the influence of low summer temperatures on growth rates, phenology, cover/density and development of trees (Motta and Nola 2001, Grace et al. 2002, Gamache and Payette 2004, Kullman 2007, Macek et al. 2009, Hertel and Schöling 2011, Xu et al. 2012, Anadon-Rosell et al. 2014) (Table 1 and Fig. 2). Understanding the interaction between environmental factors contributing to treeline formation and how these factors influence different life stages remains a major research challenge (Barbeito et al. 2012). This studies of the spatial and temporal dynamics of tree mortality and growth at treelines in the Swiss Alps provide experimental evidence that tree survival and height growth require different environmental conditions and that even small changes in the duration of snow cover, in addition to changes in temperature, can strongly impact tree survival and growth patterns at treelines. Further, their results show that the relative importance of different environmental variables for tree seedlings changes during the juvenile phase as they grow taller.
THE ROLE OF POSITIVE FEEDBACKS
The role of positive feedbacks in structuring the spatial pattern and temporal dynamics of treelines must be considered in models that put forward treelines as indicators of climate and environmental change. Positive feedbacks arise when vegetation communities actively modify their environment in a direction that enhances its own growth and survival, and simultaneously hinders or constrains other vegetation types (Wood et al. 2011). Positive feedbacks are emerging as pivotal controls of the distribution of alternative stable states (Scheffer et al. 2001) of plant communities at a variety of spatial scales from herbaceous sea beds, to regional dichotomies of treeless and woody vegetation in the temperate zone (Warman and Moles 2009, Odion et al. 2010, Knox and Clarke 2012, Wood and Bowman 2012), to savannahs and closed forests across the tropics (Hirota et al. 2011, Mayer and Khalyani 2011, Staver et al. 2011, Murphy and Bowman 2012). Identifying whether vegetation communities exist as alternative stable states is crucial because of the non-linear dynamics that dictate their past, present and future trajectories. Instead of gradual linear changes in response to gradual changes in climate, nutrient loading or habitat fragmentation, systems maintained as alternative stable states are characterized by lags in response to climate or abrupt switches to a contrasting state, as the resilience afforded by positive feedbacks breaks down (Scheffer et al. 2001).
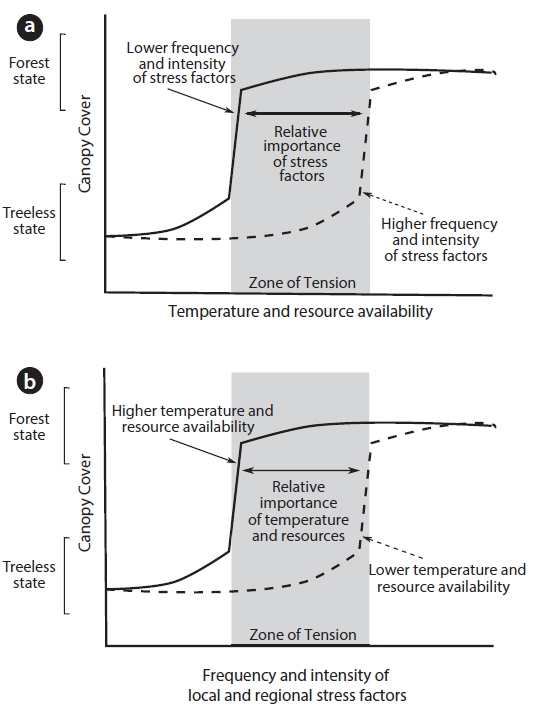
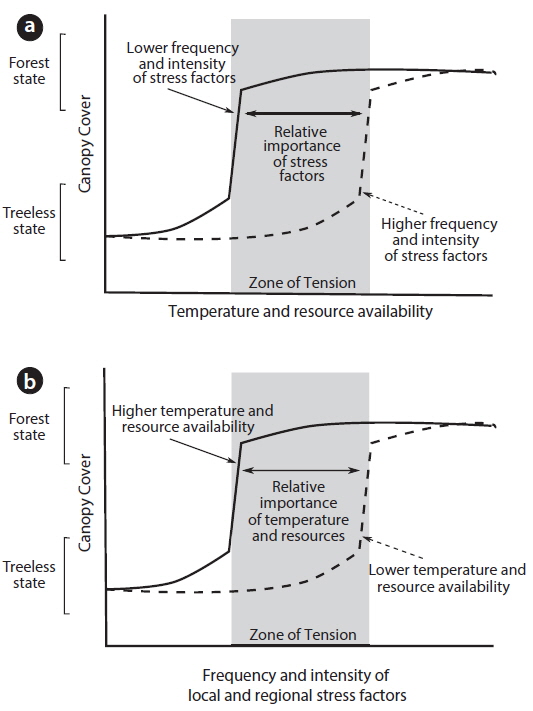
At the treeline, forest and treeless alpine vegetation communities can be considered as alternative stable states when the two opposing vegetation types are maintained by strong feedbacks driven by interactions between forest canopy cover, gradients of temperature and resource availability and the frequency and intensity of local stress factors such as wind, snow, frost and fire (Fig 3). Through canopy cover, trees are able to modify environmental conditions to promote tree establishment and growth such that abrupt treeline boundaries are not related to patterns in substrate, topography or temperature, but are brought about by plant interactions alone (Wilson and Agnew 1992, Malanson et al. 2011). Field and modelling studies have shown that by modifying their microclimate, tree canopies can promote the perpetuation of their community through a range of mechanisms including shielding of wind (Alftine and Malanson 2004, Holtmeier and Broll 2010), exclusion of fire (Bader et al. 2008), moderation of snow pack and frost events (Wilson and Agnew 1992, Bekker 2005, Batllori et al. 2009), accumulation of nutrients (Cairns 1999), or offering shade and protection from exposure to high levels of solar radiation (Ball et al. 1997, Germino and Smith 1999, Bader 2007).
The identification of positive feedbacks operating within treelines has important implications for their usefulness as barometers of climate and environmental change. These feedbacks ensure that an established forest can rejuvenate and persist even if the external climate deteriorates. Thus, treelines are not necessarily in equilibrium with climatic conditions and may exhibit a lagged response to climate change (Scheffer et al. 2001) (see zone of tension in Fig. 3). A further emergent property of alternative stable state ecosystems is abrupt shifts from one state to another (i.e., tree to alpine) once a threshold in external conditions is reached or due to a disturbance (Scheffer et al. 2001) (Fig. 3). To date, field observations of the temporal trends expected in systems operating as alternative stable states (i.e., lags and catastrophic shifts) are lacking for alpine treeline environments (and indeed, for most long-lived forest ecosystems), although simulation modelling approaches have been used to explore the effects of positive feedbacks on the temporal dynamics at the treeline (Malanson 2001, Alftine and Malanson 2004, Bader et al. 2008). Definitive evidence for treelines exhibiting the complex characteristics of alternative stable states is still nascent, but can be progressed though the adoption of the emerging research approaches undertaken in forest-treeless systems such as the spatial analyses of forests in tropical savannahs (Staver et al. 2011, Murphy and Bowman 2012) and the elucidation of temporal trends in vegetation states identified from sediments in temperate forests (Jeffers et al. 2011, Fletcher et al. 2014).
In general, treeline positions are the result of a combination of unfavorable conditions for tree regeneration, seedling establishment and tree growth (Table 1 and Figs. 2 and 3). The commonly accepted hypothesis is that growth at a treeline is limited by growing season temperature (Grabherr et al. 1994, Shi et al. 2006, Susiluoto et al. 2007, Hoch and Körner 2009). This growth limitation hypothesis is consistent with the correlation of tree growth with mid-summer temperatures, frequently observed at the treeline, and can be evoked to explain why the altitude of treelines is well correlated with July temperatures in the northern hemisphere (Körner 1998). The two factors determining the treeline positions are regional and local factors and species-specific traits. These factors interact with and, in some cases, supersede the influence of climate to explain treeline positions, structure and dynamics. Also, the role of positive feedbacks in decoupling the spatial pattern and process at the treeline from the underlying environmental variables is particularly important because it has implications for future trajectories of boundary shifts in the face of climate change. Thus, research on the climate change impacts on treelines at regional and local scales needs to identify multiple potential sources of variation in the structure and dynamics of treelines (Daniels and Veblen 2003). In order to estimate and predict the effect of climate change on upward advance of alpine treelines and to conserve and restore the treeline species, the following suggestions should receive particular attention in the future (Sveinbjörnsson 2000, Dullinger et al. 2004, Díaz-Varela et al. 2010, Malanson et al. 2011, Szerencsits 2012, Carlson et al. 2013). First, additional experimental studies are required to differentiate between the growth limitation and carbon limitation hypotheses. Such experiments must be carried out in a wide range of environments and for multiple species before robust generalizations are made. Second, further exploration is required to elucidate how positive feedbacks and speciesspecific traits may affect future range dynamics and how they interact with variation in regional and local climate trends. Third, an effort should be made to obtain better landscape scale predictors controlling treeline positions through the use of the rapidly expanding availability of remote sensed information. Fourth, further research initiatives in high mountain environments are needed to establish an effective projection and conservation system. This should include international and interdisciplinary cooperation, and include long-term monitoring, ecophysiological and phenological studies and predictive modelling. Ergo, there is a pressing need for an effective network of monitoring and conservation programs such as LTER (Long Term Ecological Research) or worldwide research collaborations such as GLORIA (Global Observation Research Initiative in Alpine Environments) to be established throughout the treelines.