청어목(Clupeiformes) 청어과(Clupeidae) 어류에 속하는 청어(
2015년 1월말에서 2월초 부산광역시 다대포항 인근 해역인 목도와 북형제도에서 삼중자망에 부착된 부화직전 9-10일차 수정란을 실험실로 가져와 부화시켰다. 부화장의 수온은 9℃였으며, 수정일은 어업을 목적으로 삼중자망을 설치 후 몇 시간 뒤 부착한 청어 어란으로 확인하였다. 이후, 부화직전인 자치어의 사육조건으로 수온은 자연수온과 동일하게 9-10℃로 일정하게 유지시켰으며, 사육 용수는 매일 3분의 1씩 환수하였다. 먹이는 부화 3일 후부터 클로렐라(
자치어의 성장과 형태분석을 실시하기 위해 매일 무작위로 추출하여 얼음에 마취시켜 10% 중성 포르말린에 고정하였다. 자치어의 발생단계 및 각 부위의 측정은 Okiyama (1988)의 방법을 따라 입체현미경(Olympus SZX-16, Japan) 아래에서 0.01mm 단위까지 측정하였으며, 사진촬영 하였다. 지느러미 분화를 관찰하기 위해 Alizarin red 시약으로 염색하여 계수하였다. 표본은 국립수산과학원 난자치어 분석실(Ichthyoplankton laboratory)에 보관하였다.
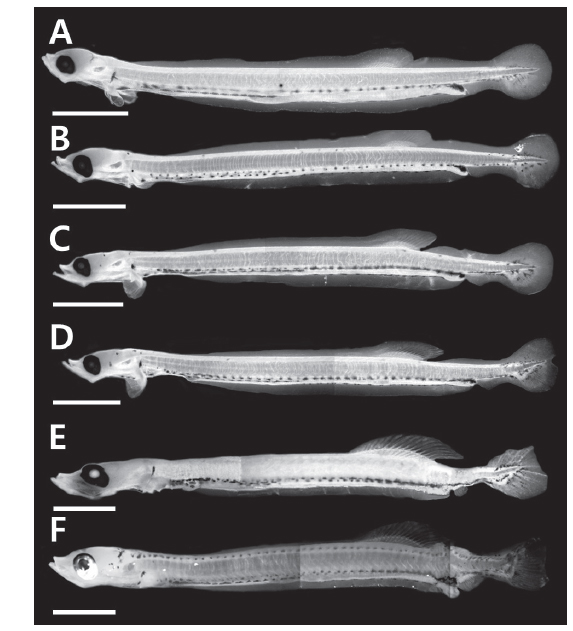
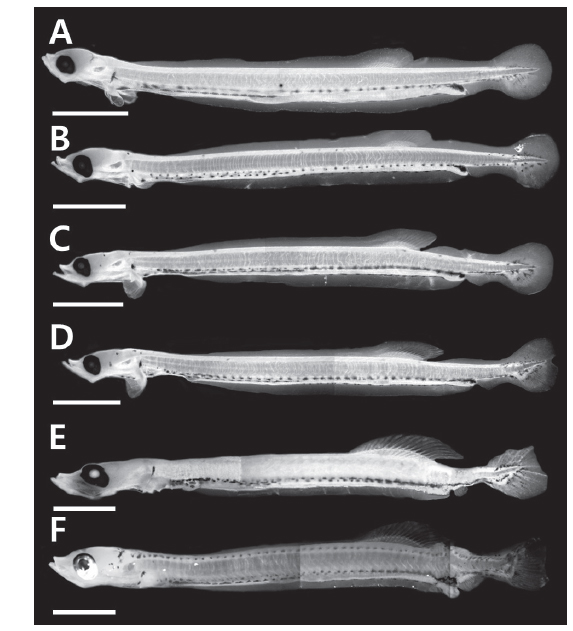
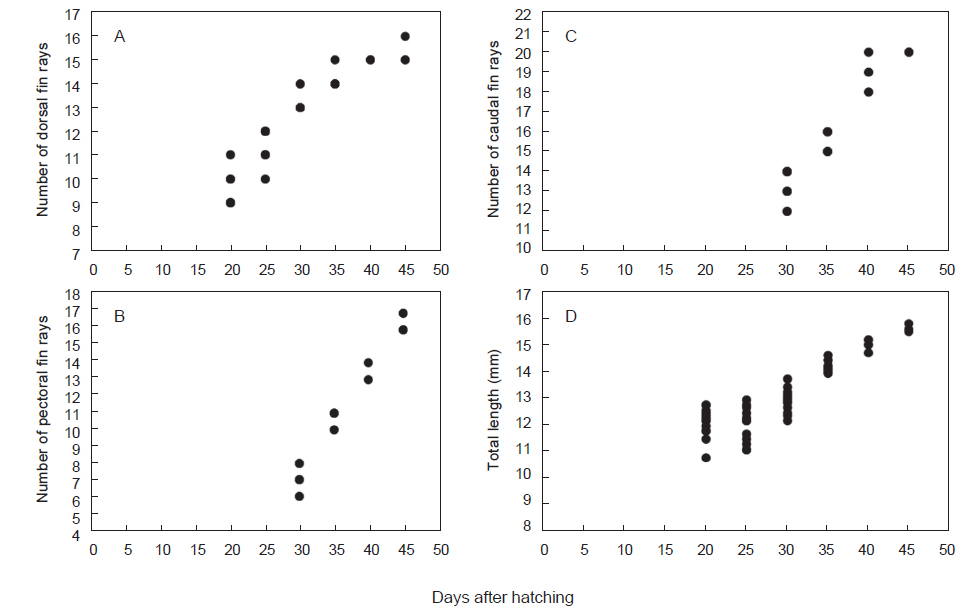
부화 20일째 자어는 전장 10.8-12.2 mm (n=13, 11.7±0.77)로 항문전장은 전장의 82.4-86.1%로 항문이 몸의 후반에 위치하며, 등지느러미앞길이는 전장의 61.8-68.5%로 항문보다 다소 전방에 위치하였다. 두장은 전장의 11.7-14.4%, 안경은 두장의 27.8-34.4%였다. 몸 전체에는 막지느러미가 관찰되며, 척색말단은 일직선이다. 가슴지느러미는 막상으로 분화하였으며, 등지느러미 담기골은 9-11개가 관찰되며 연조가 분화하기 시작하였다(Figs. 1A, 2A). 두정부에 별모양의 흑색소포가 분포하며, 머리 뒤쪽으로 나뭇가지 모양의 흑색소포가 빽빽히 소화관의 등쪽을 따라 1열로 소화관 중앙까지 나 있고, 소화관 중앙 이후부터는 등쪽과 배쪽에 항문앞까지 1열로 나 있다. 별모양 또는 나뭇가지 모양의 흑색소포는 가슴지느러미 기부와 복강의 배쪽에 각각 1개가 나있고, 항문의 등쪽과 배쪽 아래, 척색말단 주변의 등쪽과 배쪽에 짙게 분포한다(Fig. 1A).
부화 25일째 자어는 전장 11.5-12.8 mm (n=12, 12.4±0.59)로 항문전장은 전장의 76.9-84.7%, 등지느러미앞길이는 전장의 63.1-67.6%로 이전과 유사하였으며, 두장은 전장의 13.1-15.6%로 이전보다 증가하였다. 등지느러미 연조수는 10-13개로 이전보다 약간 증가하였다(Fig. 2A). 별모양의 흑색소포는 새롭게 두정부와 뺨에 분포하는 것이 특징적이다. 이전과 달리 소화관 중앙 이전의 흑색소포는 점모양 또는 별모양으로 1열로 나 있으며, 항문 주변에는 여전히 짙은 흑색소포가 분포한다(Fig. 1B)
부화 30일째 자어는 전장 12.2-13.5 mm (n=14, 13.0±0.43)로 항문전장은 전장의 78.6-84.5%, 등지느러미앞길이는 62.3-66.9%, 두장은 14.4-15.4%, 안경은 두장의 25.0-31.6%로 이전과 유사하였다. 두부의 골격이 더욱 두드러진다. 유영능력이 상당히 빠르며 섭이활동이 더욱 적극적이다. 등지느러미 연조수는 13-14개로 이전보다 증가하였으며, 마지막 연조 뒤쪽으로 막지느러미와 분리되었다(Figs. 1C, 2A). 이시기에는 가슴지느러미는 연조가 6-8개 출현하였으며, 꼬리지느러미 연조가 12-14개로 척색말단 아래부터 분화하기 시작하였다(Fig. 2B, 2C).
부화 35일째 자어는 14.0-14.7 mm (n=7, 14.3±0.24)로 항문전장은 전장의 81.6-83.6%, 등지느러미앞길이는 62.6-65.0%, 두장은 14.8-15.6%, 안경은 두장의 26.1-31.6%로 이전과 유사하였다. 등지느러미 연조수는 14-15개로 이전보다 증가하고 더욱 뚜렷하게 분화하였다(Figs. 1D, 2A). 가슴지느러미 연조수는 10-11개, 꼬리지느러미 연조수는 15-16개로 이전보다 증가하였다(Fig. 2B, 2C). 이 시기에는 척색말단이 등쪽으로 만곡되기 시작한다(Fig. 1D). 흑색소포 분포는 이전과 유사하였다.
부화 40일째 자어는 14.8-15.3 mm (n=3, 15.1±0.21)로 항문전장은 전장의 79.5-80.4%, 등지느러미앞길이는 64.1-65.5%, 두장은 14.9-16.3%, 안경은 두장의 24.0-27.3%로 이전과 유사하였다. 등지느러미 연조수는 15개로 이전과 유사하였으며, 가슴지느러미 연조수는 13-14개, 꼬리지느러미 연조수는 19-20개로 이전보다 증가하였다(Fig. 2B, 2C). 이시기에는 뒷지느러미 원기가 출현하기 시작하였으며 7-9개의 연조가 관찰되었다 (Fig. 1E). 흑색소포 분포는 이전과 유사하였다.
부화 45일째 자어는 전장 15.6-15.9 mm (n= 3, 15.7±0.12)로 두장은 전장의 16.3-17.6%로 이전에 비해 약간 커졌다. 등지느러미 연조수는 15-16개로 성어의 정수에 달하였다(Figs. 1F, 2A). 가슴지느러미 연조수는 16-17개, 꼬리지느러미 연조수는 19-20개로 이전보다 증가하였으며, 꼬리지느러미는 분화가 완료되어 연조가 뚜렷하게 관찰되었다(Fig. 2B, 2C). 뒷지느러미 연조수는 12-13개로 이전보다 증가하였다. 흑색소포는 별모양으로 두정부에 더욱 크게 발달하며, 새롭게 머리 뒤부터 몸의 등쪽 측면에 1열로 꼬리지느러미 앞까지 나 있다(Fig. 1F). 별모양의 흑색소포는 꼬리지느러미 전체에 넓게 분포한다.
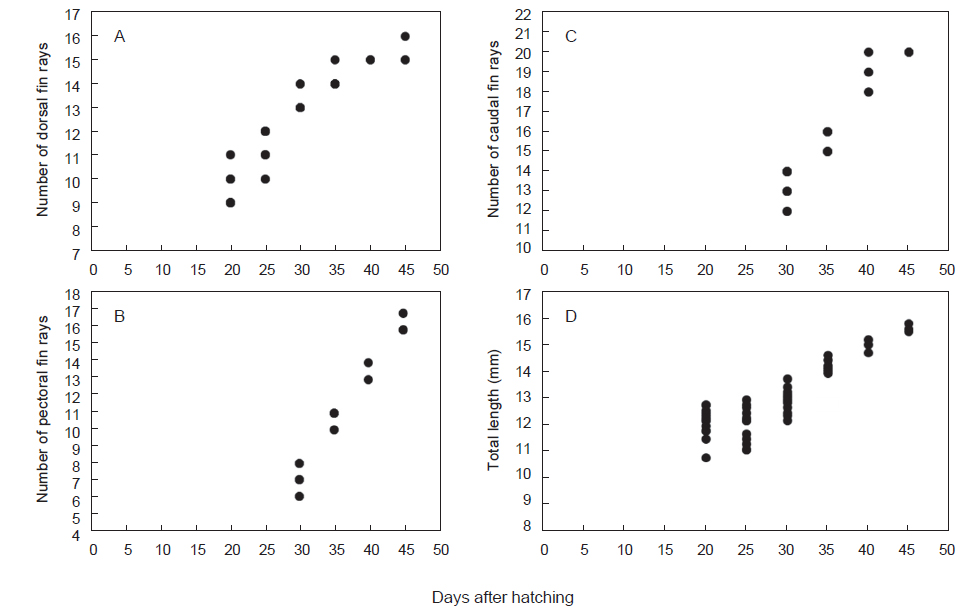
청어의 부화일수에 대한 등지느러미 발달을 살펴보면, 부화 20일차부터 부화 25일차까지는 9-12개로 발달양상이 완만하고 개체간 차이를 나타내었으나, 부화 30일 이후부터는 13-14개로 증가하였으며, 부화 35일차에는 14-15개, 45일차에는 15-16개로 성어의 정수에 달하였다(Fig. 2A). 가슴지느러미는 부화 30일차에 6-8개가 출현하였으며, 35일차에 10-11개, 40일차에 13-14개, 45일차에 16-17개로 증가하였다(Fig. 2B). 꼬리지느러미는 부화 30일차에 12-14개로 분화되었으며, 35일차에는 15-16개, 부화 40-45일차에 19-20개로 분화가 완료되었다(Fig. 2C).
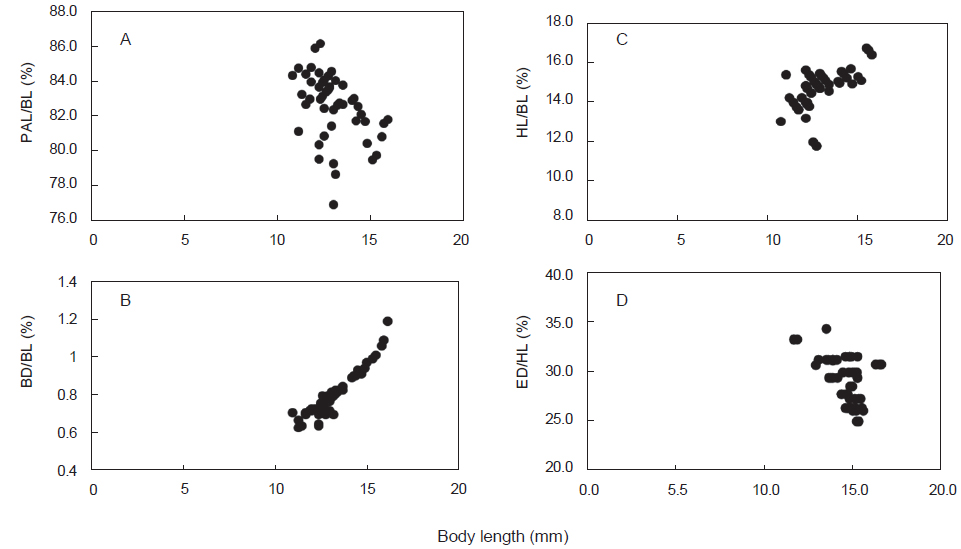
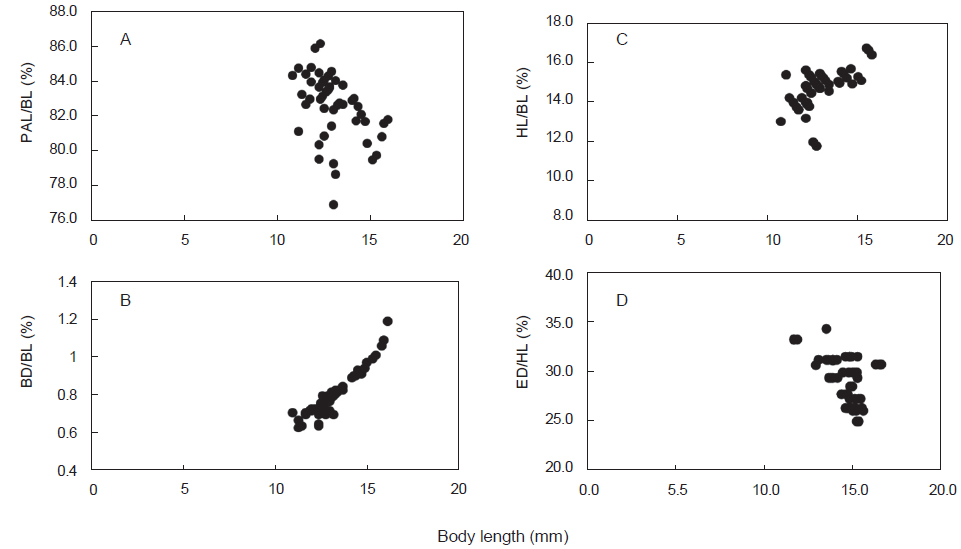
청어의 부화일수에 대한 상대성장은, 부화 후 20일부터 25일까지는 성장율이 완만하고, 개체간 전장 차이가 컸으며, 30일 이후부터는 성장율이 빨라 지는 경향을 나타내었다(Fig. 2D). 체장(BL)에 대한 백분비로 두장과 체고는 성장하면서 점차 증가하는 경향을 나타내었으며, 항문장과 두장에 대한 안경은 개체간 변이만 관찰되었다(Fig. 3).
본 연구에서는 부산 다대포 인근 연안에서 삼중자망에 부착된 부화직전 청어 어란을 실험실로 운반하여 부화 후 20일부터 45일까지 자어의 형태발달을 관찰하였다. 다대포산 청어는 전장 14.0 mm 이상부터 척색말단이 만곡되기 시작하였는데, Han et al. (2011)에 의하면 전장 15.8 mm에서 만곡되어 약간의 차이를 나타내었다. 이는 사육환경에 따른 사육수온(본 연구에서는 9-10℃ vs. 동해산 청어 13-14℃)과 먹이(본 연구에서는 14일차부터 알테미아 급이 vs. 20일까지 로티퍼만 급이)에 따른 차이인 것으로 판단된다(Han et al., 2011). 유사종과의 척색말단 만곡시점을 비교하면 전어(
다대포산 청어의 발육단계별 등지느러미 형성은 전장 10.8-12.2 mm에서 9-11개, 전장 12.2-13.5 mm에서 13-14개로 증가하였으며 전장 15.6-15.9 mm에서 성어 청어의 등지느러미수(15-19개)와 일치하여 정수에 달하였다(Fig. 2A). 한편, 남해산 청어의 등지느러미 출현시기는 전장 15.8 mm로 5-6개의 연조가 분화하였는데, 본 연구와 전장에서는 유사하였으나 지느러미 발달에서는 많은 차이를 나타내었다(Han et al., 2011). 이는 사육수온과 사육일수에 따른 차이일 것으로 판단된다. 발육단계별 가슴지느러미 형성은 전장 12.2-13.5 mm에서 6-8개로 출현하였으며, 전장 14.0-14.7 mm에서 10-11개, 전장 14.8-15.3 mm에서 13-14개, 전장 15.6-15.9 mm에서 16-17개로 증가하였다(Fig. 2B). 본 연구의 45일차 청어 자어(전장 15.6-15.9mm)의 가슴지느러미 연조는 약하게 분화하였으며, 일본산 청어의 경우 전장 20.0 mm자어에서 뚜렷하게 형성되어 치어기에 이르러 분화가 완료되는 것으로 판단된다(Uchida, 1958). 발육단계별 꼬리지느러미 형성은 전장 12.2-13.5 mm에서 12-14개, 전장 14.0-14.7 mm에서 15-16개, 전장 15.6-15.9 mm에서 20개로 연조가 뚜렷하게 분화완료 되었다(Fig. 2C). 일본산 청어는 전장 16.8 mm에서 꼬리지느러미 연조수가 20개로 본 연구와 유사하였다(Uchida, 1958).
본 연구의 부화 40일차 청어 자어(전장 14.8-15.3 mm)에서 처음 뒷지느러미 원기가 출현하였는데, 일본산 청어(전장 16.8 mm)의 뒷지느러미 출현시기와 서로 유사하였으며, 유사종인 전어(전장 13.9 mm), 정어리(전장 13.3 mm), 눈퉁멸(전장 11.6 mm) 보다 약간 느린 것으로 나타났다(Fig. 1E; Uchida et al., 1958). 다대포산 청어 자치어의 지느러미 발달은 등지느러미, 꼬리지느러미, 가슴지느러미, 뒷지느러미 순이었으며, 배지느러미의 경우 45일차 청어 자어에서는 관찰되지 않았으나, 일본산 청어 전장 25.3 mm 에서 발달하여 가장 늦게 생성되는 것으로 나타났다(Uchida, 1958).
한편, 부화 45일차 청어 자어(전장 15.6 mm)에서 몸의 등쪽 나뭇가지 모양의 흑색소포가 1열로 꼬리지느러미 앞까지 새롭게 분포하기 시작하였는데, 일본산 청어(전장 16.8 mm, 25.3 mm)의 형태 스케치에서는 확인되지 않았다(Uchida et al., 1958; Okiyama, 1988). 한편, 남해산 전어를 인공사육한 연구에서 전장 11.6 mm까지는 몸의 등쪽에 흑색소포가 없다가 전장 16.8 mm 부터 새롭게 1열로 발달하는 것으로 보고되어있다(Kim et al., 2007). 따라서, 청어 또한 후기자어로 성장하면서 체측 등쪽의 흑색소포가 발달하는 것으로 사료된다.
다대포산 청어 자어의 발육단계별 계측형질 변화를 살펴보면, 전장에 대한 두장, 체고는 양의 상대 성장을 나타내었으나 전장에 대한 항문장, 두장에 대한 안경은 성장에 따른 차이 없이 개체간 차이를 나타내었다(Fig. 3). 본 연구는 부산 다대포 인근 해역이 청어의 산란장 중 하나인 것으로 확인되었으며, 1월말에서 2월초에 집중 산란하는 것으로 보인다.