영양염 농도, N/P비(nitrogen-phosphorus ratio), 유량, 광도, 수온 등은 조류의 천이에 영향을 주는 주요 환경요소이다(Jan et al., 2006). 낙동강에는 2012년에 8개의 보가 건설되어 대형 물리적 환경변화가 불가피 하게 되었다. 낙동강 살리기 사업으로 인한 가장 큰 변화는 저수용량의 증가와 유속 및 총인(TP) 농도의 감소이다. 유속감소는 조류성장을 유리하게 하여 낙동강 조류 대발생의 원인이 되었다는 비난에 직면해 있다. 정부에서는 조류발생 문제에 대응하여 총인 처리시설을 대폭 확대하였으나(Jun and Kim, 2011) 그 효과는 아직 밝혀진 것이 없으며, 조류성장을 억제하기에는 충분하지 않다고 인식하고 있는 실정이다.
조류저감을 위해 인위적으로 접근하는 일반적인 방법은 영양염 농도의 저감이다. 조류가 선호하는 영양염의 형태는 조류 종에 따라 서로 다른 것으로 알려져 있다. 규조류와 같은 진핵조류는 높은 질산염(NO3-N)을 좋아하는데 고수온기에 접어들면서 규조류가 약화되는 것은 질산염의 고갈과 관련이 있다고 했다(Gry et al., 2003). 남조류는 낮은 질산염 농도와 높은 암모늄(NH4-N) 농도를 좋아하며(Xia et al., 2011) 그래서 남조류에 있어서 NO3/NH4 비는 중요하다고 했다. 조류는 일반적으로 질소(N)와 인(P)를 Redfield 비율인 16:1로 섭취하는 것으로 알려져 있는데(Redfield, 1958) 이 비율은 조류의 평균 원소조성의 원자비율을 나타낸 것이며 영양염의 농도와는 무관하게 제시된 것이다. 남조류는 N/P 비가 15 이하의 낮은 것을 선호하며 질소고정 남조류는 N/P 비의 영향을 받지 않는다(Rita, 2007). Xia et al. (2011)은 N/P 비가 10 이하에서
수체의 안정성 변화는 환경인자의 변동 뿐만 아니라 조류군집 구조에 큰 영향을 미친다(Shitao et al., 2012). 유속이 빠르고 교란이 심하면 세포에 물리적인 손상이 발생하고 개체가 유광층에 노출되는 기회를 잃게 됨으로써 조류의 성장이 억제된다(Richmond and Vonshak, 1978). 많은 연구자들에 의해서 어느 임계치 이하의 속도에서는 조류의 성장이 촉진되는 것으로 보고되고 있다(Drapcho and Brune, 2000; Doyon et al., 2000; Richmond and Vonshak, 1978; Weissman et al., 1988; William et al., 1990). 즉, 적당한 교란은 세포와 수체 사이의 상대속도를 증가시킴으로써 세포 경계층 두께를 감소시켜 영양염의 전달율을 높여주며 (Marshall and Huang, 2010), 미소환경을 변화시켜 세포내분비물의 배설을 용이하게 하는 등(Li et al., 2012) 조류성장을 촉진시킨다는 것이다. Richmond and Vonshak (1978)은
그러나 그 동안 낙동강의 조류 군집구조 변화관련 연구는 중 · 하류에 집중되어 왔다(Chung et al., 1994; Joung et al., 2013; Moon et al., 2001; Son, 2013a; Son, 2013b). 최근의 연구로는 Yu et al. (2014)이 낙동강의 5개 보에 대해 수온, pH, 탁도, 적산일사량 등 일반항목을 중심으로 환경요인이 조류군집 구성에 미치는 영향에 대해 연구하였다. 본 연구에서는 낙동강의 구미보와 합천창녕보까지의 5개 보에 대해 2010년 5월부터 2013년 12월까지 수리 · 수문, 수질, 조류 군집분포 등의 조사 결과를 이용하여 영양염농도 변화 를 중심으로 환경요인이 조류의 종 조성 및 현존량 변화에 미치는 영향과 상관성에 대해 알아보고자 했다.
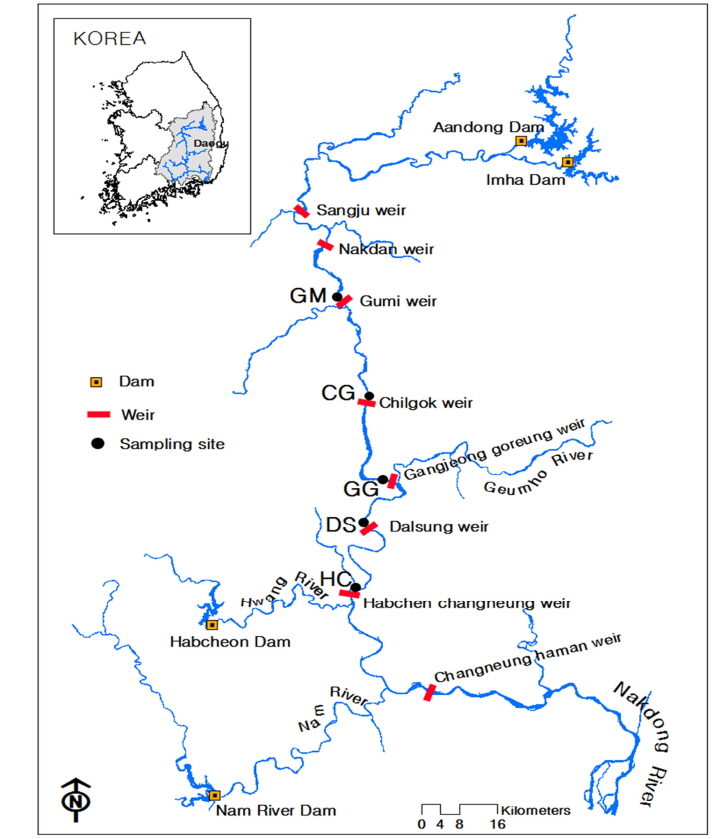
낙동강은 유역면적이 23,717 km2이며, 중 · 하류 지역에는 대형 오염원이 산재해 있고 하류에는 부산 · 경남 등의 취수원이 밀집되어 있어 오염에 대한 갈등이 심한 지역이다. 특히 하절기에 집중되는 강우는 그 양이 적어 상 · 하류 모두 양질의 수자원 확보문제가 현안이 되어 왔다. 2011년에는 낙동강 살리기사업 추진으로 상주보 등 8개 보가 설치되었다(Jun and Kim, 2011). 보 구간의 총 길이는 190.8 km이고 평균 구간거리는 23.8 km이다. 상주보가 13 km로서 가장 짧으며 창녕함안보는 42.9 km로서 가장 길다. 저류량은 총 569.4 백만m3 이고 상주보가 28.7 백만m3으로서 가장 적으며 창녕함안보가 127.1 백만m3으로서 가장 크다. 이번 연구의 조사지점은 Fig. 1에 나타낸 바와 같이 GM(구미보), CG(칠곡보), GG(강정고령보), DS(달성보), HC(합천창녕보) 등 5개 지점이며 현장조사 위치는 보 상류 2 km 지점이다. 조사시기는 2010년 5월부터 2013년 12월까지이며, 월 2회 조사를 실시하였다.
보 구간의 수리・수문 현황조사는 국가 수자원관리 종합정보 홈페이지(http://www.wamis.go.kr) 자료를 활용하였다. 수온, pH, 전기전도도, DO(dissolved oxygen), 탁도 등 물리적, 화학적 항목은 YSI 556 MPS를 이용하여 현장에서 측정하였다. 적산일사량(accumulated solar radiation; ASR)은 기상청 자료를 이용하였다. 수질 및 조류조사를 위한 시료는 2010년 5월부터 2013년 12월까지이며, 월초와 월말 등 2회로 나누어 채수되었다. 시료는 하천 중앙의 수심 0.5m 지점에서 Ruttner water sampler (Hydrobios, Germany, 1000mL)를 이용하여 4 L의 폴리에틸렌 용기를 이용하여 채수 하였다. 채수 시료는 냉온 보관하여 실험실까지 운반되었다. 조류분석을 위한 시료는 Lugol's solution으로 현장에서 고정하고 실험실로 옮겨 48시간 이상 침강시킨 후 상등액을 제거하고 5배 이상 농축하여 사용하였다. 동물플랑크톤을 위한 시료는 5 L 채취하여 64 μm sieve를 이용하여 100 mL로 농축하였으며 5% sucrose-formalin solution (Haney and Hall, 1972)으로 즉시 고정한 후 실험실에서 계수 및 동정하였다.
SS (suspended solid)는 Whatman GF/C glass fiber filter를 사용하였으며 105 ~ 110°C로 건조하였다. BOD5 (biochemical oxygen demand)는 20°C에서 5일 동안 식종희석법로서 분석하였으며 CODMn (chemical oxygen demand)는 과망간산칼륨법으로 분석하였다. 인산염인(PO4-P)과 총인(TP)은 아스코르빈산환원법 및 흡광광도법으로 분석하였다. 암모니아성질소(NH3-N), 질산성질소(NO3-N), 총질소(TN) 등은 각각 인도페놀법, 이온크로마토그래피법 및 과황산법으로 분석하였다. Chlorophyll a (Chl.a) 농도는 extraction methods를 사용하여 90% 아세톤(Wetzel and Likens, 1991)에서 흡광분석법으로 분석하였다. 조류시료는 농축된 1 mL를 10분 이상 침전시킨 후 광학현미경(M1, Carl Zeiss)을 이용하여 200배 ~ 1,000배로 검경하였다. 동물플랑크톤 분석은 SMZ-U(Carl Zeiss)와 M1(Carl Zeiss)으로 10 ~ 100배율 하에서 동정하였다.
자료의 변수의 비교는 정규분포의 경우는 일원배치분산분석법(ANOVA)으로 수행했으며, 그렇지 않은 경우는 KruskalWallis ANOVA를 이용해 수행되었다(SIGMASTAT, version 12). 환경인자와 조류개체 및 현존량과의 상호 관계는 Pearson’s correlation을 이용하여 평가하였다. 조류군집에 환경변수의 영향을 분석하기 위해 Ordination method의 선형 분석 기법인 RDA (redundancy analysis)가 수행되었다(CANOCO version 5.0).
조사기간 동안의 수질현황은 Table 1과 같다. 수온은 동절기에 0.6°C~1.5°C까지 강하하지만 하절기에는 31.6°C~34.3°C까지 상승하고 있어 뚜렷한 계절변동성을 보여주고 있었다. pH의 변화는 보별로 유사하였으며 7.0~9.7의 수준이었다. 지표수의 pH는 조류 개체의 성장에 강력한 영향을 주는 환경요소이다(Agrawal and Singh, 2000). 이번 조사에서 pH는 조류의 종수보다는 현존량과의 상관성이 0.45~0.55 (p<0.001)로서 높게 나타났으며 녹조류는 다른 분류군과 비교하여 종수 및 현존량과와 양의 상관성이 높게 나타났다(r = 0.32~0.43, p<0.001). Rao and Burns (1991)은 녹조류는 높은 pH에 내성이 강해서 알카리성 토양에 광범위하게 번식하고 있다고 보고하였다. TN과 TP는 5개 보에서 각각 1.397~6.552 mg L−1 및 0.020~0.373 mg L−1의 범위를 보여주고 있었다. 지점별로는 구미시 하수처리장 방류수가 유입되는 CG지점과 금호강이 유입되는 DS지점에서 급격한 증가가 있었는데, 이들 두 지점은 Kruskal-Wallis One Way ANOVA에 의한 차이검정 결과 통계적으로 유의한 차이가 나타났다(p<0.001). 구미시 하수처리장 유입 이후인 CG 지점과 GG지점, 금호강 유입 이후인 DS지점과 HC지점은 차이가 유의하게 나타나지 않아 낙동강의 영양염류는 구미시와 금호강으로부터 유입되는 부하량의 영향이 큰 것으로 판단이 된다. 조류의 현존량과 밀접한 관계가 있는 chl.a의 평균농도는 DS 지점이 31.0 mg m−3 으로 가장 높았다. DS 지점은 또한 조사기간 평균규조류 현존량이 4,712 cells mL−1으로 가장 높았고 Chl.a와 규조류 현존량과의 상관계수도 0.81 (p<0.001)로서 가장 높았다. 따라서 DS지점의 Chl.a의 농도가 높은 것은 다른 지점보다 상대적으로 높은 규조류의 현존량에 기인하는 것으로 판단된다.

Variation of average water quality concentration each survey site from May 2010 to December 2013 (n=87) in the Nakdong River
조사기간 동안의 조류의 분류군별, 지점별 종수 및 현존량 변화는 Table 2에 나타내었고 낙동강의 주요 조류종인 남조류와 규조류의 5개지점 평균 현존량 변화는 Fig. 2에 나타내었다. 규조류는 종수 및 현존량에 있어서 다른 분류군에 비해 가장 높게 나타났다. 규조류의 봄철 대발생은 2월에 시작하여 3월에 최고 수준에 도달하고 있었다. 규조류는 DS 지점이 4,712 244 cells mL−1로 가장 높았는데 DS 지점의 규조류 농도가 특히 높았던 것은 2013년 3월초의

Variation of species and abundance each algal genera and each site from May 2010 to December 2013 in the Nakdong River (mean ± standard deviation)
남조류는 현존량에 비해 종수는 지점별 평균 1.7~2.4로서 비교적 낮았다. 남조류의 연간 평균현존량은 영양염의 농도가 가장 높았던 DS 지점보다 HC 지점이 3,244 cells mL−1로 가장 높았으며 규조류와 다른 특성을 보이고 있었다. 종의 변화에 대한 보 방류유량과 유입유량의 영향은 HC지점이 DS 지점보다 높게 나타났다(Table 3). DS지점은 금호강 등 유입하천의 영향이 높았을 것으로 추정된다.

Correlation coefficients of weir influent and effluent with algal species and abundance from Aug. 2012 to Dec. 2013 (n=33)
하천의 수리 ・ 수문학적 요인은 환경인자의 변동과 조류분포에 많은 영향을 미친다(Shitao et al., 2012). 그러나 보 건설 이후 변화된 낙동강의 흐름특성이 조류분포에 어떠한 영향을 미치는지 정량적으로 규명하기 위해서는 많은 자료가 필요하나 본 연구에서는 자료의 한계가 있었다. 다만, 보별 평균유속은 저류량을 평균방류량으로 나누어 구할 수 있는데, 이렇게 구한 평균유속(m s−1)은 구미보 0.041, 칠곡보 0.043, 강정고령보 0.034, 달성보 0.067, 합천창녕보 0.091 등으로 나타났다(2012.8. ~ 2013.12., n=33).
보별 방류유량 및 유입유량이 조류 종 및 현존량에 미치는 상관성을 구한 결과 종의 수는 방류유량이나 유입유량에 대부분 양의 상관성을 가지고 있었다(Table 3). 이는 유입 또는 방류 유량으로 인해 상류 보의 조류개체가 하류로 확산되고 있음을 의미한다. 달성보에서는 유입 및 방류유량과의 상관성이 낮고 유의성이 없었던 것은 금호강의 유입영향이 크기 때문인 것으로 판단이 된다.
상류에 출현한 개체의 현존량 크기도 하류 종 분포에 영향을 주고 있었다(Table 4). 세포수(cells mL−1)가 0~50의 분포일 때는 하류에서 발견될 확률이 59%이었으나 150 이상일 때에는 92.8%로 나타났다. 이것은 낙동강 보의 유입 및 방류 유량이 조류의 확산에 양의 영향을 주고 있음을 말해준다. 조류 현존량은 Table 3에서 보는 바와 같이 유입 및 방류량 모두 음의 상관성을 보이고 있었으나 통계적 유의성은 없었다.

Detection probability of algae species in down stream weir each cell detected range of upper stream weir (n=435)
조류는 성장하면서 서로 다른 양의 영양염을 필요로 하는데, 가장 낮은 수준의 영양염이 먼저 고갈 되고 그 다음에 조류의 성장이 억제된다(Kedong, 2001). 조류가 영양염에 반응하는 것은 환경조건에 따라 일정하지 않은데, 낮은 유속과 높은 영양상태에서도 남조류의 대량증식 현상이 나타나지 않는 경우가 있으며, 낮은 영양상태에서도 남조류가 대량증식 하는 경우도 있다(Wei et al., 2010). White et al. (1991). 낙동강 유역에는 조류억제를 위해 총인 처리시설에 많은 투자를 했으나 조류성장 억제효과에 대해서는 아직까지 보고된 것이 없다. Fig. 3은 5개 조사지점의 41개월 동안의 TP농도에 대한 변화 곡선이다.
’11년의 5개지점 TP의 평균농도는 0.145 mg L−1이었으나 ’12년 및 ’13년에는 0.081 mg L-1 및 0.069 mg L−1 로서 ’11년보다 44.1% 및 52.4% 낮아졌으며 이는 총인처리시설의 확충에 기인하는 것으로 판단된다. 5개 지점의 TP 최소농도는 0.020~0.039 mg L−1 의 수준으로 나타났다.
TP 농도의 감소에 따른 Chl.a 농도의 변화는 Fig. 4에 나타내었다. 2011년의 5개지점 Chl.a 의 농도는 22.4 mg/m3이었으나 2012년 및 2013년에는 각각 22.7 mg/m3 및 25.9 mg/m3로서 ’11년보다 다소 높게 나타났다. 조류 현존량의 영향을 받는 Chl.a 농도의 변화는 계절성을 보여주고 있어 TP 농도변화 패턴과는 다르게 나타나고 있었으며, TP 농도의 감소에 따른 Chl.a 농도의 감소효과도 분명하지 않은 것을 볼 수 있었다. 다만, 하류의 HC 지점에서는 ’11년보다 다소 감소하고 있었다.
GM 지점에서 ’13년 8월 상반기의 남조류 수화현상은 TP의 농도가 0.148 mg L−1 까지 증가하여 TN/TP비가 11.2로 감소하였을 때 시작하여 TP 농도가 0.033 mg L−1 까지 감소하여 TN/TP비는 45.1로 증가하였을 때 거의 소멸되었다. GG지점의 ’13년 9월의 수화는 인의 농도가 0.253 mg L−1로 증가하고 TN/TP비가 7.7까지 낮아졌을 때 시작하여 인의 농도가 0.040 mg L−1까지 감소하고 TN/TP비가 41.7까지 상승하였을 때 수화현상은 거의 사라졌다. DS 지점에서는 ’13년 8월 상반기에 TP 농도가 0.191 mg L−1로 상승하고 TN/TP비기 11.6으로 낮아 졌을 때 수화현상이 시작되었고 TP 농도가 0.046 mg L−1로 감소하여 TN/TP비가 55.8이 되었을 때 수화현상은 거의 소멸되었다. 그러나 HC 지점의 남조류 수화현상의 시작과 종료시의 TP 농도와 TN/TP비가 다소 다른 특성을 보여주고 있었는데, ’13년 7월 하반기의 남조류 수화는 TP농도가 0.010 mg L−1로 낮고 TN/TP비가 35.6일 때 시작하여 이후 TP 농도가 0.001∼0.037 mg L−1정도로 낮을 때에도 수화현상은 지속되었다.
전체 조사기간 동안 TN 및 NO3-N과의 유의한 상관성은 나타나지 않았다. 남조류는 낮은 질산염 농도와 높은 암모늄(NH3-N) 농도를 좋아한다고 하였으나(Xia et al., 2011), 이번 조사에서 조류 분류군별 종 및 현존량에 있어 NH3-N과 양의 상관성을 보이는 것은 없었다. 이는 높은 값의 NO3/NH3 비에 기인하는 것으로 판단이 된다. 남조류는 암모늄을 선호하나 다른 형태의 질소원도 섭취를 한다(Isabel et al., 2003). 따라서 암모늄 농도가 낮은 것이 남조류 개체밀도 감소의 원인으로 작용하지는 않는 것으로 판단된다. NO3/NH3 비에 대해 양의 상관성이 유의하게 나타난 것은(p<0.05) 규조류의 종 및 현존량이었다.
TN/TP 비의 월평균 변동은 Fig. 4에 보여주고 있는데, 2011년에는 TN/TP 비가 14.7~44.0의 범위였으며 2012년에는 18.3~63.6, 2013년에는 23.7~95.5 으로서 변동성이 대단히 심했지만 겨울과 봄에는 높고 여름과 가을철에는 비교적 낮았다(Xia et al., 2011). 하절기에 TN/TP비가 특히 낮은 것은 강우유출에 의한 TP 부하의 증가에 기인하는 것으로 보인다. 높은 P의 농도, 그리고 낮은 TN/TP비는 남조류 대량증식에 유리한 것으로 알려져 있다(Schindler, 1997). Smith (1983)는 온대지방은 많은 호소 데이터를 분석한 결과, TN/TP 비는 29:1를 기점으로 하여 이것보다 작은 값에서는 남조류가 우점하고, 이것 보다 큰 값에서는 남조류의 대량증식이 없다고 하였다. Xia et al. (2011)도 낮은 TN/TP 비에서 높은 인 농도, 성층현상, 투명도 감소, 수온상승, pH상승 등은 남조류의 수화현상의 원인이 된다고 하였다. 낙동강에서 수화현상의 시작단계의 TN/TP비는 대부분 29:1 보다 낮았다. 그러나 HC 지점에서는 N/P 비가 35.6일 때 남조류 수화현상이 시작하기도 하였다.
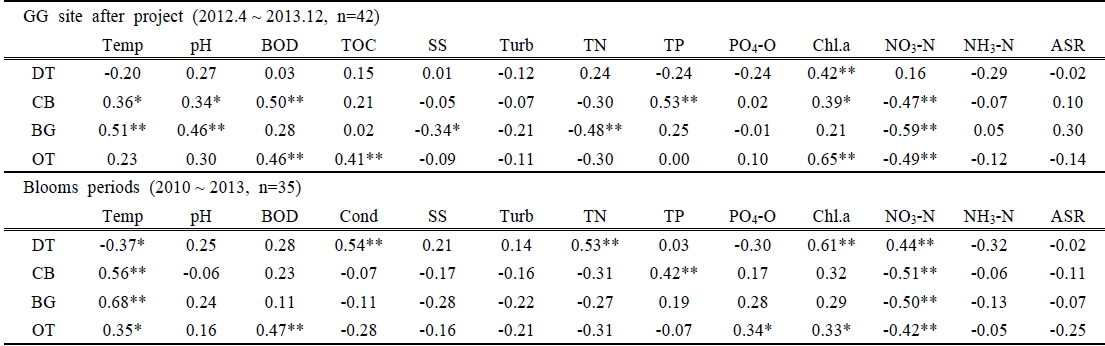
조사기간 동안 조류 개체수와 양의 상관을 나타낸 것은 수온과 pH, BOD, Chl.a 등이었으며 영양염 중에 TN 및 nitrate는 대부분 높은 음의 상관성을 나타내었다. Nitrate가 음의 상관성을 보인 것은 조류성장과 함께 nitrate의 농도는 감소하였으나 그럼에도 조류성장에는 충분하여 억제요인으로 작용하지 못했기 때문인 것으로 판단이 된다. 낙동강의 TP 농도는 감소하였으나 조사기간 동안 조류개체수는 TP와 유의한 상관성이 나타나지 않았다. 다만, 4대강 공사가 끝나고 담수를 시작한 이후부터(2012.4~2013.12, n=42)의 분석에서는 GG 지점에서만 TP와의 Pearson 상관계수가 남조류 0.53(p<0.001)으로 통계적으로 유의하게 나타났으며(Table 5), 전체 조사기간 동안 5개 조사지점의 수화가 발생한 시기(7회, 10주/회)의 상관분석(n=35)에서는 남조류 개체수와 TP와의 상관계수가 0.42(p<0.01)로 통계적 유의성이 강하게 나타났다(Table 5).
[Table 5.] Pearson correlation coefficients between the environmental valuables and algal abundances
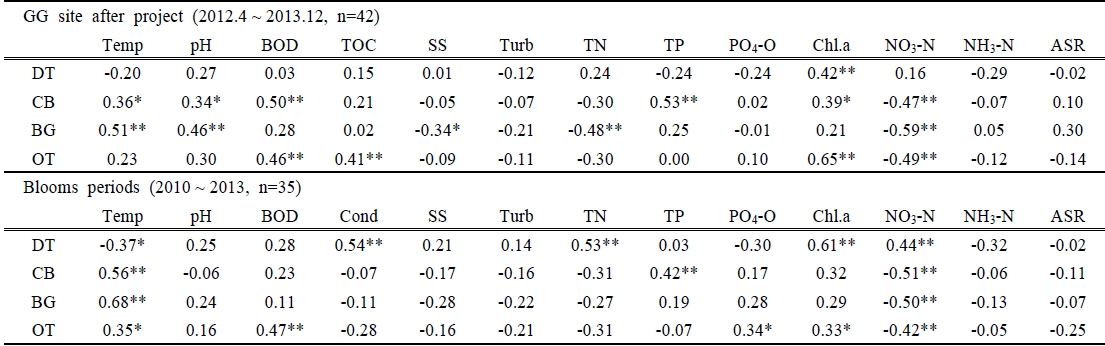
Pearson correlation coefficients between the environmental valuables and algal abundances
GG 지점의 TP 농도는 상류의 CG지점보다 2% 및 하류의 DS 지점보다는 50.0%가 낮지만, 남조류 현존량은 CG 지점보다는 49.4%, DS 지점보다는 14.4%가 높은 등 자체 영양염 농도 대비 조류 현존량이 높은 특성을 지니고 있다. 사업 이후에는 TP 농도가 0.030~0.050 mg L−1 의 범위에 해당하는 기간이 42.5%였으며 대부분 봄철 수화와 여름철 수화가 발생한 시기와 관련이 있었다. 이번 조사에서 낙동강의 수화 종료 시기에 있어서 TP 농도는 대부분 0.050 mg L−1로 나타났다.
GG 지점 및 수화발생 시기에 조류 군집변화와 환경요인과의 관련성을 찾기 위하여 RDA(Wollenberg, 1977) 분석을 실시하였다. 사업후 GG 지점의 RDA분석 결과에서(Fig. 5) axis1과 axis2는 종-환경요인 관계에 있어서 전체 변동의 89.8%를 설명하고 있으며, axis1과 axis2의 고유값(eigenvalue)은 각각 0.773과 0.813으로 나타났다. Axis1은 수온(r = −0.577), NO3-N (r = 0.640), TN (r = 0.537), pH (r = −0.460), ASR (r = −0.258) 등과 높은 상관성을 보였으며 axis2는 TP (r = −0.420), NH3-N (r = -0.343), Chl.a (r = 0.324), PO4-P (r = −0.335), 탁도(−0.207) 등과 상관성을 보였다(Table 6). 남조류 세포수는 axis1에 음의 관련성(r = −0.744)을 보였으며, 규조류는 axis2에는 양의 관련성(r = 0.773)을 보였다(Fig. 5). GG 지점의 RDA ordination에서 남조류, 녹조류 및 기타조류는 수온, TP, PO4-P, ASR 등과는 양의 관련성을 보인 반면, 총질소, nitrate, 탁도 등과는 음의 상관성을 나타내었다. 규조류는 총질소, nitrate, SS 등과는 등과는 양의 관련성을 보인 반면 수온, TP, PO4-P, 탁도, NH3-N 등과는 음의 관련성을 잘 나타내고 있었다(Fig. 5).

Summary of RDA between environmental factors and algal abundance in the Nakdong River for GG site (after project) and all site (blooms periods)
수화발생 시기의 RDA분석(Fig. 5)에서 axis1과 axis2는 종-환경요인 관계에 있어서 전체 변동의 91.5%를 설명하고 있으며, axis1과 axis2의 고유값은 각각 0.952와 0.897로 나타났다. Axis1은 수온(r = −0.888), nitrate (r = 0.757), DO (r = 0.491), 탁도(r = 0.466), SS (r = 0.428) 등과 상관성을 보였으며, axis2는 NH3-N (r = −0.490), 전기전도도(r = 0.377), Chl.a (r = 0.350), TP (0.1729) 등과 상관성을 보였다(Table 6). 남조류는 axis1에 음의 관련성(r = −0.935)을 보였으며, 규조류는 axis2에 양의 관련성(r = 0.619을 보였다(Fig. 5).
수화발생 시기의 RDA ordination에서 남조류와 녹조류는 수온, TP, PO4-P, ASR 등과는 양의 관련성을 보인 반면, 총질소, nitrate, 탁도, SS 농도, 전기전도도 등과는 음의 상관성을 나타내었다. 반면에 규조류는 총질소, nitrate, 탁도, SS, TP, ASR 등과는 양의 관련성을 보인 반면 수온, PO4-P, NH3-N 등과는 음의 관련성을 잘 나타내고 있었다.
영양염의 생태적 역치농도(nutrient thresholds concentration)는 수체의 유량이나 유속, 수역의 영양염 수준 등에 따라서 다르게 나타나는데, 연구결과를 살펴보면 Chambers et al. (2012)는 캐나다의 농촌하천(agricultural streams)을 대상으로 연구한 결과 TP 농도는 0.02~0.10 mg L−1, TN 농도는 0.21~1.10 mg L−1 이라고 보고하였고, Xinquang et al. (2014)은 중국의 도시하천(urban river system)을 대상으로 연구한 결과 조류의 수화현상을 파괴하는 역치농도는 TP는 0.02 mg L−1, TN은 0.2 mg L−1 라고 했다. 낙동강에서는 역치농도 수준은 분명하지 않으나 일부지역 및 수화발생시기에 있어서의 낮은 TP 농도는 낙동강에서 수화지속시간을 감소시키는 것으로 판단이 된다(Table 5).
HC 지점에서는 낮은 TP농도에서도 수화현상이 지속되는 특성을 보여 주였다. 이 지점의 평균유속은 0.091 m s−1로서 다른 지점의 0.034~0.067보다 높았는데, Li et al. (2012)은 수체의 적당한 유속은 조류세포와 수체의 경계층 두께를 감소시키고 영양염의 확산거리를 짧게 함으로써 영양염의 대사율을 증가시키며 조류의 미소환경을 변화시킴으로써 세포의 분비물 배설에 유리하게 하여 조류성장을 도운다고 했다. 따라서 HC 지점의 다소 빠른 유속이 남조류의 영양염 대사율 증가 등의 영향으로 높은 성장을 유지하는데 일부 기여한 것으로 판단이 된다. 그러나 보다 정확한 원인파악을 위해서는 조류의 생리적 영양염 이용특성 등에 대한 추가적인 연구가 필요하다고 하겠다.
낙동강의 대형 보 건설이 조류 대발생 등 수생태계의 변화에 어떤 영향을 미치는지 연구하는 것은 중요과제로 되어 있다. 본 논문에서는 낙동강 5개 보 구간의 수질, 수리 · 수문, 조류군집에 대한 4년동안의 조사결과를 바탕으로 영양염 중심의 환경요인과 조류군집 변화와의 상관성에 대해 연구하였다. 보 설치 이후 가장 큰 물리 · 화학적 변화는 구미보 등 중 · 상류에 있어서 보 설치 이전보다 유속의 변화가 특히 심한 것과 총인의 농도가 전체 조사구간에서 큰 폭으로 감소한 것이었다. 영양염의 농도는 CG 지점 및 DS 지점에서 특히 높았고 이들 두 지점은 다른 지점과의 차이가 유의하게 나타났으며(p<0.001), 따라서 구미 및 김천지역 하 · 폐수와 금호강의 유입수가 낙동강의 영양염 변화에 가장 큰 영향인자로 판단이 된다.
규조류는 다른 분류군(class)보다 가장 높은 종수 및 현존량을 보였다. 시기별로는 3월에는
보별 방류유량 및 유입유량의 가장 큰 영향은 조류 종의 확산이었으며, 상류 보에서 특정 종의 개체수가 150 cells mL−1 이상으로 관찰될 때 하류의 보에는 92.8% 관찰되는 것으로 나타났다. 그러나 체류시간이 조류의 현존량 변화에 미치는 영향에 대한 유의성 있는 결과는 본 연구에서는 얻지 못했으며 이를 위해 보다 장기적인 연구가 필요하다고 하겠다. 조사기간 동안 TP농도는 공사중인 2011년보다 공사 후의 2012년 및 2013년에는 각각 44.1% 및 52.4% 낮아졌다. 이러한 결과를 바탕으로 낮아진 TP농도와 조류개체수와의 상관성을 확인하기 위해 RDA 분석과 Pearson 상관성 분석을 실시하였다. 분석결과 TP와 총 조류현존량과의 상관성은 지점별 및 계절별 모두 유의한 상관성이 나타나지 않았다. 다만 GG 지점에서의 4대강 사업 이후 기간(2012.4 ~ 2013.12)과 조사지점에서 7회의 수화현상 발생시기에 있어서의 남조류와 TP와의 Pearson 상관계수가 각각 0.53(p<0.0001) 및 0.42 (p<0.01)로서 통계적으로 유의성이 강하게 나타났다. GG 지점에서 TP와 상관성이 유의하게 나타난 것은 상류의 CG 지점에 다량 유입된 영양염이 체류시간이 긴 GG 지점의 조류 대발생으로 인해 영양염의 소모가 많았기 때문인 것으로 추정이 된다. 따라서 낙동강에서의 역치농도 수준은 분명하지 않으나 일부지역 및 수화발생시기에 있어서 낮은 TP 농도가 조류성장의 감소요인으로 작용하는 것으로 판단이 된다. Nitrate는 조류개체수와 음의 상관성이 유의하게 나타났으며, 조류성장에 따라 nitrate의 농도는 감소하나 조류성장을 억제하기에는 충분하지 않은 것으로 판단이 된다.
조사기간 동안 탁도와 SS 농도, ASR(적산일사량) 및 질소농도 등은 조류성장 억제 요인으로 작용하지 못했고, 다만 하절기의 수온은 조류개체수와 양의 상관성을 나타냈다.