A feeding trial was conducted to determine the optimum dietary protein and lipid levels for the growth of juvenile muddy loach
성장이 빠르고 건강한 양식 어류를 생산하기 위해서는 먼저 대상종의 필수영양소 요구에 대한 연구를 수행하여 그 종에 적합한 배합사료를 개발하여야 한다. 필수 영양소 중에서 단백질은 어류의 성장에 가장 많은 영향을 미치는 중요한 요소일 뿐 아니라, 단백질 원료 가격이 비싸기 때문에 사료 단가에도 영향을 미칠 수 있다(Lovell, 1989). 일반적으로 육식성 어류에서 사료 내 단백질 함량의 증가는 어류 생산성을 향상시키지만 단백질 함량이 증가 할수록 사료단가는 높아진다. 어류의 성장을 위한 단백질의 이용성은 지질이나 탄수화물과 같은 비 단백질 에너지원으로 사료 단백질을 부분적으로 대체함으로서 향상될 수 있다(Lee et al., 2002). 하지만, 과잉의 사료 에너지는 사료 섭취를 감소시켜 성장을 위한 필수 영양소의 섭취가 부족할 수 있다. 이와 반대로, 사료 내 비단백질 에너지가 불충분하면 체내에서 단백질이 에너지로 사용되어 단백질이 낭비될 수 있을 뿐 아니라 단백질이 에너지로 전환될 때 발생되는 암모니아는 수질을 악화시킨다. 따라서 섭취된 사료 단백질이 체내에서 단백질 합성 외의 목적으로 사용되지 않도록 사료 단백질 이용성을 향상시키는 것은 중요하다. 배합사료의 단백질을 절약하기 위해 비단백질 유래의 에너지 함량을 증가시키는 연구가 여러 어종에 대해 연구되고 있다(NRC, 2011). 특히, 지질은 에너지가(value)가 높은 영양소로 초식성 어류보다 잡식성이나 육식성 어류들에서 더 쉽게 이용되며, 필수지방산 공급원으로 양식어의 정상적인 성장과 체내 대사에 필수적인 역할을 하는 영양소이다(Kim et al., 2002). 사료 내 비 단백질 에너지원의 종류나 함량은 어류의 성장, 영양소 이용률, 체지방 조성 등에 영향을 미치므로, 이에 대한 연구가 필요하다(Garling and Wilson, 1976; Daniels and Robinson, 1986; Sargent et al., 1989; Cho and Kaushik, 1990).
미꾸리(
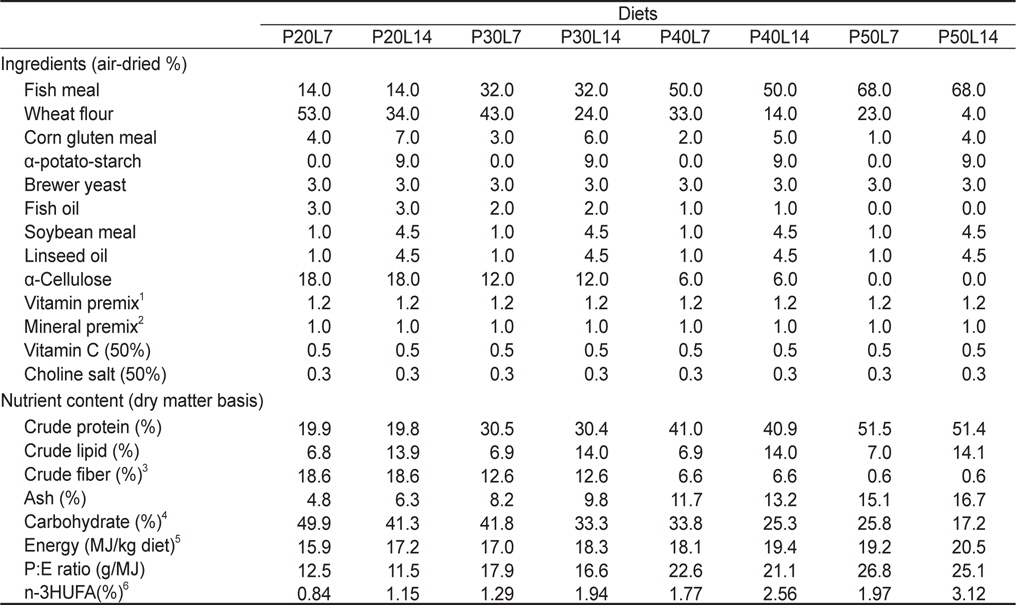
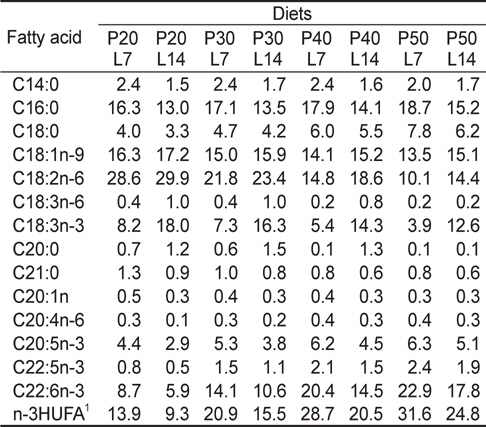
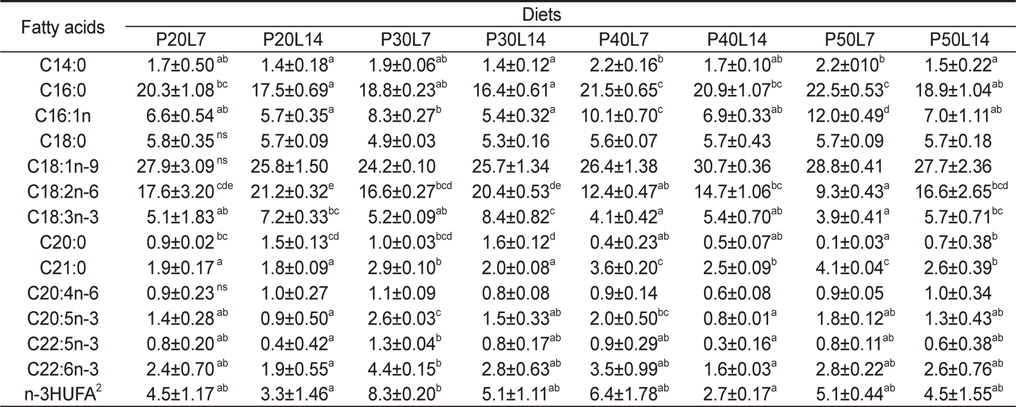
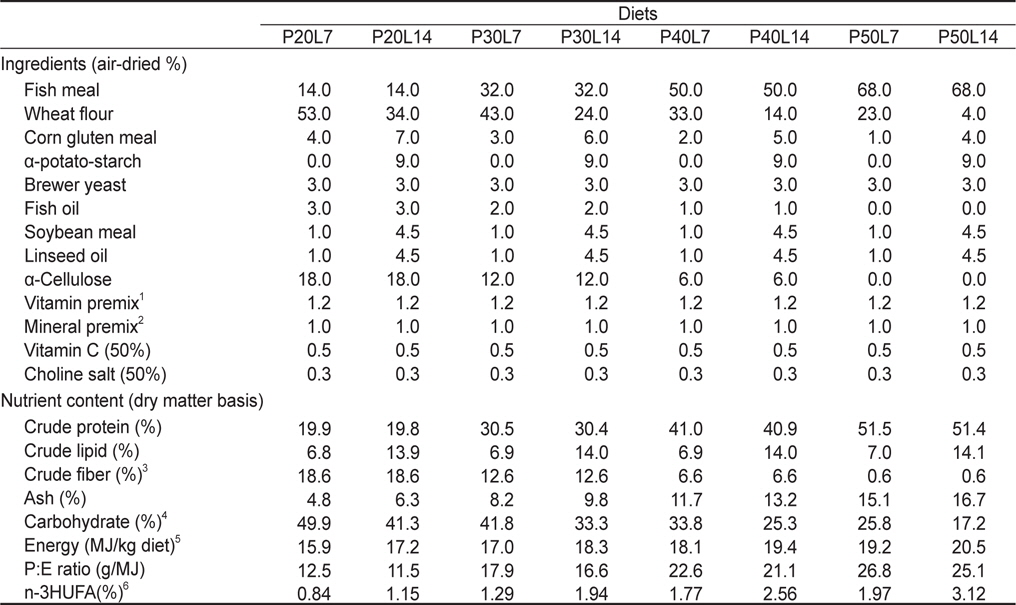
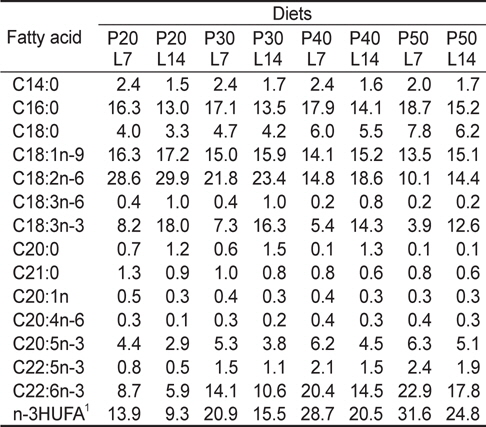
실험사료의 원료조성은 Table 1에 나타내었다. 4×2 factorial 디자인으로 4종류의 단백질 함량(20%, 30%, 40% 및 50%)과 2종류의 지질 함량(7% 및 14%)을 함유하고 있는 총 8종류의 실험사료를 제조하였다. 실험사료의 단백질원으로 어분을 지질원으로 어유, 대두유 및 아마인유를 그리고 탄수화물원으로 소맥분을 각각 사용하였고, 사료의 단백질과 지질 함량은 배합되는 원료들의 비율에 따라 조절되었다. 이와 같이 설계된 원료들을 잘 혼합한 후 원료 100 g 당 물 30 g을 첨가하여 moist pellet 제조기로 압출 성형한 후, 실온에서 24시간 건조하였다. 그리고 실험사료를 실험어가 섭취할 수 있는 크기의 pellet size로 절단하여 -30℃에 보관하면서 공급하였다. 실험사료의 지방산 조성은 Table 2에 나타내었다.
[Table 1.] Ingredient and proximate composition of experimental diets
Ingredient and proximate composition of experimental diets
[Table 2.] Fatty acid composition (% of the total fatty acids) of the experimental diets
Fatty acid composition (% of the total fatty acids) of the experimental diets
실험어로 국립수산과학원 내수면양식연구센터에서 종묘생산된 미꾸리 치어를 사용하였다. 2주간 실험수조에서 실험어를 순치시킨 후, 외형적으로 건강한 평균체중 1.49±0.10 g의 미꾸리 치어를 선별하여 총 24개의 20 L 사각 수조에 각각 30마리씩 3반복으로 수용하여 8주간 사육하였다. 사육기간 동안 수온은 24.5±0.2˚C였으며, 각 수조마다 약하게 폭기시켜 산소를 공급하였다. 실험사료는 1일 2회(9:00, 17:00) 실험어가 먹을 때까지 손으로 만복 공급하였다. 사육기간 중 죽은 개체는 매일 제거하였으며, 하루에 한번(13:00) 환수하면서 실험수조를 청소하였다. 일일사료섭취율과 폐사어 등은 매일 기록하였다.
어체 측정은 사육실험 시작시와 종료시에 측정 전일 절식시킨 후 tricaine methanesulfonate (MS 222, Sigma, St. Louis, MO, USA) 100 ppm 수용액에 마취시켜 실험어 전체무게를 측정하였다. 실험사료, 어체의 일반성분은 AOAC (1990)의 방법에 따라 조단백질(N×6.25)은 Auto Kjeldahl system (Buchi B-324/435/412, Switzerland; Metrohm 8-719/806, Swizerland)을 사용하여 분석하였고, 조지방은 ether 추출법을 사용하였으며, 수분은 105˚C dry oven에서 6시간 동안 건조 후 측정하였고 회분은 600˚C 회화로에서 4시간 동안 태운 후 측정하였다. 지방산 분석은 Folch et al. (1957)의 방법에 따라 클로로포름과 메탄올 혼합액(2:1)으로 총 지질을 추출하여 14% BF3-methanol (Sigma, USA) 용액으로 지방산을 methylation 시킨 후, capillary column (SP™-2560, 100 m×0.25 mm i. d., film thickness 0.20 um, USA)이 장착된 gas chromatography (Perkin Elmer, Clarus 600, USA)로 지방산을 분석하였다. Carrier gas는 헬륨을 사용하였으며, oven 온도는 최초 140˚C에서 240˚C까지 4˚C/min 증가시켰다. 이때, injector 온도는 240˚C, detector (FID) 온도는 240˚C로 각각 설정하였으며, 표준 지방산으로 37개 지방산 혼합물(PUFA 37 Component FAME Mix, USA)을 사용하였다.
결과의 통계처리는 SPSS Version 20.0 (SPSS Inc., Chicago, IL, USA) program을 사용하여 One-way ANOVA-test를 실시한 후, Duncan’s multiple range test (Duncan, 1995) 로 평균간의 유의성(
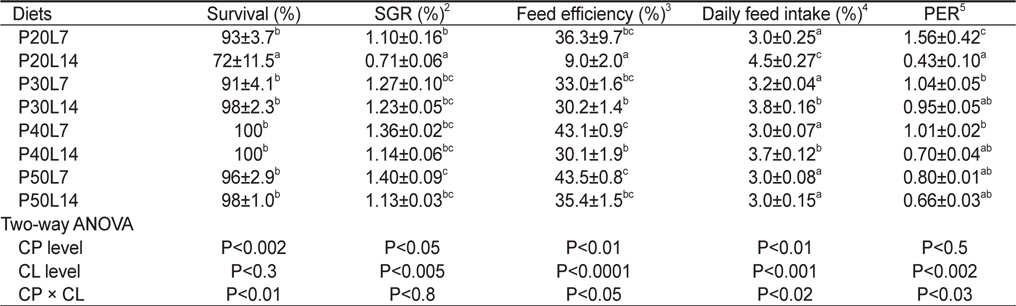
사육실험이 끝난 후, 8주간의 성장 및 사료이용효율을 Table 3에 나타내었다. 사육기간 동안의 생존율은 사료 내 단백질 함량(
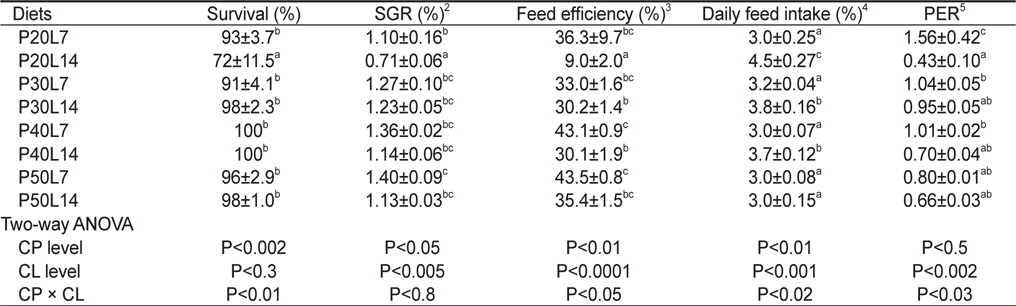
Growth performance of juvenile muddy loach Misgurnus anguillicaudatus (initial body weight, 1.49±0.10 g/fish) fed the experimental diets containing different protein (CP) and lipid (CL) levels for 8 weeks1
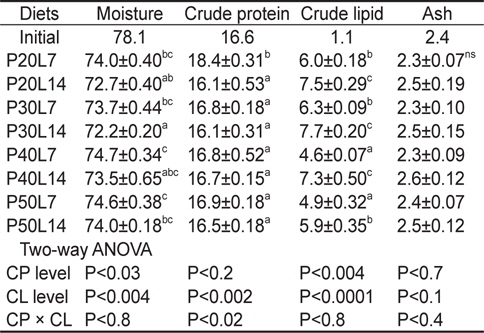
전어체의 일반성분 분석 결과를 Table 4에 나타내었다. 전어체의 수분 및 지질 함량은 사료 내 단백질 및 지질 함량에 영향을 받았고, 단백질은 사료 지질 함량에 영향을 받았다. 전어체의 수분 함량은 같은 단백질 함량에서 지질 함량이 증가할 때 감소하는 경향이 나타났으며, P40L7 및 P50L7실험구가 P20L14와 P30L14 실험구보다 유의하게 높았다. 전어체의 단백질은 P20L7실험구가 가장 높은 값을 나타내었다(
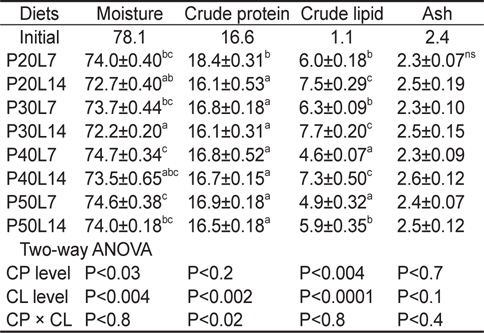
Proximate composition (%) of the whole body in juvenile muddy loach Misgurnus anguillicaudatus fed the diets containing different protein (CP) and lipid (CL) levels for 8 weeks1
전어체의 지방산 조성 변화를 Table 5에 나타내었다. C16:0, C18:1n-9, C18:2n-6, C18:3n-3 및 n-3HUFA (highly unsaturated fatty acids)의 함량은 사료의 C16:0, C18:1n-9, C18:2n-6, C18:3n-3 및 n-3HUFA와 비슷한 경향으로 조성되어 있는 것으로 보아 미꾸리 체내 지방산 조성은 실험사료 지방산 조성에 직접적인 영향을 받는 것으로 나타났다. 동일한 사료 단백질 함량에서, 사료 내 지질함량이 증가함에 따라 C18:2n-6 및 C18:3n-3의 함량은 증가한 반면, C20:5n-3 및 C22:6n-3과 같은 n-3HUFA 의 함량은 감소하는 경향이 나타났다.
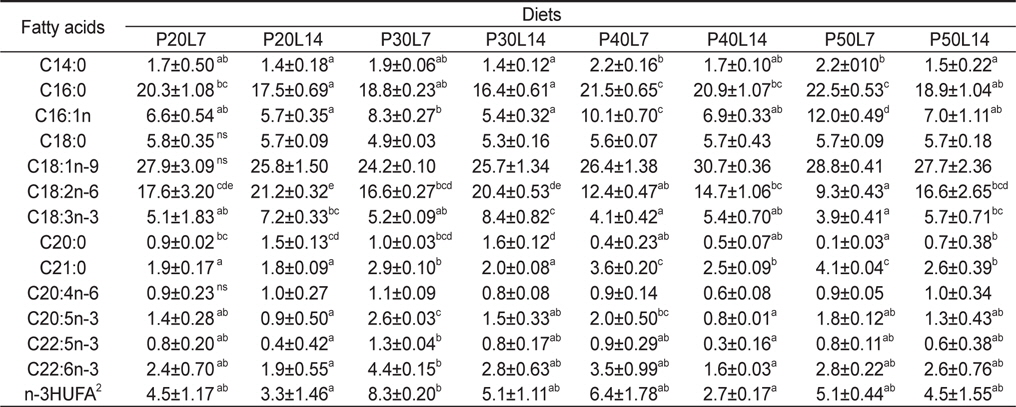
Fatty acid composition of whole body in juvenile muddy loach Misgurnus anguillicaudatus fed the diets containing different protein and lipid levels for 8 weeks1
본 연구에서는 사료 지질 14% 실험구의 성장율은 단백질 30%까지는 유의하게 증가하였으나 그 이상에서는 더 이상 증가하지 않았고, 사료 지질 7%실험구의 성장율도 단백질 30%이상에서 유의한 차이가 없었다. 따라서, 미꾸리 치어 양식을 위한 배합사료에는 단백질을 30% 함유 시키는 것이 바람직할 것으로 판단된다. 이와 같은 미꾸리 단백질 요구량이 30%로 비교적 낮은 것은 미꾸리의 식성이 잡식성임을 암시한다. 또한, 미꾸리의 성장과 사료효율을 고려하여 볼 때, 미꾸리 치어용 배합사료의 지질 함량은 14%보다 7%로 함유시키는 것이 더 좋을 것으로 판단된다. 미꾸리는 논두렁이나 늪 같은 곳에 주로 서식하면서 실지렁이, 곤충의 유충, 식물의 어린싹, 녹조류 및 부식 유기물 등을 섭취한다. 이처럼 미꾸리가 섭취하는 먹이의 지질 함량이 높지 않은 것으로 미루어 보아 미꾸리의 지질 이용성은 높지 않음을 알 수 있고, 본 연구 결과도 이를 뒷받침하고 있다. 어류의 단백질 요구량은 어종에 따라서 달라질 뿐 아니라, 어체의 크기, 수온, 사료 급여 방법, 사료 단백질의 품질 및 에너지 함량 등 여러 요인에 영향을 받을 수 있는 것으로 보고되어 있다(NRC, 2011). 타 담수어종의 단백질 요구량을 살펴보면, 송어(
본 연구에서 사료효율이 예상보다 낮게 나타났는데, 이러한 현상에 대한 정확한 원인을 현재로서 규명할 수는 없지만, 아마도 본 연구에 사용된 미꾸리의 생리적인 특성에 의한 것으로 보인다. 예를 들어, 미꾸리는 바닥에 쌓인 부식유기물을 쉽게 섭취할 수 있도록 진화된 입의 구조와 소화기관 등에 의한 특성 때문에 나타나는 현상일 수 있다고 판단된다.
본 연구에서 배합사료의 지질 함량 증가에 의한 단백질 절약효과는 나타나지 않았는데, 이러한 현상은 미꾸리의 경우 사료 지질 이용성이 타 어류에 비해 낮거나, 사료에 첨가된 지질의 종류가 이 어종의 기호성에 맞지 않았을 수도 있었기 때문으로 판단된다. 다른 어종의 연구에서도 사료의 지질 함량 증가에 의한 단백질 절약효과가 없었다는 결과들이 보고된바 있다(Berge and Storebakken 1991; Lee and Kim, 2005; Regost et al., 2001). 또한, 사료 내 과잉의 지질 함량은 지방산 합성을 억제시킬 뿐 아니라 성장을 감소시킬 수 있고(Sargent et al., 1989), 여분의 에너지가 체내 지질로 축적되어 어체 품질에 영향을 줄 수 있으므로, 이를 고려하여야 한다(Lee and Kim, 2005). 본 연구에서도 사료의 지질 함량이 높은 실험구의 전어체 지질 함량이 낮은 지질 함유 사료 실험구보다 유의하게 높았다. 이러한 경향은 사료의 에너지 함량과 밀접하게 관련되어 있는 것으로 보이며, 고에너지 사료를 공급한 어류의 지질 함량이 저에너지 사료를 공급한 어류보다 더 높았던 다른 연구결과와 일치한다(Lie et al., 1988; Hillestad and Johnsen, 1994; Catacutan and Coloso, 1995).
일반적으로 사료 내 지질 종류 또는 함량은 대상 생물의 지방산 조성에 영향을 미친다고 보고되어 있다(Geurden et al., 1997; Lee and Lim, 2005). 동일한 사료 단백질 함량에서 사료 내 지질 함량이 증가함에 따라 18:2n-6과 18:3n-3의 함량은 증가한 반면, 20:5n-3 및 22:6n-3과 같은 n-3HUFA 함량은 낮아졌다. 이러한 결과는 실험사료의 지질 함량을 높이기 위해 18:2n-6 및 18:3n-3이 많이 함유된 대두유 및 아마인유 첨가에 따른 결과로 보이며, 다른 어종에서도 유사한 경향을 보이고 있다(Lee et al. 2000; Bowden et al., 2004; Piedecausa et al., 2007). 미꾸리의 필수지방산 요구량은 알려지지 않았지만, 본 연구에 있어 n-3HUFA 요구량을 고려하여 사료 내 어분 및 어유를 사용하였으며, 본 연구에서 사용된 실험 사료의 n-3HUFA 함량은 0.8-3.1%으로 대부분의 담수어류의 적정 성장에 필요한 사료의 지방산 요구량의 범위 내에 있다(NRC, 2011). 금후에 미꾸리의 필수지방산 요구에 관한 상세한 연구가 계속 수행되어야 할 것으로 판단된다.
본 연구의 결과로 볼 때, 미꾸리의 경우 사료의 지질 함량의 증가는 사료 단백질 절약에 대한 효과를 나타내지 않았으며, 단백질 30%와 지질 7%가 함유된 배합사료가 미꾸리 치어의 성장에 적합할 것으로 전망된다.