Marine macroalgal community structures and characteristics of ocean environmental factors were examined seasonally at four sites in Jeju Island, Korea, from March to November 2012. A total of 71 macroalgal species were identified, including 9 green, 7 brown, and 55 red algae.
조하대 해조류 군집은 다양한 어·패류와 무척추동물의 산란, 생육 및 먹이장으로서 연안생태계의 생물다양성과 생산성에서 중요한 기능을 가진다(Terawaki et al., 2001). 전 세계 해조류의 경제적 가치는 약 110억 달러 정도이며, 특히 켈프숲은 육상의 경작지에 비해 약 4배 이상의 생산성을 보인다(Millar, 2011). 이외에도 해조류는 전통적으로 인간에게 식용과 약용으로 활용되었으며, 최근에는 이차산물인 아가, 카라기난, 알긴산 및 다른 화합물들이 건강식품과 식품산업에 다양한 형태로 이용되고 있으며, 신약과 화장품의 원료로도 관심을 받고 있다(Oh et al., 1990; Cha et al., 2006). 또한, 해조류는 해수에서 영양염을 직접 흡수하기 때문에 생물정화자(bio-filter)로서, 그리고 인간의 직접적인 활동(예, 부영양화, 양식, 퇴적물, 해양건설, 먹이망 간섭 등)과 간접적인 활동(예, 기후 변화, 질병 발생)에 의해 군집구조가 변화되거나 소멸되므로 생물지시자(bio-indicator)로 사용되고 있다(Díez et al., 1999; Orfanidis et al., 2001; Arévalo et al., 2007; Wells et al., 2007; Song et al., 2011). 최근 전세계인의 관심 대상인 기후변화로 지구 대기 온도는 지난 세기 동안에 약 0.74℃ 상승하였으며(IPCC, 2007), 우리나라 연근해의 표층 수온은 지난 41년 동안에 1.30℃ 상승하였다(Seong et al., 2010). 특히, 우리나라 남쪽에 위치한 제주도 연안의 표층 수온은 최근 86년 동안에 1.94℃(연안정지 관측자료 분석 결과) 상승하여 전 세계 평균수온의 상승 정도인 0.67℃보다 훨씬 높게 나타남으로써(Belkin, 2009), 제주도가 아열대 해역으로 급변하는 것으로 보고되었다(Suh et al., 2011).
제주 연안에서 해조류 연구는 1960년에 153종을 최초로 보고하였으며(Kang, 1960), 이후 암반 조간대의 해조상과 군집 구조에 대한 연구(Lee, 1974; Lee and Lee, 1976; Lee, 1976; Lee and Lee, 1982; Yoon, 1985; Boo, 1988; Kim, 1991; Kim et al., 2008)와 천이 양상(Kim and Park, 1997) 및 정성과 정량적 분석방법(Park et al., 1994)에 대한 연구가 수행되었다. 이러한 연구는 대부분 조간대 해조군집에 한정되었으며, 제주도 조하대에 관한 연구는 문섬과 우도에 서식하는 해조류의 계절별 생물량 변화(Ko et al., 2008; Kang et al., 2011)와 갯녹음의 현황파악을 위한 서귀포 연안에서 수행된 연구가 있다(Chung et al., 1998). 조하대 지역은 조간대에 비해 상대적으로 환경이 안정되고, 넓은 면적을 차지하며, 해조류의 종다양성과 생물량이 풍부한 특징을 보이고 있어 조하대 해조군집에 대한 연구자료 는 향후 환경과 연안생태계 변화를 확인할 수 있는 중요한 척도로 사용될 수 있다(Yoo, 2003; Choi et al., 2006). 하지만, 조하대 생태계의 일차생산자인 해조류의 종다양성과 계절별 생물량 변화를 파악하는 것은 SCUBA diving을 통해서만 연구가 가능한 제한성으로 인해 많은 비용과 노동력이 요구되므로 연안생태계를 이해하고 관리 및 보존할 수 있는 과학적인 기초 자료가 부족한 실정이다. 특히, 제주도는 쿠로시오 난류의 지류인 대마난류와 기후변화로 인해 수온이 상승한 중국 양자강의 영향을 받고 있어 조하대 해조류 군집구조를 파악하는 것은 향후 기후 변화에 따른 우리나라 연안의 생태계 변화를 예측할 수 있는 중요한 자료가 될 것이다(Belkin, 2009). 제주도는 여름철에 고온과 저염의 중국대륙 연안수의 영향을 받으며, 계절에 따라 남해연안수와 황해저층냉수 등의 영향과 함께 해안선 인근의 용천수와 여름철 육지에서의 담수유입으로 인해 해양환경의 변화가 매우 복잡한 특성을 보인다(Kim and Rho, 1994). 또한, 제주도 연안에서는 온대해역인 대한민국의 본토와는 달리 열대와 아열대성 생물들이 다양하게 서식하며, 해조류의 종다양성도 우리나라의 다른 해역에 비해 높은 것으로 나타났다(Park et al., 1994; Yang et al., 2007).
해조류의 수평분포는 그들의 생장과 성숙을 제한하는 수온에 의해, 그리고 수직분포는 환경스트레스인 건조 내성(조간대)과 조도(조하대)에 의해 결정된다(Lüning, 1990; Breeman and Pakker, 1994). 제주도 문섬의 조하대 해조류 생물량(kg wet wt/m2)은 수심 5-10 m에서 감태(1-3 kg), 유절석회조류(0.1-1.5 kg), 모자반 속과 미역 등의 해조류가 번무하였으며, 연평균 생물량은 수심 1 m에 비해5-10 m에서 높게(약 3 kg wet wt/m2) 나타났다(Ko et al., 2008). 갯녹음이 극심한 서귀포(보목과 새섬) 조하대에는 무절산호조류가 수심 4-5 m까지 번무하였으며 수심 6-7 m에서 감태 군락이 일부 관찰되었다(Chung et al., 1998). 우도에서 해조류의 연 평균 생물량은 약 3 kg wet wt/m2였으며 감태가 1-10 m의 모든 수심에서 높은 생물량을 보였다(Kang et al., 2011). 이처럼, 제주도의 조하대는 켈프인 감태와 무절·유절산호조류가 번무하는 것으로 나타났으나, 현재까지의 연구는 제주도 남쪽(서귀포 인근)과 동쪽(우도)에 관한 자료만 있을 뿐 서쪽과 북쪽에 대한 조하대 해조군집 및 수직분포에 대한 자료는 없는 상태이다. 더구나, 인간활동에 의한 기후변화와 환경오염, 그리고 관광객 증가 등의 다양한 환경요인 변화는 갯녹음 해역을 확대시키고 있으며(Chung et al., 1998), 기후변화로 인한 수온 상승은 열대성 해조류 출현종과 생물량의 증가 및 종다양성 감소와 같은 해조류 군집구조의 변화를 가져오고 있다(Carballo et al., 2002).
따라서, 본 연구는 우리나라 다른 해역에 비해 아열대화가 빠르게 진행되고 있는 제주도 동·서·남·북 조하대 해역의 해양환경특성과 연안생태계 먹이사슬에서 기초생산자인 해조류의 종 다양성, 자원량, 수직분포 등 군집구조를 분석하여, 산업화와 환경 오염 및 기후변화로 인한 갯녹음 발생, 열대성 해조류 번무, 군집구조의 단순화와 같은 연안생태계 변화를 예측하고 해조류 자원의 보존 및 관리방안 등을 위한 과학적인 자료를 제공할 목적으로 수행되었다.
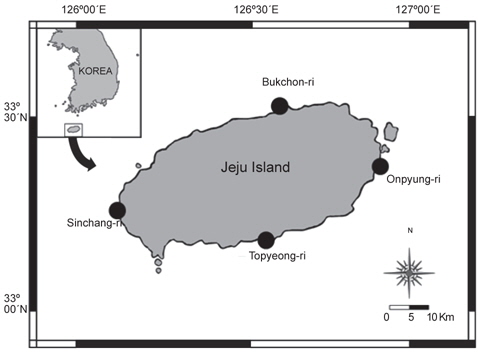
본 연구를 수행하기 위해 해조류 채집 및 해양환경 조사는 제주도의 동·서·남·북에 위치한 온평(33˚24΄N, 126˚54΄E), 신창(33˚21΄N, 126˚10΄E), 토평(33˚14΄N, 126˚34΄E), 북촌(33˚33΄N, 126˚41΄E)에서 2012년 3월부터 11월까지 계절별로 SCUBA Diving에 의해 실시되었다(Fig. 1).
각 정점별 표층과 저층(수심 10 m)의 수온, 염분은 YSI (6600-V2)를 이용하여 현장에서 측정하였으며, 해수 시료는 Niskin 채수기를 이용하여 저층에서 채집 후 냉장상태(4℃)를 유지하면서 실험실로 운반하여 분석하였다. 부유물질(SS)은 미리 무게를 잰 GF/F 여과지로 해수시료 2 L를 여과하고 110℃에서 항량으로 건조한 후 전자저울(CP124S, Sartorius)로 무게를 측정하였다. Chl-a는 해수시료 약 1 L를 GF/F 여과지로 여과한 후, 90% 아세톤 용액 10 mL를 첨가하여 엽록소를 추출한 다음 원심분리기로 분리된 상등액을 spectrophotometer (UVIKONxs, SCOMAM)를 이용하여 측정하였다(Parsons et al., 1984). 영양염류는 해양환경공정시험기준(MLTMA, 2010)에 따라 암모니아 질소(NH4+-N), 아질산 질소(NO2--N), 질산 질소(NO3--N), 인산 인(PO4-3-P)은 각각 Indophenol method, NED method, cadmium reduction method, Ascorbic acid method로 분석하였으며, 규산 규소[Si(OH4)-Si]는 Molibden blue method로 spectrophotometer (UVIKONxs, SCOMAM)를 이용하여 비색정량하였다. 용존무기질소(DIN)는 암모니아와 아질산 및 질산 질소의 합으로 나타냈다.
각 정점의 조하대 수심 5, 10, 20 m에서 10×10 cm로 구획된 방형구(50×50 cm)를 각각 2개씩 놓고 방형구 내에 출현하는 해조류를 끌칼을 이용하여 전량 채집하였다. 채집된 해조류는 현장에서 포르말린-해수 용액(5-10%)으로 고정시켜 실험실로 운반한 후 현미경을 사용하여 동정하였으며, 출현종 학명 및 국명은 Lee and Kang (2002)에 따랐다. 정량 채집된 해조류는 담수로 수회 세척하여 모래 및 불순물을 제거하고 종별로 습중량을 측정하였으며, 단위 면적당 생물량(g wet wt/m2)으로 환산하였다.
연구 정점별 출현종과 생물량으로 Margalef (1958)의 풍도지수(richness index, R), Fowler and Cohen (1990)의 균등도 지수(eveness index, J')와 Shannon's 다양도 지수(diversity index, H')를 계산하였다. 우점도 지수(dominance index, DI)는 방형구내 전체 생물량에 대한 제 1, 2 우점종 생물량의 합의 비로 산출하였다(McNaughton, 1967). 또한, 출현한 해조류의 우점도는 K-dominance 곡선으로 비교하였고(Lambshead et al., 1983), 해조류의 출현종 유∙무로 각 정점의 유사도(similarity)를 분석하였으며(Bray and Curtis, 1957), 그룹 간 유의차는 SIMPROF (similarity profile) tests를 실시하여 검정하였다. 그룹 간 유의차가 확인되면 SIMPER (Similarity percentage) 분석을 실시하여 각 그룹 간 비유사도(dissimilarity)에 기여하는 종을 확인하였다. 군집분석은 PRIMER version 6 (Clarke and Gorley, 2006)를 이용하여 산출 및 도식화 하였다.
연구 정점, 계절 및 수심별 생물량에 관한 통계분석은 Cochran의 등분산검정(test of homogeneity of variances)을 실시하여 데이터의 동질성이 확인되면 two-way ANOVA test를 실시하여 유의차를 검정하였고, 유의차가 발견되면 Tukey's HSD test 방법으로 사후 검정하였다(Sokal and Rohlf, 1995). 생물량 데이터는 통계분석을 위해 Log 변환(Log transformation)하였으며, 분석을 위해 사용된 통계 프로그램은 STATISTICA version 5.0 이었다.
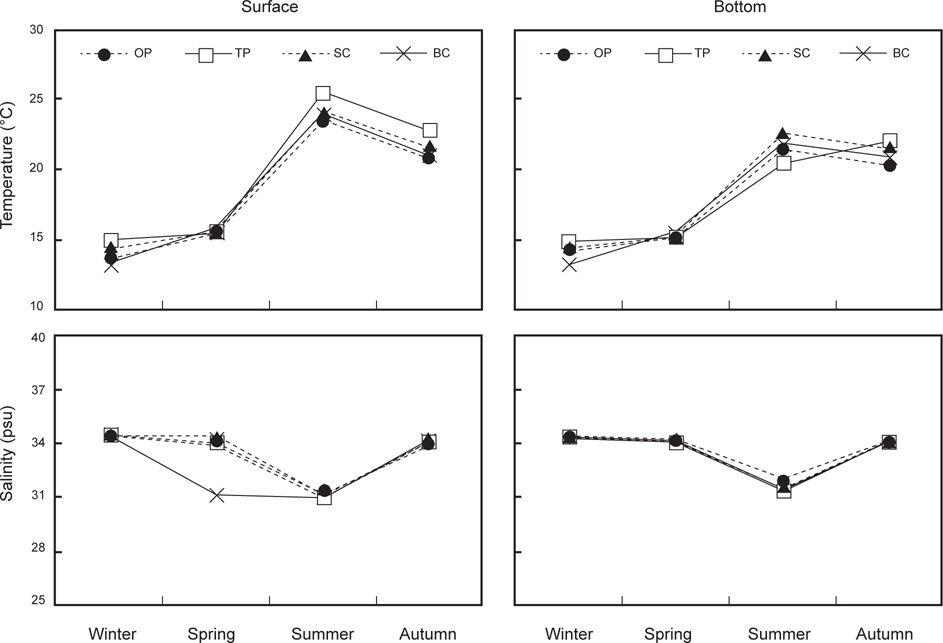
연구 정점 4개에서 표층 수온은 14.0-24.3℃ (평균 18.9±2.4℃)이고 저층 수온은 14.1-21.6℃ (18.0±1.9℃)로 겨울에 최저였고 여름에 최고였으며, 표층과 저층의 수온차는 여름에 최대로 나타났다(Fig. 2). 온평에서 표층 수온은 계절별로 13.6-23.5℃ (평균 18.4℃), 저층은 14.4-21.4℃ (17.7℃)로 겨울에 가장 낮았고 여름에 가장 높았다. 토평에서 표층은 14.9-25.5℃ (19.7℃)로 겨울에 최소였고 여름에 최대였지만, 저층은 가을 에 22.0℃로 최대 수온을 보였다. 신창에서 표·저층 수온은 각각 18.9℃ (14.4-24.2℃)와 18.4℃ (14.3-22.6℃)였으며, 북촌에서는 각각 18.5℃ (13.2-24.0℃)와 17.9℃ (13.2-21.9℃)로 모두 겨울에 최소였고 여름에 최대였다.
염분은 표층에서 평균 33.3 psu (31.2-34.5 psu), 저층에서 33.7 psu (31.7-34.5 psu)로 나타났으며, 계절별로는 수온과 반대로 표·저층 염분 모두 여름에 최소, 겨울에 최대로 확인되었다 (Fig. 2). 정점별 염분은 표층에서 32.7-33.6 psu로 북촌에서 최소였고 신창에서 최대였으며 모든 연구정점에서 여름에 최소였고 겨울에 최대였다. 저층 염분은 33.6-33.8 psu로 정점간 염분의 변화는 거의 없었으며, 계절에 따른 변화 양상은 표층 염분과 마찬가지로 여름에 최소였으며 겨울에 최대를 보였다.
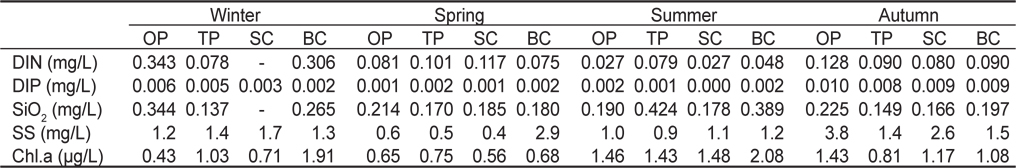
연구기간 동안 용존무기질소(DIN) 농도는 연평균 0.109 mg/L로, 계절별로는 0.045-0.242 mg/L의 범위로 여름에 최소, 겨울에 최대였으며, 정점별로 0.075-0.145 mg/L로 신창에서 최소, 온평에서 최대값을 보였다(Table 1). 용존무기인(DIP)의 경우 연평균 0.004 mg/L로, 계절별로는 0.001-0.009 mg/L의 농도로 여름에 최소, 가을에 최대였고, 정점별로는 0.003-0.005 mg/L로 정점 간 큰 차이는 없었으며, 온평에서 최대로 나타났다. 규산(SiO2)의 농도는 연평균 0.224 mg/L로, 계절별로는 0.184-0.295 mg/L로 가을에 최소, 여름에 최대로 확인되었고, 정점별로는 0.176-0.258 mg/L로 신창에서 가장 낮고, 북촌에서 가장 높았다. 부유물질(SS)의 경우 연평균 1.5 mg/L로 계절별로는 1.1-2.3 mg/L의 범위를 보이며 여름에 최소값, 가을에 최대값을 보였으며, 정점별로는 1.1-1.7 mg/L로 토평에서 최소, 북촌에서 최대였다. Chlorophyll-a의 경우 연평균 농도는 1.105 μg/L로 나타났으며, 계절별 분포 양상은 0.660-1.641 μg/L의 범위로 봄에 최소, 여름에 최대였고, 정점별로는 0.981-1.440 μg /L로 신창에서 최소, 북촌에서 최대값을 보였다.
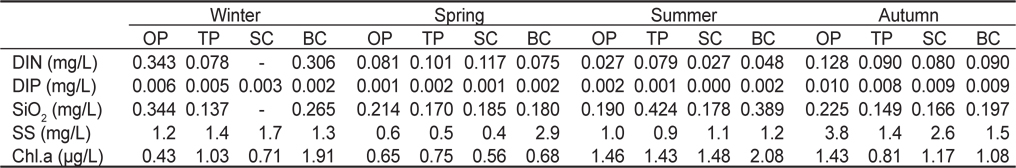
Seasonal variations of physical and chemical values observed in the bottom water at four study sites in Jeju Island, Korea, from March to November 2012
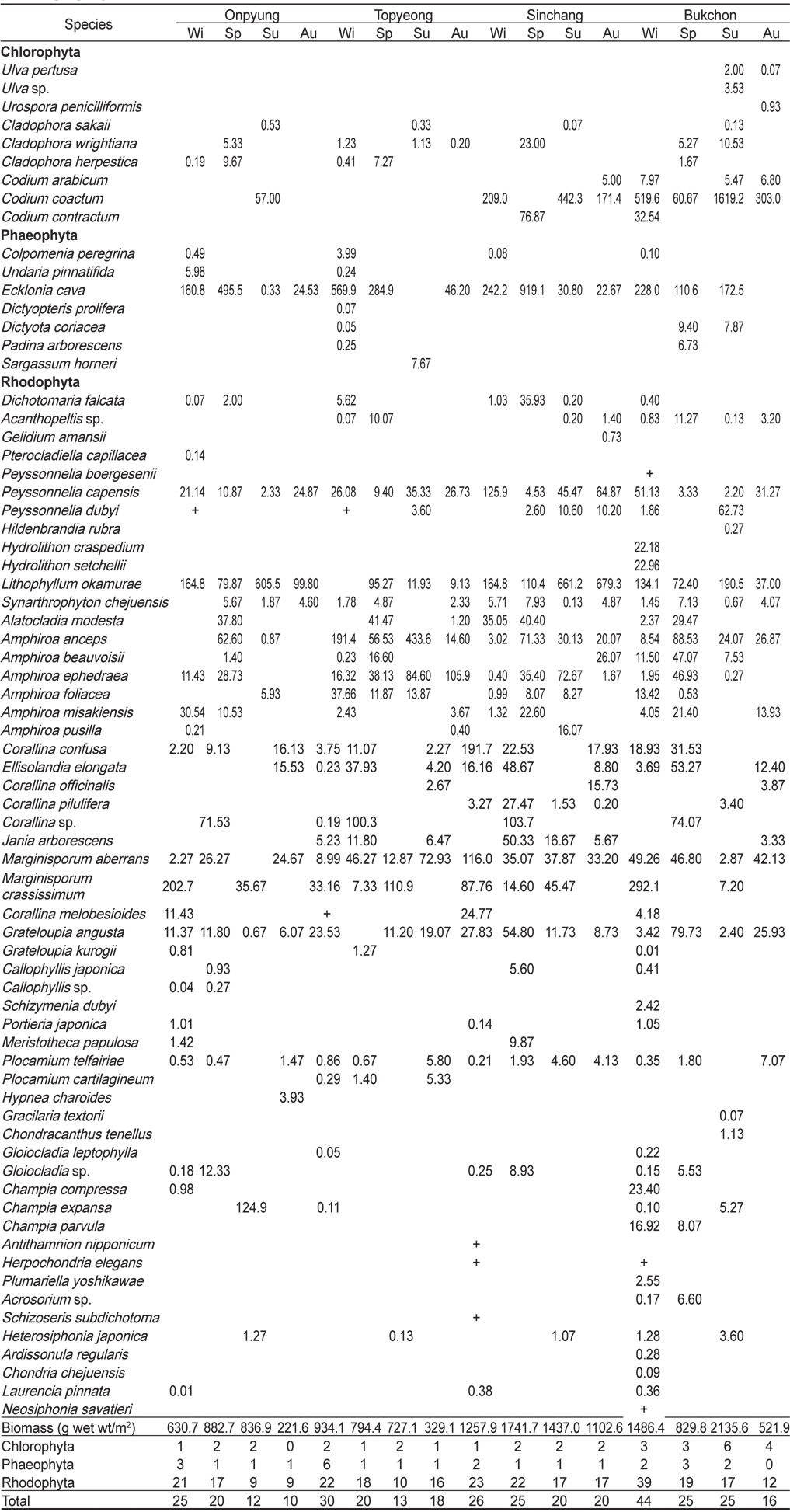
본 연구기간 동안 제주도 4개 정점에서 출현한 해조류는 총 71종으로 녹조류 9종(12.68%), 갈조류 7종(9.86%), 홍조류 55종(77.46%)이었으며, 계절별 출현종수는 28-60종으로 가을에 최소, 겨울에 최대로 나타났다(Table 2). 정점별 출현종수는 38-59종으로서 토평에서 가장 적었고 북촌에서 최대였다. 모든 정점에서 연중 출현한 해조류는 자루바다표고(
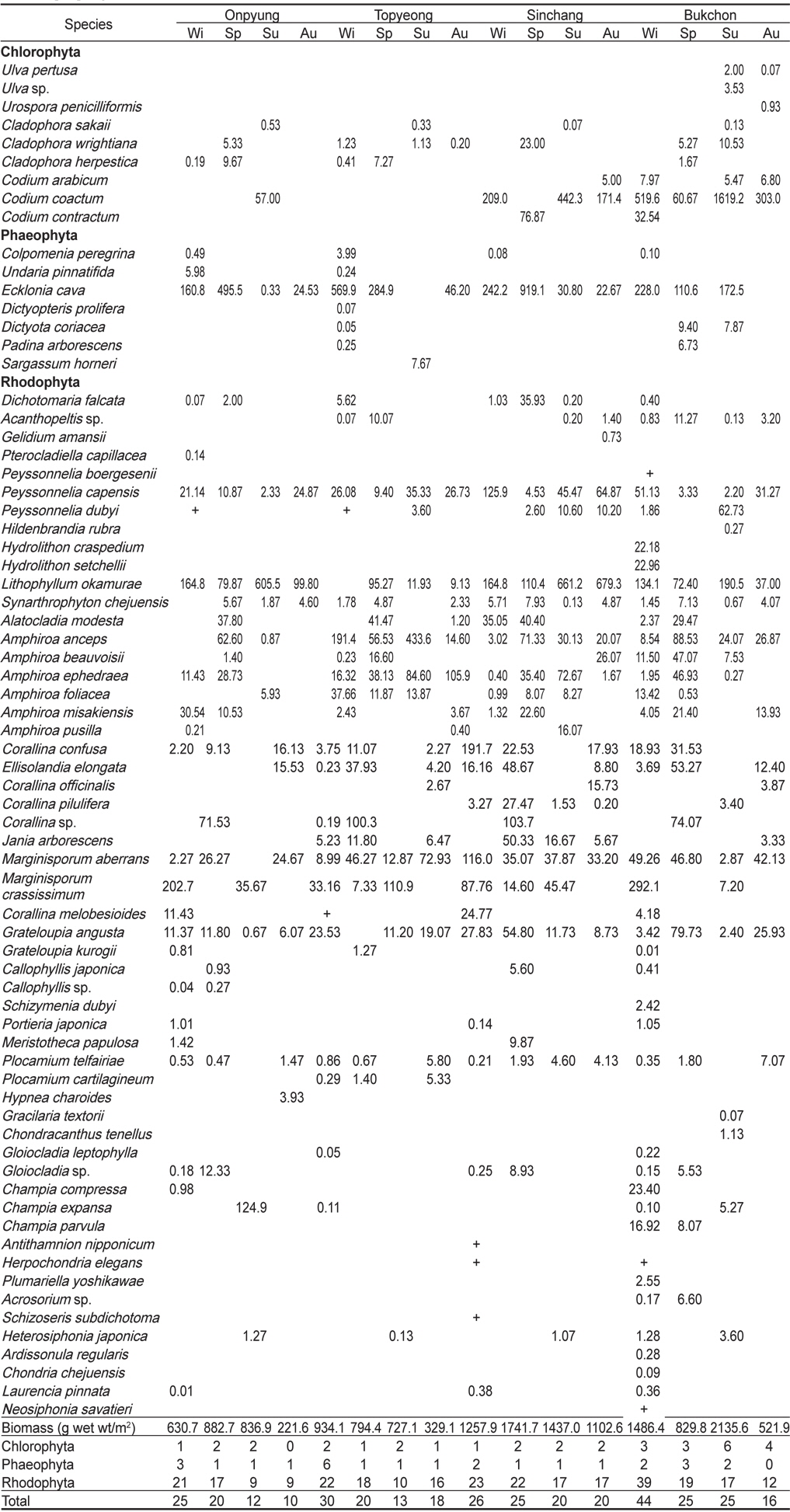
Marine algal lists and biomass (g wet wt/m2) occurred at four study sites in Jeju Island, Korea, from March to November 2012. Wi, Winter; Sp, Spring; Su, Summer; Au, Autumn
온평 연안에서 연간 출현한 해조류는 39종으로 녹조류 4종(10.26%), 갈조류 3종(7.69%), 홍조류 32종(82.05%)으로 홍조류의 비율이 가장 높았으며, 계절별로는 10-25종으로 가을에 최소, 겨울에 최대였다. 연중 출현한 종은 감태(
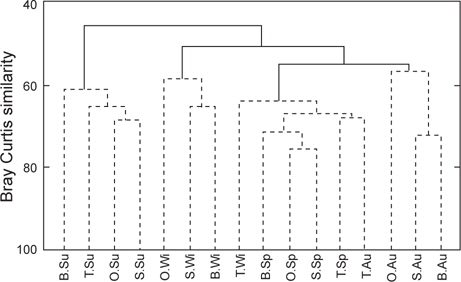
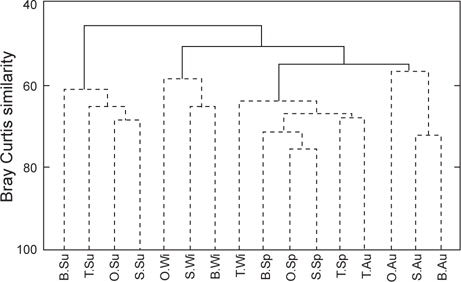
출현종의 유∙무로 유사도 분석을 실시한 결과, 통계학적으로 유의차를 보이는 4개 그룹 즉, 그룹 A (여름), 그룹 B (온평, 신창, 북촌의 겨울), 그룹 C (온평, 신창, 북촌의 봄, 토평 겨울, 봄, 가을), 그룹 D (온평, 신창, 북촌의 가을)로 구분되어 계절적인 차이를 보이는 것이 확인되었다(Fig. 3). 여름의 출현종(그룹 A)은 나머지 3개 그룹과 45.98%의 가장 낮은 유사도를 보였으며, 그룹 C와 D 사이의 유사도는 54.87%, C∙D 그룹과 B 그룹간의 유사도는 50.80%로 유의차를 나타냈다(SIMPROF test,
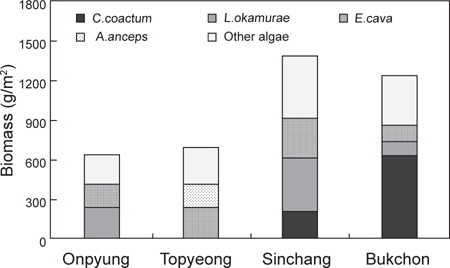
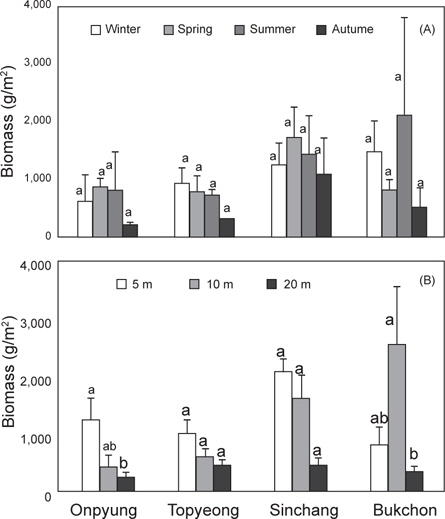
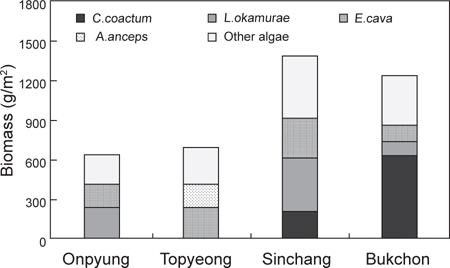
제주도 4개 해역(온평, 토평, 신창, 북촌)의 조하대 해조류 연평균 생물량은 991.84 g wet wt/m2이며, 분류군별로는 녹조류가 224.40 g/m2 (22.62%), 갈조류 209.43 g/m2 (21.12%), 홍조류가 558.01 g/m2 (56.26%)으로 홍조류가 최대 생물량을 보였다. 정점별 생물량은 642.96-1,384.79 g/m2로 온평에서 최소, 신창에서 최대였으며, 계절별 생물량은 543.80-1,284.17 g/m2 로 가을에 최소, 여름에 최대였다(Table 2). 4개 정점의 계절별 생물량을 two-way ANOVA로 분석한 결과 정점(F3,32=2.21,
수심별 연평균 생물량은 온평에서 241.33-1,258.77 g wet wt/m2, 토평에서 473.64-1,014.78 g/m2, 신창에서 477.06-2,076.65 g/m2으로 수심 20 m에서 최소였고, 5 m에서 최대였으며, 북촌에서는 343.93-2,561.78 g/m2으로 수심 20 m에서 최소, 10 m에서 최대였다. 4개 정점에서의 수심별 생물량을 twoway ANOVA로 분석한 결과 정점(F3,36=4.39,
제주해역의 해조류 중에서 연간 평균 생물량이 가장 높은 종은 누운청각(211.39 g/m2, 21.31%)이었으며, 그 다음은 감태(206.75 g/m2, 20.85%), 혹돌잎(194.75 g/m2, 19.64%) 순으로 나타났다. 각 정점별로는 온평에서 혹돌잎(237.49 g/m2)과 감태(170.29 g/m2)가 전체 생물량(642.96 g/m2)의 각각 36.94%, 26.48%를 차지하며 우점하였다(Fig. 5). 조사시기에 따라서는, 겨울에는 둘레게발혹, 혹돌잎, 감태가, 봄에는 감태 1종이, 여름에는 혹돌잎, 넓은사슬풀, 가을에는 혹돌잎 1종이 생물량을 구성하는 우점종으로 확인되었다. 토평 해역에서는 감태(225.24 g/m2)와 넓은게발(174.03 g/m2)이 평균 생물량(696.18 g/m2)의 각각 32.35%, 25.00%의 비율을 보였으며(Fig. 5), 계절별 우점종은 겨울에는 감태, 넓은게발, 봄에는 감태,
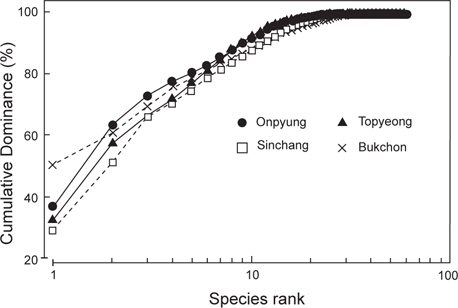
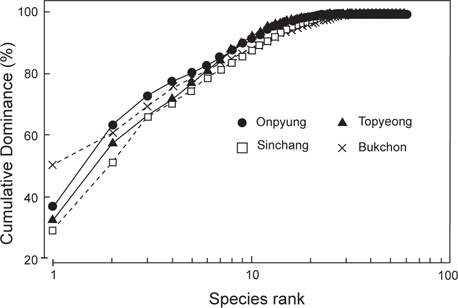
한편, 정점별로 출현한 종의 연평균 생물량을 k-dominance 곡선으로 비교하면, 우점종 3종의 생물량이 신창(혹돌잎, 감태, 누운청각)과 토평(감태, 넓은게발, 둥근띠게발)에서는 전체 생물량의 70% 이하를 차지하고 북촌(누운청각, 감태, 혹돌잎)에서는 70% 정도를 차지하였으며, 온평에서는 우점종 3종인 혹돌잎, 감태, 둘레게발혹이 전체 생물량의 70% 이상을 나타냈다 (Fig. 6).
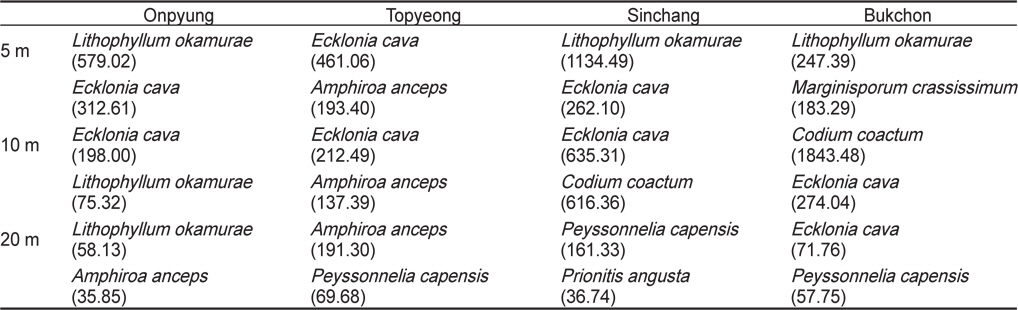
연평균 생물량을 토대로 한 우점종의 수직분포를 분석한 결과, 온평에서는 혹돌잎·감태(5, 10 m) - 혹돌잎·넓은게발(20 m)로 혹돌잎이 전 수심에서 우점하였으며, 토평 해역은 수심 5 m와 10 m에서 감태·넓은게발이, 수심 20 m에서는 넓은게발· 자루바다표고로 확인되어 넓은게발이 전 수심에서 우점하여 나타났다. 신창에서는 혹돌잎·감태(5 m) - 감태·누운청각(10 m) - 자루바다표고·붉은뼈까막살(20 m)의 수직분포를 보였으며, 북촌의 수심 5 m에서는 혹돌잎과 둘레게발혹이, 10 m에서는 누운청각과 감태, 20 m에서는 감태와 자루바다표고가 우점하였다. 녹조류인 누운청각은 수심 10 m, 홍조류인 자루바다표고는 수심 20 m에서 높은 생물량을 보였다(Table 3).
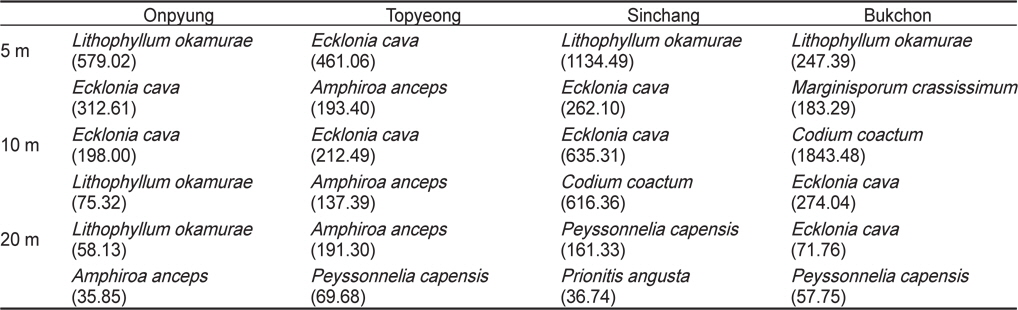
Vertical distribution of dominant and subdominant seaweed based on mean biomass (g wet wt/m2) at four study sites in Jeju Island, Korea
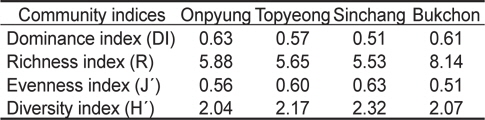
정점별로 4계절에 출현한 해조류의 평균 생물량으로 계산한 우점도지수(DI)는 0.51-0.63으로, 온평에서는 제 1, 2 우점종인 혹돌잎(237.49 g/m2)과 감태(170.29 g/m2)가 전체 생물량(642.96 g/m2)의 63.42%를 차지하여 최대였고, 신창에서는 온평과 마찬가지로 제 1, 2 우점종인 혹돌잎(403.91 g/m2)과 감태 (303.69 g/m2)가 전체 생물량(1,384.79 g/m2)의 51.10%를 차지하여 최소로 확인되었다. 풍도지수(R)는 5.53-8.14로 북촌에서 최대, 신창에서 최소였으며, 출현종수와 생물량에 따라 변화하는 균등도지수(J')는 풍도지수와는 반대로 신창에서 최대(0.63), 북촌에서 최소(0.51)로 나타났다. 출현종수와 생물량의 분포인 균등도지수에 의해 결정되는 다양도지수(H')는 2.04-2.32로 나타나 온평에서 최소, 신창에서 최대였다(Table 4).
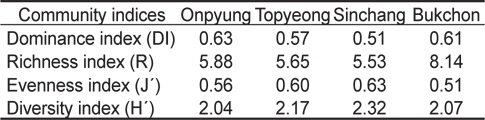
Various community indices of macroalgal flora for mean biomass (g wet wt/m2) at four study sites in Jeju Island, Korea
제주 해역은 대마난류수, 중국대륙연안수, 황해저층냉수, 한국남해연안수 등 다양한 수괴들의 영향으로 복잡한 해양환경을 가지며, 대마난류수, 기후변화 등으로 인해 열대 및 아열대성생물들이 제주 연안에 서식하여 종다양성이 높은 독특한 생태계를 보이고 있다(Yang et al., 2007). 이러한 특징 때문에 제주도 해양생물 조사가 많은 학자들에 의해 활발히 진행되고 있으며(Yang et al., 2007), 해조류 또한 오래 전부터 군집구조에 대한 연구가 수행되어 왔다(Lee and Lee, 1976; Park et al., 1994; Lee et al., 1998; Yoo, 2003; Kang and Kim, 2012). 본 연구에서는 제주도 4개 해역에서 총 71종(녹조 9종, 갈조 7종, 홍조 55종)의 해조류가 출현하였고, 정점별로는 38-59종으로 북부해역인 북촌에서 가장 많은 출현종수를 보였다. 이는 제주 8개 해역(온평, 위미, 법환, 대포, 화순, 한림, 외도, 함덕)의 조하대에서 출현한 70-109종(Lee et al., 1998)에 비해 낮게 나타났으며, 출현종수가 가장 많은 북촌을 제외한 3개 정점의 출현종수는 Oak et al. (2004)이 숲섬에서 보고한 41종, Ko et al. (2008)이 문섬에서 보고한 46종과 유사하였다. 최근에는 제주도에 속한 추자도에서 162종(Kim et al., 2008), 우도에서 262종(Kang et al., 2011), 마라도에서 144종(Kang and Kim, 2012)으로 많은 종이 출현하여 본 연구결과와 현저한 차이를 보였다. 본 조사에서 해조류 생물량(습중량)은 991.84 g/m2이었으며, 문섬(2,784 g/m2, Ko et al., 2008), 우도(2,934.6 g/m2, Kang et al., 2011), 마라도(2,932.3 g/m2, Kang and Kim, 2012)와 같은 제주도 주변 도서에 비해 낮았으나, 이는 2012년 여름의 대형 태풍(볼라벤과 산바 등)으로 인해 암반이 뒤집히면서 해조류의 탈락 및 유실, 제주 연안에 발생한 갯녹음 현상(Chung et al., 1998), 양식장에서 대량의 유기물 유입(Delgado et al., 1997) 및 해양레저 관광객 증가에 의한 생태계 교란(Davenport and Davenport, 2006) 등 다양한 환경요인의 복합적인 작용으로 판단된다. 특히, 제주 동·남부는 대마난류의 영향을 가장 많이 받는 해역으로 수온이 높아 제주도 양식장의 60% 이상이 위치해 있고, 해 양레저 이용객 수가 북부에 비해 약 5배 정도 높았으며(Chung and Kim, 2012), 해조류 생물량도 북부의 북촌(1,243.42 g/m2)과 서부의 신창(1,384.79 g/m2)에서 동부와 남부에 위치한 온평 (642.96 g/m2)과 토평(696.18 g/m2)에 비해 2배 정도 높게 나타남으로써 기후변화(수온상승, 해양산성화)와 더불어 환경요인이 해조군집의 변화와 관련됨을 암시한다. 하지만, 본 연구에서 환경자료와 해조류의 생물량에서는 뚜렷한 패턴이 보이지 않았는데, 이는 연구 해역에 대한 환경 자료가 축적되지 않았기 때문으로 사료된다. 따라서, 기후변화와 환경변화가 본 연구 해역에 서식하는 해조류의 생물다양성 및 생물량에 어떠한 영향을 미치는지를 파악하기 위한 장기적인 모니터링 연구가 요구된다.
조하대 해조류 군집은 대형 갈조류가 생물량의 대부분을 차지하고 있는데, 제주도 연안은 그 중에서 감태와 모자반류가 주요종의 역할을 하고 있다(Oak et al., 2004; Ko et al., 2008; Kang et al., 2011). 본 연구에서 생물량을 근거로 한 최우점종은 신창과 북촌에서만 우점하였던 누운청각(211.39 g/m2)이었으나, 감태(127.79-303.69 g/m2, 평균 206.75 g/m2)는 모든 정점에서 높은 생물량을 보여 조사해역의 해중림을 구성하는 주요종이라할 수 있다. 또한, 연구 정점에서 공통적으로 무절산호조류인 혹돌잎의 생물량(194.75 g/m2)이 높게 나타났는데, 이는 숲섬과 성산포 해역에서 감태와 무절산호조류가 우점한다는 이전의 연구 결과와 일치하였다(Oak et al., 2004). 수심별 우점종은 혹돌잎·감태(5 m) , 누운청각·감태(10 m)로서 감태가 수심 5-10 m에서 우점하는 것으로 확인되었으며, 수심 20 m에서는 그늘진 조하대 암반이나 어두운 곳과 같은 약한 광에서 생육하는 것으로 알려진 자루바다표고가 우점하였다.
수온은 해조류의 수평분포를 결정하는 중요한 환경요인으로 알려져 있으며(van den Hoek, 1982), 수온의 상승은 이들의 지리적 분포를 변화시킨다고 하였다(Tenger and Dayton, 1987; Serisawa et al., 2004). 제주 연안의 표층 수온은 지난 86년(1924-2009년) 동안에 약 1.94℃ 상승하였으나(Suh et al., 2011), 해조류 생물량에 대한 이전의 자료가 부족하여 수온상승과 해조군집 변화의 관계를 명확하게 설명한다는 것은 거의 불가능하다. 하지만, 제주에서 채취되는 해조류(톳, 우뭇가사리) 생산량과 수온 변동을 연계시켜 보면, 수온은 최근 20년(1991-2010년)에 약 0.64℃ 상승하였고, 톳 생산량은 3,751톤(1990년)에서 360.6톤(2010년)으로 90%정도 감소하였으며, 우뭇가사리는 동일 기간에 약 86% 감소하였다. Serisawa et al. (2004)은 일본 Tosa만에서 70년대 이전과 이후(80, 90, 2000년대) 수온이 지속적으로 상승하는 것을 확인하였고, 이로 인해 70년대 이전에 서식했던 감태 군락이 완전히 소멸하였음을 보고하였다. 또한, Kim (2006)은 수온 변동 추세가 현재 상태로 유지된다면 제주 연안의 주요종인 감태가 향후 10년 뒤에도 해중림을 형성할 수 있을지는 예측할 수 없으며, 이를 대체할 남방계 해조류의 탐색이 필요하다고 하였다. Kang (1966)은 수온, 조류 등 해황을 바탕으로 한국 연안을 5개 해역(북동해구, 남동해구, 동남해구, 서해구 및 제주구)으로 구분하였고, 제주구의 해조류 조성이 북방계 2%, 온대계 74%, 남방계 10% 및 범세계종 15%로 되어 있다고 하였다. 최근, Kang et al. (2011)이 우도의 해조상에서 남방계 해조류가 37.80% 출현한다고 밝혀 그 출현비율이 현저하게 증가하였다고 밝혔였다. Kang et al. (2011)이 사 용한 방법(http://www.algaebase.org, Guiry and Guiry, 2013)으로 본 연구에서 출현한 해조류 중 남방계 해조류는 21종의 29.58%로 Kang et al. (2011)의 결과에 비해 다소 낮게 나타났으나, 특히, 엷은잎바위주걱(
결론적으로 본 연구가 수행된 제주도 4개 연안에서 서식하는 해조류는 총 71종이며, 가을에 가장 적은 출현종수를 보였다. 해조류의 연평균 생물량은 991.84 g wet wt/m2이며, 여름에 1,284.17 g/m2으로 가장 높게 나타났다. 생물량을 기준으로한 우점종은 누운청각, 감태, 혹돌잎이었으며, 4정점 모두 생물량이 높게 나타난 감태가 제주도 연안의 대표적 주요종이라 할 수 있다. 이러한 해조류의 종조성 및 생물량은 이전의 제주 주변해역의 연구 결과에 비해 다소 낮았으나 동·서·남해의 다른 정점들에 비해 높게 나타나 현재에도 제주도 해조류의 종다양성은 높다는 것이 확인되었다. 또한, 아열대성 남방계 해조류 출현율이 약 50년 이전에 비해 19-27%가 증가되는 것으로 확인되어 기후변화는 연안생태계의 기초생산자인 해조류의 생물다양성과 생물량 감소와 더불어 군집구조를 변화시킬 것으로 생각되므로 조사 정점에 대한 지속적인 모니터링 연구가 필요하다.