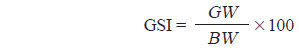Two
대게류(
대게는 한국 동해안에서 맛이 뛰어난 고급 수산물로서 상품가치가 높고 겨울에는 동해안 관광자원으로도 중요한 어종이다. 한국연안에서는 일반적으로 자망이나 저인망, 그리고 통발어업에 의해 주로 어획되고 있으며 현재 연간 1,000톤 전후의 어획량을 보이고 있다. 한국은 대게 자원보호를 위해 총허용어획량(TAC) 제도를 시행중에 있으며, 모든 크기의 암컷과 두흉갑장 9 cm이하의 수컷은 채포를 금지하고 있다. 또한 6월부터 10월까지는 금어기를 설정하여 자원을 관리 보호하고 있다. 대게에 관한 국내 연구로는 성숙과 산란(Chun et al., 2008), 탈피와 성장(Chun et al., 2009)에 관한 연구만이 있으며, 국외에서는 생식소 구조(Kon and Honma, 1970; Chiba and Honma, 1971), 난소내 포란수(Kon, 1974; Kanno, 1987), 난의 숙도(Ito, 1963; Brown and Powell, 1972; Comeau and Conan, 1992), 교미 및 산란(Watson, 1970), 성숙대게 분포(Elner and Beninger, 1992; Ennis et al., 1988) 및 먹이생물(Hubert and Earl, 2003) 등 많은 분야에서 연구가 수행되고 있다.
붉은대게(
본 연구는 한국 동해안에 분포하는 대게류의 재생산 및 분포 특성에 대한 생태를 밝히고자 대게와 붉은대게의 생식소숙도 지수, 외포란단계 변화, 포란수, 군성숙체장, 탈피단계를 조사하고, 대게류에 대한 수심별 분포특성을 구명하여 최근 감소되는 대게류 자원의 회복 및 관리방안을 마련하는데 목적을 두고 있다.
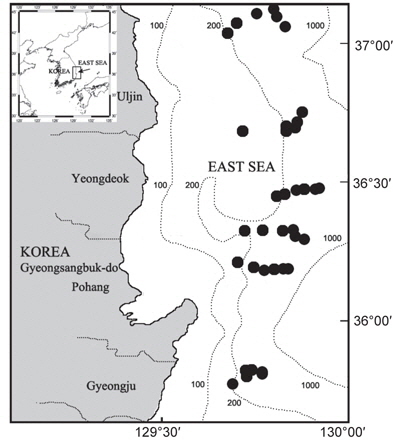
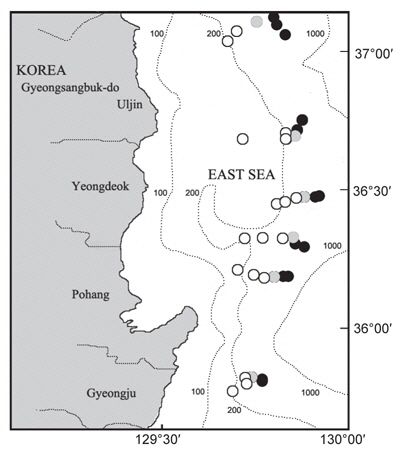
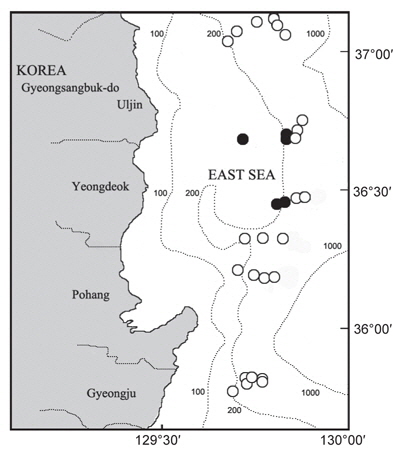
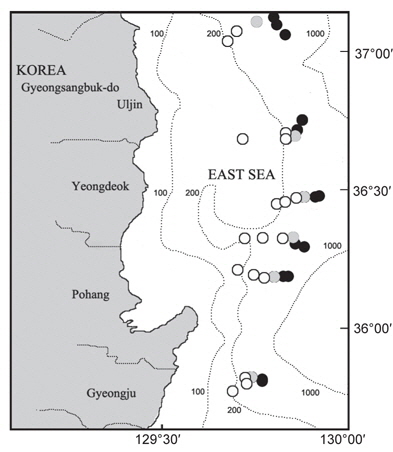
한국 동해에 서식하는 대게류 자원의 산란생태 및 분포특성을 파악하기 위해 경상북도 울진군에서 경주시까지 6개의 정선을 선정한 후, 각 정선에 수심별 200, 300, 400, 450, 500, 600 m로 6개의 정점을 나누어 총 36개 정점에서 2007년 9월부터 2008년 8월까지 매월 어획 및 생태조사를 실시하였다(Fig. 1).
본 조사에 사용한 어구는 망목 125 mm의 붉은대게 통발로, 각 정점별로 통발10개를 투망하였고, 양망은 투망 후 5일 이내에 실시하였다.
어획조사에 의해 채집된 대게류에 대해 어종별로 분류학적 형질을 기준으로 암컷과 수컷으로 분류한 후 생태학적 정밀조사를 수행하였고, 두흉갑장(carapace length)과 두흉갑폭(carapace width)은 0.1 mm 단위까지, 체중(total weight)과 생식선 중량(gonad weight), 외포란중량(egg weight)은 0.01 g까지 측정하였다.
대게류의 생식소 발달상태를 알기 위하여 생식소 숙도지수(Gonadosomatic index, GSI)를 다음식으로 구하여 월별 변화를 추정하였다.
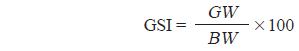
여기서 GW는 생식소 중량(g, 습중량), BW는 체중(g, 습중량)을 각각 나타낸다.
대게류 암컷 성숙의 육안판별을 위해 외포란 단계를 파악하였는데, 외포란 단계는 Ito (1963)와 Hilsinger (1976)의 방법을 참고하여 난의 색과 광학현미경하에서의 난 내 발생상태를 기준으로 다음과 같이 4단계로 구분 하였다.
Stage Ⅰ: 연한 오렌지색을 띠며, 현미경상에서는 난 내에 작은 과립상 난황으로 채워져 있으나 눈은 생기지 않음.Stage Ⅱ: 오렌지색을 띠며, 현미경상에서는 난 내에 3/4 가량의 난황이 채워짐.Stage Ⅲ: 다갈색을 띠고, 현미경상에서는 난 내에 삼각형의 작은 눈이 생김Stage Ⅳ: 난은 거의 흑갈색에 가까우며, 현미경상의 난 내에는 눈이 삼각형으로 크고 검음.
대게류의 외포란수는 성숙한 암컷개체의 복절에 부착된 외포란을 미세한 침에 의해 완전히 분리하여 해부현미경으로 계수한 후, 습중량법(Bagenal and Brown, 1978)으로 구하였다.
생물학적 성숙체장을 구하기 위해 성숙개체의 판별은 Watson(1970)의 방법을 참고하였고, 복절이 작아 복부 전체를 덮지 못하고 생식선 내의 난소색이 유백색 또는 연한 오렌지색을 나타내는 개체는 미숙, 복절에 외포란을 가졌거나 복부 전체를 덮고 생식선 내의 난소색이 짙은 오렌지색을 나타내는 개체를 완숙으로 판정하여, 체장계급을 1 mm 단위로 구분하여 성숙개체의 비율을 구한 후 logistic 식(Zhang, 1991)에 적용시켜 50% 군성 숙체장을 구하였다.
대게류의 탈피단계는 손가락으로 갑각을 누르면 함입되지 않고 딱딱한 것은 경갑(hard-shell)이고, 손가락으로 갑각을 눌러 바로 합입되는 것은 연갑(soft-shell)으로 정하여 월별 비율을 분석하였다.
대게류의 분포특성을 파악하기 위하여 수심별로 대게 및 붉은대게의 종조성을 조사하였고, 대게류 정점별 분포를 통해 해역별 수심별 분포를 파악하였다. 그리고 대게류의 산란장을 추정하기 위해 암컷 대게류의 해역별 수심별 분포를 조사하였다.
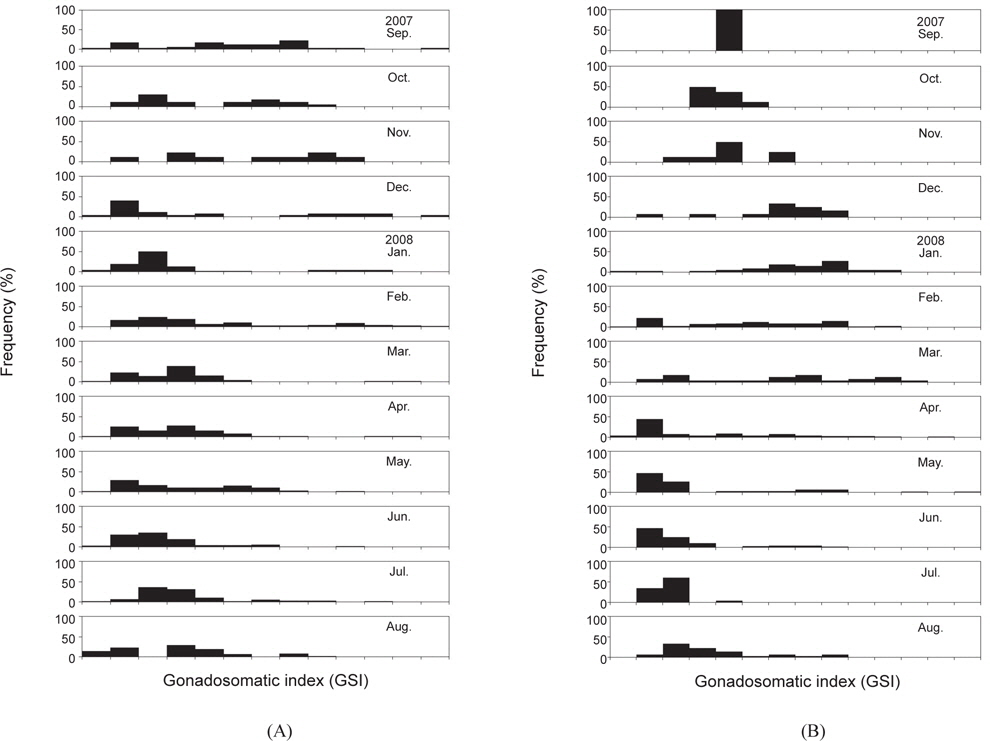
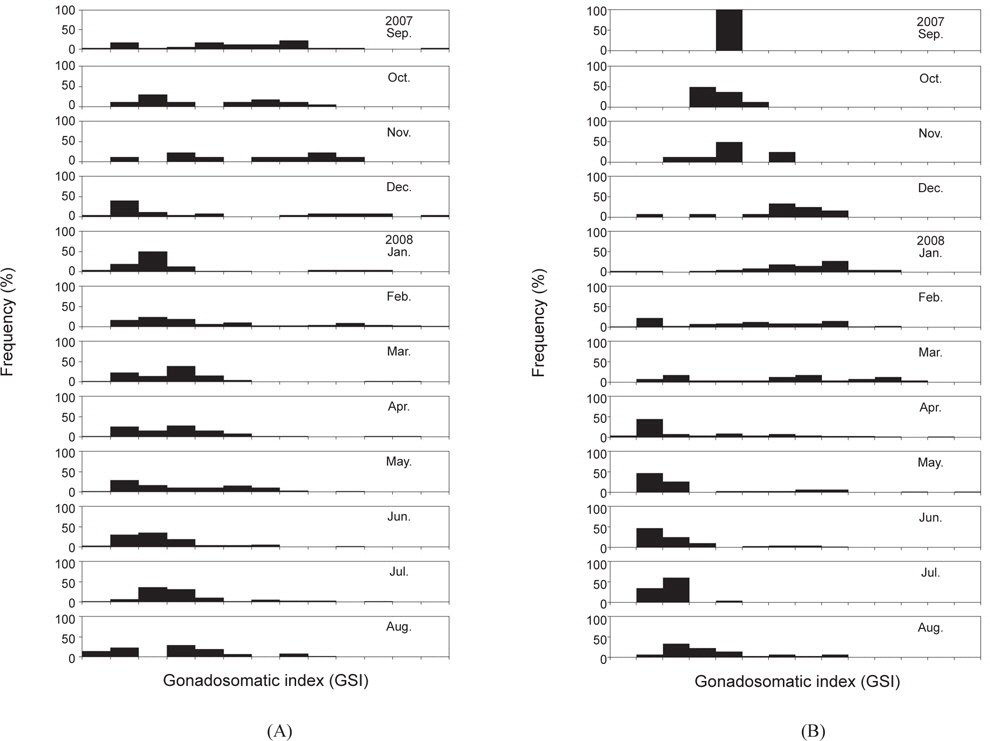
대게 암컷에 대한 생식소숙도지수(GSI)는 2007년 9월부터 2008년 4월까지 GSI 값이 0-12까지의 광범위한 값을 나타내었으며, GSI 값이 6 이하인 개체들은 대부분이 난소가 유백색이거나 연한 오렌지색으로 미숙상태였다. GSI 값이 7 이상인 개체들의 난소색은 미숙 상태보다 짙은 오렌지색을 나타내었고, 난소가 복강의 대부분을 차지하고 있었으며, 외포란색은 갈색 또는 흑색을 띄었다. 3월 이후 GSI 값의 주 모드가 크게 감소하여 대부분이 6 이하의 낮은 값을 나타내었으며, 3월-4월에 GSI값이 10-11인 개체가 일부 출현하였다[Fig. 2(A)].
붉은대게 암컷의 GSI는 2007년 9월부터 11월에 GSI 값이 2-6으로 범위가 가장 좁았고, 2007년 12월-2008년 8월에는 GSI 값이 0-13으로 대체로 넓은 분포 범위를 보였다. 2008년 5월에 GSI 값이 최고치를 나타내었고, 2008년 4월과 5월에 난소의 색이 유백색이거나 연한 오렌지색인 개체들이 많이 출현하였으며, 그 외에는 대부분이 짙은 오렌지색을 나타내었다[Fig. 2(B)].
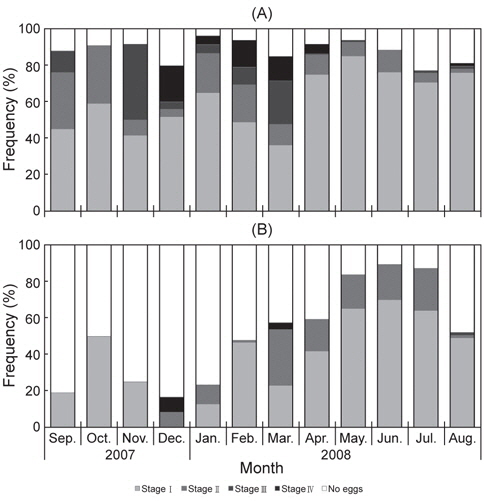
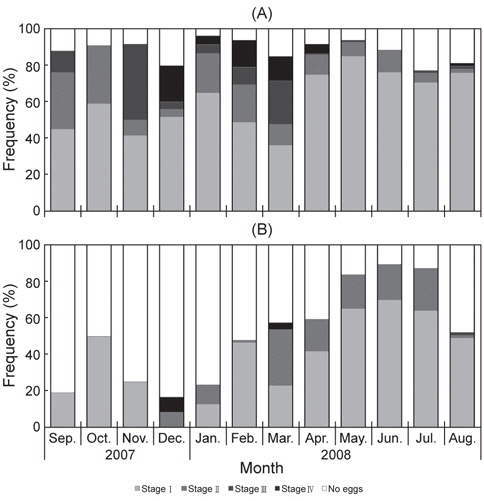
대게 암컷에 대한 외포란 단계를 보면 Stage Ⅰ과 Ⅱ의 개체들이 2007년 9-10월까지 대부분을 차지하였고, Stage Ⅲ의 개체는 2007년 9월에 약 12%로 출현 하여 11월에 42%로 가장 높았으며, 2008년 3월까지 계속 높은 출현율을 보였다. Stage Ⅲ의 개체는 2008년 4월 이후에는 거의 출현하지 않았고, 7-8월에 일부 개체가 출현하였다. Stage Ⅳ의 개체들은 2007년 12월부터 출현하기 시작하여 2008년 4월까지 출현하였으며, 매월 출현율은 20%, 5%, 15%, 13% 그리고 5%로 월별 출현율이 다르게 나타났다[Fig. 3(A)].
붉은대게 암컷에 대한 외포란 단계를 보면 StageⅠ의 개체들은 2007년 12월을 제외하고 전 조사기간 중 13-65%로 출현하였고, Stage Ⅱ의 개체는 2007년 12월부터 2008년 8월까지 1-31%로 출현하였다. Stage Ⅲ의 개체들은 2008년 8월에 출현하였고, Stage Ⅳ의 개체들은 2007년 12월과 2008년 3월에 출현하여 월별 뚜렷한 경향을 보이지 않았다[Fig. 3(B)].
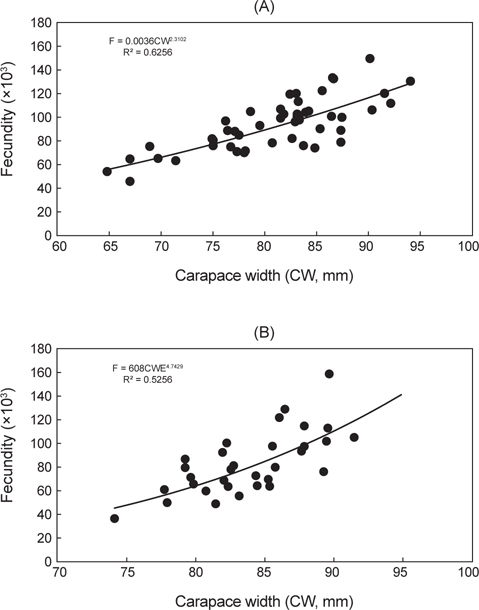
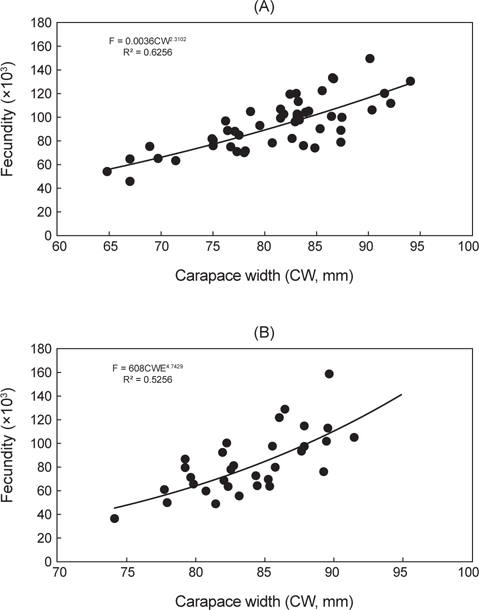
대게의 재생산력을 알아보기 위해 외포란을 하고 있는 암컷 개체 49마리를 대상으로 포란수를 계수한 결과, 포란수는 최소 44,791개(갑폭 66.9 mm), 최다 151,538개(갑폭 90.0 mm)로 계수되었다. 갑폭에 따른 절대포란수는 체장이 커질수록 증가하는 것으로 나타났다[Fig. 4(A)].
붉은대게 45마리를 대상으로 포란수를 계수한 결과, 최소 35,012개(갑폭 74.1 mm), 최다 159,593개(갑폭 89.6 mm)로 계수되었다. 갑폭에 따른 포란수는 체장이 커질수록 증가하는 것으로 나타났다[Fig. 4(B)].
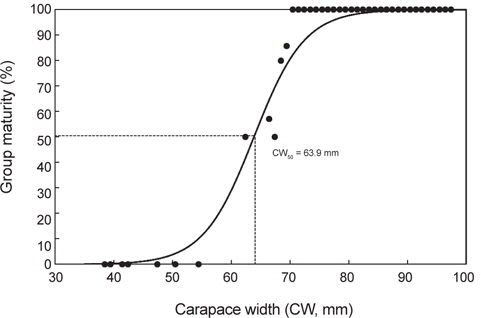
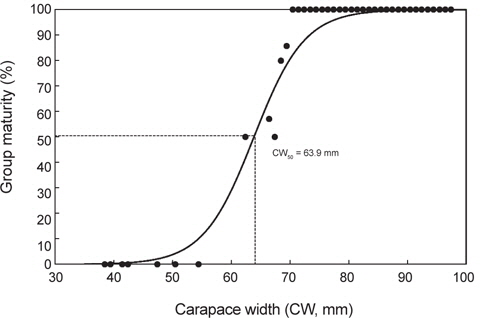
대게 암컷이 재생산에 참여하는 생물학적 성숙체장을 알아보기 위해 산란기 전후로 추정되는 1-5월의 암컷 260마리를 대상으로 체급별 성숙개체의 출현율을 조사하였다. 체급별 성숙개체는 갑폭 54 mm 이하에서는 성숙개체가 출현하지 않았고, 62 mm에서 50%, 66 mm에서 57%, 68 mm에서 80%, 69 mm에서 86%, 그리고 70 mm 이상에서는 모든 개체가 성숙개체로 판명되었다. 따라서 대게 암컷의 50% 군성숙체장은 이들 체급별 성숙개체의 출현비율을 logistic 식에 적용하여 추정한 결과, 갑폭 63.9 mm로 산출되었다(Fig. 5).
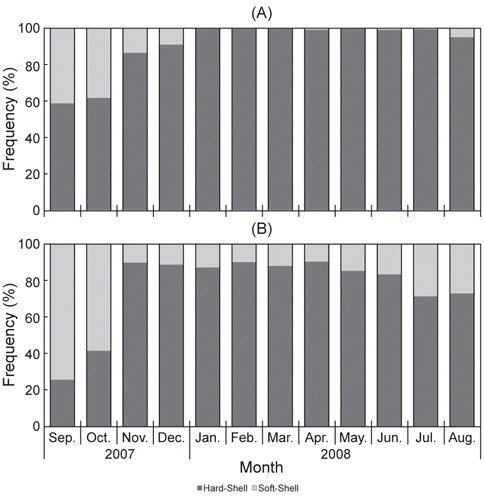
대게 수컷의 경갑상태 개체의 출현율은 2007년 9월에 49%로 가장 낮았고, 10월에 60%, 11월 86%, 12월에 90%로 나타났으며, 2008년 1-3월까지는 출현한 모든 개체가 경갑상태였다. 4월 이후부터 다시 연갑상태가 일부 개체에서 출현하기 시작하였고, 8월에 경갑상태 개체의 출현율은 91%였다[Fig. 6(A)].
붉은대게 수컷 경갑상태 개체의 출현율은 2007년 9월에 26%로 가장 낮았고, 10월에 41%이었다. 11월부터 경갑상태 개체가 크게 증가하여 2008년 6월까지 80% 이상의 출현율을 보였으며, 7월 이후 다소 감소하여 69%, 8월에 72%를 나타내었다[Fig. 6(B)].
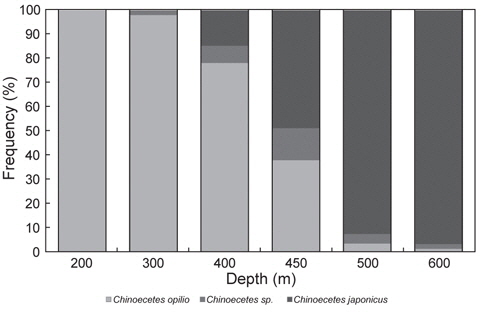
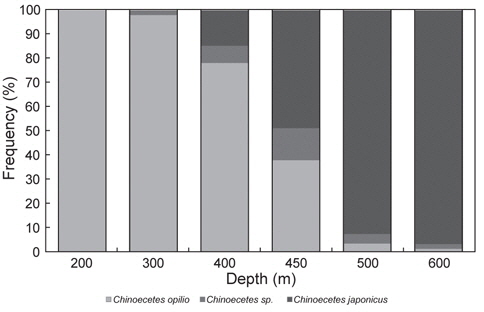
대게류의 수심별 종조성은 수심 200-300 m에서 대게만이 출현하였고, 400 m에서는 대게가 85%, 붉은대게가 15%로 대게의 출현율이 높았다(Fig. 7). 450 m에서는 대게가 51%, 붉은대게가 49%로 출현율이 비슷하였으며, 500-600 m에서는 대게가 3-8%, 붉은대게가 92-97%로 수심별로 어종별 출현율의 차이가 명확하였다(Fig. 7).
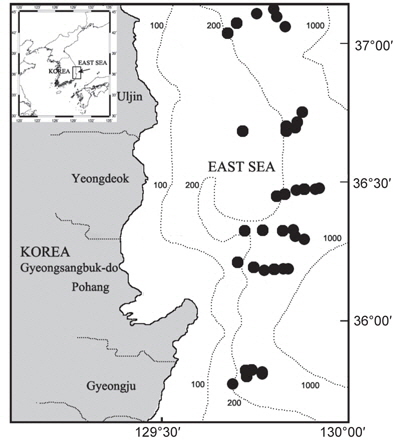
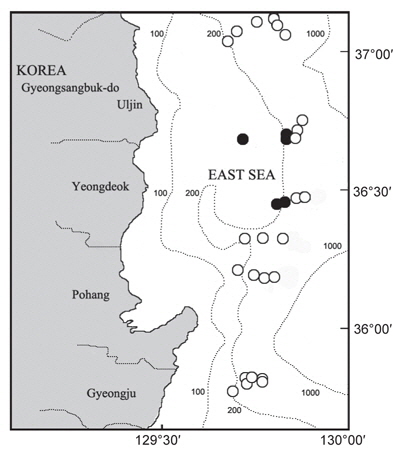
대게류의 정점별 분포를 살펴보면(Fig. 8), 수심 200-300 m에서는 대게만이 출현하였고, 대게 암컷 및 두흉갑폭 9 cm 이하의 소형 수컷 개체들이 많이 어획되어 조사수심이 대게의 주요 산란장 및 성육장의 역할을 하고 있는 것으로 추정된다. 수심 400 m에서는 울진정점에서 한 시기에 붉은대게가 우점하여 출현한 정점이 있었으나, 대체로 대게의 출현율이 높았다. 수심 450 m에서는 지역에 따라 대게의 분포 밀도가 높거나 혹은 붉은대게의 분포 밀도가 높았으며, 동일한 조사정점이더라도 시기에 따라 대게와 붉은대게의 분포 밀도 및 출현 양상이 서로 상반되게 나타났다. 수심 500-600 m에서는 대게가 일부 출현한 정점이 있었으나 출현한 개체의 대부분이 붉은대게로 500 m 이심에서는 대게의 분포 밀도가 극히 감소하였다.
대게 암컷의 출현은 주로 수심 200-300 m에서 출현하였는데, 울진 후포해역 수심 200-300 m와 영덕 축산해역 수심 200-300 m를 중심으로 높은 분포밀도를 나타내어 이 해역이 대게의 주요 산란장인 것으로 추정되었다(Fig. 9).
대게류는 복절 형태를 통해 성별 구분이 가능하며, 암컷의 성숙상태를 알 수 있다. 대게 수컷의 복절은 삼각형에 가까운 형태를 보이며, 암컷의 복절은 둥근 원형의 형태를 하고 있고, 성숙된 암컷의 복절 안쪽에는 포란된 난이 많이 부착되어 있다(Chun et al., 2008).
대게 암컷에 대한 생식선 숙도지수(GSI) 및 외포란 발달단계별 월 변화로부터, 대게 암컷의 난소가 7-8월경부터 성숙하기 시작하고 3-4월경에 외포란된 난이 부화하여 완전히 탈락하게 되는 것으로 나타나 대게의 주 산란기는 3월-4월인 것으로 추정된다. 우리나라 인근해역인 일본해역의 대게 연구결과를 살펴보면, Ito (1963)는 동해의 일본측 해역에 서식하는 대게의 주 산란기도 3-4월경인 것으로 보고하였고, Kon and Honma(1970)는 일본산 대게 암컷의 경우 7-9월경에 성숙하여 탈피한 후 첫 번째 산란을 하며, 2회째 산란은 겨울-봄철에 걸쳐 이루어지는 것으로 추정하였다. 대게는 연구자에 따라 초산란군과 경산란군으로 분류하고 있으며, 생애 처음 산란에 가입하는 초산란 시기가 8-9월(Ito, 1963), 8-11월(Yamasaki et al., 1985) 등 지역에 따라 다소 차이가 있어서, 산란횟수, batch fecundity 등 지속적인 연구가 필요한 것으로 생각된다.
붉은대게의 산란기는 GSI 값의 월 변화가 뚜렷한 경향을 보이지 않고, 외포란된 난이 탈락한 성숙크기 이상의 개체들이 매월 출현하는 것으로 보아 연중 산란할 가능성이 있는 것으로 추정된다. 붉은대게는 연중 동해의 심해 저층수온 2℃ 이하의 냉수역에 서식하므로 성숙과 산란에 있어 일반 수산생물처럼 단기간에 일시에 이루어지는 것이 아니고 장기간에 걸쳐서 서서히 이루어지는 것으로 추정된다. 이는 본 조사 시 매 계절마다 상당한 수의 개체가 외포란을 하고 있었으며, 방후의 개체도 연중 확인됨에 따른 것이다. 일본의 붉은대게 연구에서는 산란개체의 출현율을 토대로 매 2년마다 3-4월에 산란할 가능성이 있다는 보고(Yosho, 2000)를 하였으나 정확한 결과로는 해석하기 어려워 추후 연구가 필요한 실정이다.
본 연구에서는 대게의 포란수가 44,791-151,538개로 계수되었고, 일반 수산생물과 동일하게 크기가 클수록 포란수가 증가하는 경향을 보였으며, 암컷의 생물학적 성숙체장은 63.9 mm (CW)로 추정되었다. 한편, 알라스카산 대게의 포란수는 24,000-318,000개(Hilsinger, 1976), 캐나다 동부산 대게는 20,000-140,000개(Watsaon, 1969), 동해 일본측 대게는 6,000-130,000개(Ito, 1963)로 알려져 있으며, 본 조사결과와 큰 차이는 없는 것으로 나타났다.
대게류의 탈피단계에 따른 월 변화 결과를 보면, 대게와 붉은대게 모두 경갑상태의 개체가 11월-이듬해 8월에 높은 출현율을 보였고, 탈피 직전인 이중갑상태의 개체는 6-8월 사이에 출현하였으며, 연갑상태의 개체는 9-10월에 높은 출현율을 보여 대게 및 붉은대게의 주 탈피시기는 9-10월인 것으로 추정된다. 이는 좀 더 지속적이고 정밀한 연구방법을 통하여 추후 정확한 연구결과를 도출할 계획이다.
대게류 수심별 분포는 400 m 이천에서 대게의 출현율이 높았고, 수심 500 m 이심에서는 붉은대게의 출현율이 높게 나타났다. 수심 450 m에서는 조사정점 및 시기에 따라 대게와 붉은대게의 출현율이 상반되게 나타나 수심 400-450 m가 두 어종간의 경계 수심인 것으로 추정되었다. 또한 500 m 이천 수역에서 어획된 대게 중 너도대게라 불리는 몇몇 개체가 출현하였다. 동해의 일본측 해역에서 서식하는 대게와 붉은대게의 분포수심은 최대 520 m로 조사되었는데(Yosho, 1994), 이 조사결과는 한정된 지역과 횟수로 본 조사와 비교 설명하기에 무리가 있을 것으로 생각되지만 우리나라와는 다소 차이가 있으며, 이는 서식 해역의 환경차이에 기인하는 것으로 생각된다. 또한 미국 베링해에서의 대게는 수심 200 m 이내의 대륙붕에 하나의 개체군으로 서식한다고 알려져 있어(NOAA, 2014), 우리나라와는 다른 서식형태를 보이고 있다.
대게류의 산란장은 대게 암컷에 있어 울진 후포해역과 영덕 축산해역의 수심 200-300 m를 중심으로 높은 분포밀도를 나타내어 이 해역이 대게의 주요 산란장인 것으로 추정되었다. 붉은대게 암컷은 수심 400-600 m에서 출현하였으며, 수심 600 m에서 높은 분포밀도를 보였다. 그러나 본 연구에서의 최대 조사 수심대가 600 m이고, 붉은대게는 수심 2,300 m 까지 분포하는 것으로 알려져 있어(Hong et al., 2006), 붉은대게의 주 산란시기 및 산란장을 알기 위해서는 향후 조사가 보완되어야 할 것으로 생각된다.
또한 본 연구에서 대게류의 재생산과 분포특성 연구를 통해 대게류의 자원관리에 필요한 기초자료를 제시할 수 있으나, 향후 대게류 자원의 풍흉에 따른 자원생물학적 변수의 변화가 예상되므로 적절한 대게류 자원관리를 위해 해양환경 변화와 더불어 종합적 연구가 지속되어야 할 것으로 생각된다.