Erebidae is one of four quadrifid noctuoid families recently redefined by Zahiri et al. (2011) and Zaspel et al. (2012). A clade corresponding to Erebidae was first recognized by a molecular study (Mitchell et al., 1997). Later, Fibiger and Lafontaine (2005) verified the monophyly of Erebidae in terms of morphology but retained Arctiidae and Lymantriidae as separate families. These two groups were readjusted as erebid subfamilies in the latest phylogeny of Noctuoidea (Zahiri et al., 2011), and by van Nieukerken et al. (2011). Erebidae, as currently defined, comprises 1,760 genera and 24,569 species (van Nieukerken et al., 2011), representing the largest family of the Lepidoptera.
There exists no previous estimate of species diversity for the Korean erebids. Kononenko and Han (2007) critically updated the Korean Noctuidae
The purposes of this paper is to report two erebid genera and species new to Korea, to describe the female genitalia of
Specimens were dissected for genitalia, following the method of Clarke (1941), except that chlorazol black was used for staining, and specimen dissections were mounted on microslides in euparal resin. Pinned specimens were examined under a Leica MZ APO stereo zoom microscope(Leica, Wetzlar, Germany), and slide-mounted specimens under a Leica LEITZ-DMRX compound microscope. Photos of adults were made with a Nikon D40 DSR camera(Nikon, Tokyo, Japan). Genital images were captured using the VDBK digital imaging systems, adopted by the United States Department of Agriculture and installed in the Department of Entomology, USNM. Terminology for genitalia follows that of Klots (1970). All examined specimens are deposited in the Department of Plant Medicine, Chungbuk National University, Cheongju, Korea.
Genera and species new to Korea
Order Lepidoptera Family Erebidae
Luceria Walker, 1859 Luceria Walker, 1859: 853. Type species: Luceria novatusalis Walker, 1859, by original designation.
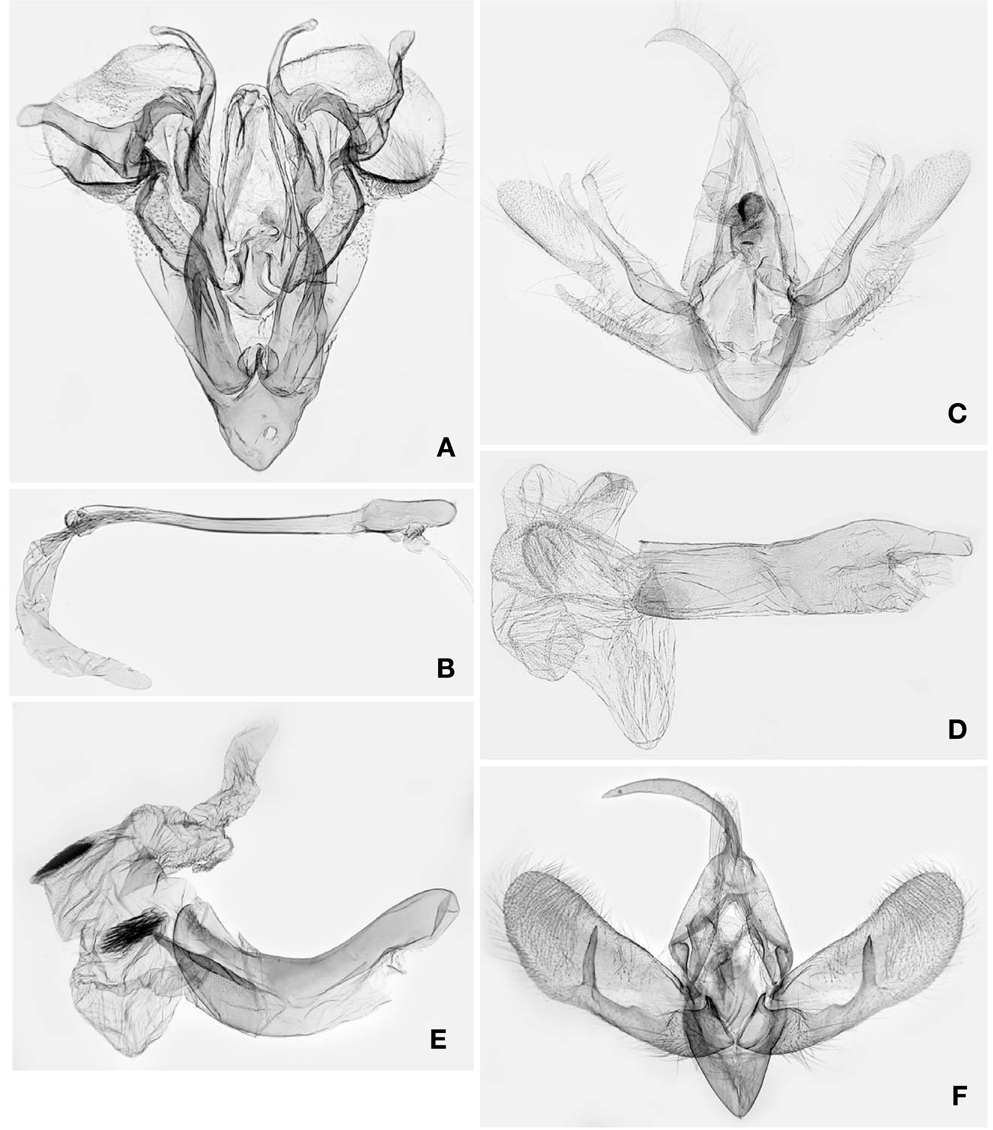
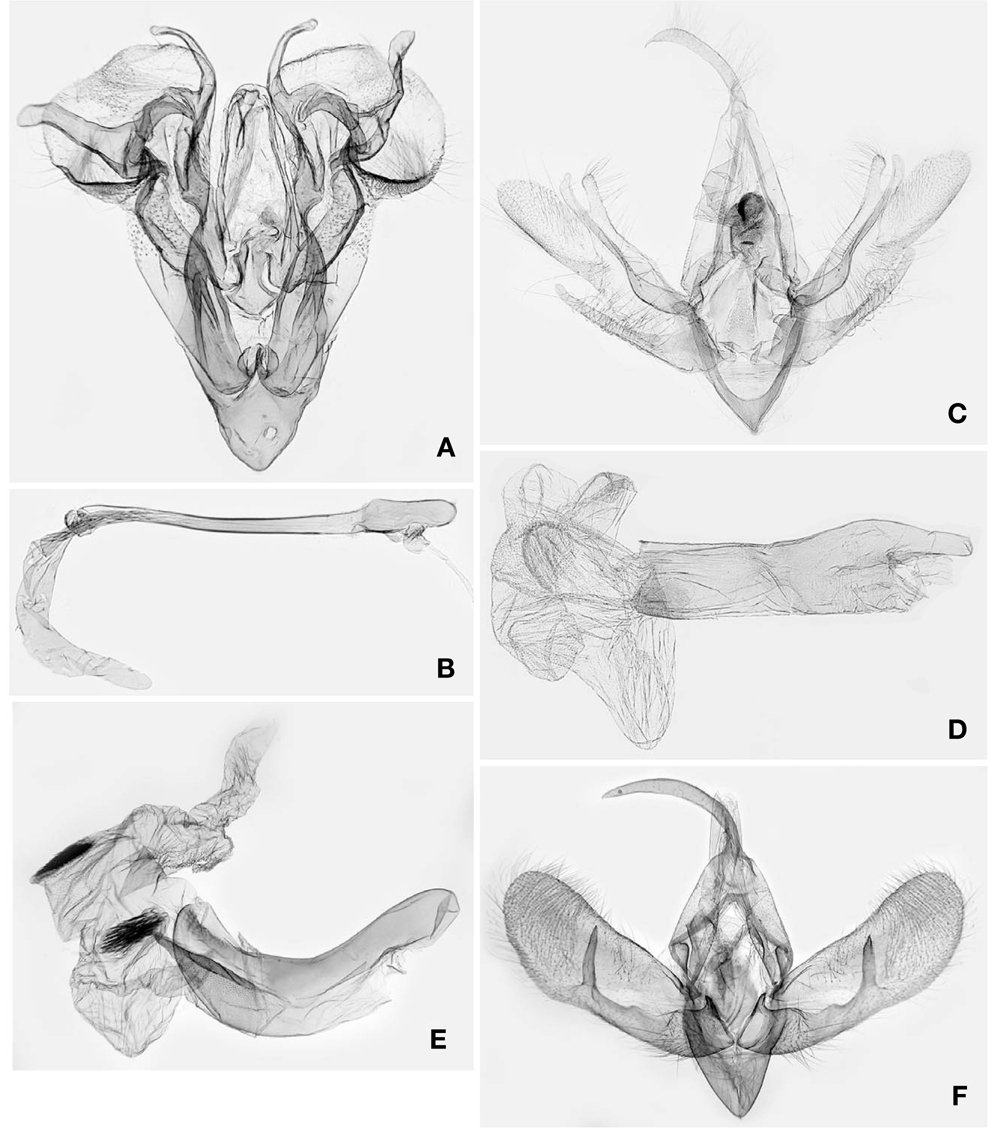
1*Luceria striata Galsworthy, 1997 (Korean name: sae-wu-zzal-leum-na-bang) (Figs. 1A, 2A, B, 3A)
Material examined. Korea: 1♂, Chungbuk Prov., Cheongju, Heungdeok,Sannam-dong, Mt. Guryongsan, 26 Aug 2005, Sohn JC, by bucket trap, SJC-381; 1♀, Gyeongnam, Isl. Geojae-do, Jangmok, Jangmunpo-weaseong(castle), 34˚ 59′30.8′′N, 128˚ 40′25.2′′E, 18 Jun 2004, Sohn JC, by light trap, SJC-743.
Diagnosis. This species is superficially similar to
Redescription. Head (Fig. 1A): Vertex and frons brownish white, sparsely intermixed with dark brown scales. Antennae filiform, pubescent ventrally in both sexes; scape brownish white, intermixed with a few dark brown scales; flagellomeres brownish white dorsally, naked ventrally. Labial palpi oblique upward, 3rd segment upcurved; 1st segment broadened to apex, 1/5 as long as 2nd segment, dark brown on exterior surface, brownish white on interior surface; 2nd segment dark brown on exterior surface, brownish white on interior surface, with shallow-triangular, pale brown scale tuft dorsally; 3rd segment 1/4 as long as 2nd, acuminate apically, dark brown, with brownish white ring apically and basally.
Thorax (Fig. 1A): Patagium and tegula pale yellow, intermixed with dark brown scales exterolaterally; mesonotum brownish white, sparsely intermixed with dark brown scales. Fore- and mid-legs dark grayish brown dorsally, brownish white ventrally. Hindlegs brownish white, intermixed with pale brownish gray scales in distal 1/3 of femur. Forewing length 6.5-6.8mm (n=2) elongate-triangular, brownish white, peppered with dark brown scales denser along costal and outer marginal areas and between ante- and postmedian lines, termen broadly round in anterior 2/3; antemedian line dark brown, oblique in posterior 1/3, intermittent, curved in anterior 2/3; median line as an oblique, dark brown bar near costa; discal spot small, dark brown; postmedian line dark brown, nearly straight, oblique toward apex; submarginal shade triangular broadened to tornus; terminal dashes black; fringe dark brown. Hindwing pale brownish gray; marginal line dark brown; fringe pale yellowish gray in basal 1/2, dark brownish gray in distal 1/2. Male genitalia (Fig. 2A, B): Uncus 2/5 as long as tegumen, triangular in basal half, slender in distal half. Tegumen narrow, inverted U-shape. Valva narrow-triangular in basal 3/5; cucullus elliptical, with setose, semicircular emargination basoventrally; costal process arising from base, sinuous, band-like, with slender, curved, clubbed process subapically, looped with saccular process distally; sacculus entirely covering the basal 1/3 of valva, broadened medially, with curved, digitate bulge in distal half; saccular process at distal end of sacculus crescentiform, with one long and one short digitate lobes. Juxta inverted spade-shape. Vinculum narrowly diverging to tegumen; saccus clypeiform. Phallus slender, nearly straight, coecum broadened; vesica elongate, with a small, dentate cornutal zone basally. Female 7th sternite and genitalia (Fig. 3A): Sternite VII triangular. Papillae anales subtrapezoidal. Apophyses posteriores as long as papillae anales. Ostium bursae beyond the posterior end of 7th sternite. Ductus bursae narrow, 5× longer than corpus bursae, broadened at posterior 1/3 and anterior 1/4, sclerotized in posterior 1/4(antrum). Corpus bursae globular. Distribution. South Korea and China (Hong Kong).
Metachrostis Hübner, 1816 [1820] Metachrostis Hübner, 1816 [1820]: 104. Type species: Noctua velox Hübner, [1813], by subsequent designation by Hampson(1894: 325). = Leptosia Guenée, 1841: 225. Type species: Noctua velox Hübner, [1813], by subsequent designation by Guenée (1852: 237). A junior homonym of Leptosia Hübner, 1818 (Lepidoptera: Pieridae). = Neoleptosia Koçak, 1980: 38. An unnecessary replacement name for Leptosia Guenée, 1841.
This genus belongs to Eublemminae (Kononenko and Matov, 2009). Among eublemmine genera,
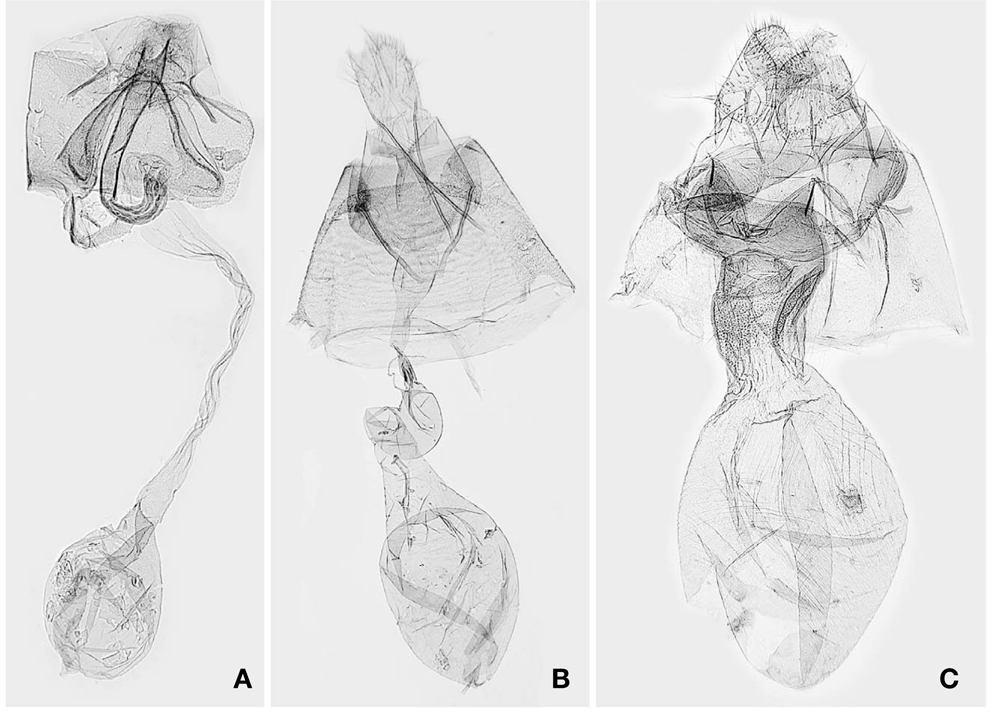
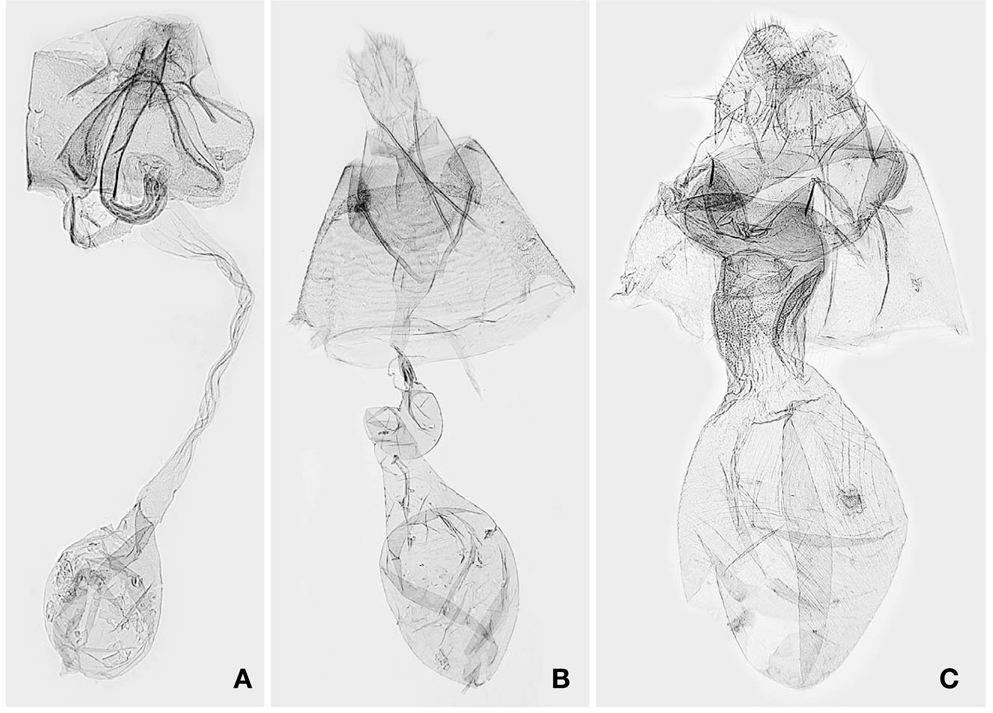
1*Metachrostis sinevi Matov et Kononenko, 2009 (Korean name: mung-dduk-nal-gae-zzal-leum-na-bang) (Figs. 1B, 3B)
Material examined. Korea: 1♀, Chungbuk Prov., Cheongju, Heungdeok-gu, Mt. Guryongsan, 29 Jun 2005, Sohn JC, SJC-748.
Diagnosis. Among the Korean erebids,
Description. Kononenko and Matov(2009) provided a detailed description for
Distribution. South Korea and Russia (Far East).
Species little known in Korea
1*Lophomilia nekrasovi Kononenko et Behounek, 2009 (Korean name: buk-bang-jul-zzal-leum-na-bang) (Figs. 1C, 2C, D, 3C) Lophomilia sp.: Kononenko, 2005: 63. Lophomilia nekrasovi Kononenko et Behounek, 2009: 10 (type locality: Russia, Slavyanka).
Material examined. Korea: 1♂, Chungbuk Prov., Gaeteojae, 31 Jul 2001, Cho S, Lyu DP, Nam S; 1♂, Gyongnam Prov., Gimhae-gun, Saengrim-myeon, Yeocha-ri, Yongsan Elementary School, 14-15 Aug 1987, paratype of L.
Diagnosis.
Description. The habitus and genitalia of
Distribution. South Korea, China (North), and Russia (Far East).
Notes. Kononenko and Behounek (2009) mentioned one male paratype from our collection. Since they did not give any detail about the specimen, it is impossible to find out which of the aforementioned specimens they referred to. Here, one male collected on 14-15 Aug 1987 is defined as the paratype.
1*Acontia martjanovi (Tschetverikov, 1904) (Korean name: sa-gu-ggo-ma-bam-na-bang) (Figs. 1D, 2E, F) Erastria martjanovi Tschetverikov, 1904: 78 (type locality: Russia, Minusinsk). Lithacodia martjanovi: Warren, 1912: 279. Lithacodia martjanovinovi(sic): Hua, 2005: 231. Acontia martjanovi: Kononenko, 2010: 141.
Material examined. Korea: 2♂, Chungnam Prov., Taean, Wonbuk, a sand dune near Sindu-ri, 3 Jun 2003, Sohn JC, SJC-318; 3♂, ditto, 21 Sep 2004, Sohn JC.
Diagnosis. This species is clearly distinguished from other congeners in Korea by having a large, white reniform stigma in dark brown forewings. In the male genitalia,
Description. Head brownish white. Antennae 3/5 as long as forewing. Labial palpi dark grayish brown, with a brownish white ring at end of second segment. Thorax with patagium, tegula, and mesonotum brownish white, intermixed with dark grayish brown scales; mesoscutellum with reddish brown scale tuft. Legs dark grayish brown; tibia and tarsomeres with brownish white ring distally. Forewings dark grayish brown; subbasal and antemedian, and postmedian line black, waved, juxtaposed with pale brown band; median area pale grayish brown; subterminal line pale yellowish brown, zigzagged; terminal dashes black, juxtaposed with pale yellowish brown dashes; orbicular stigma dark gray; reniform stigma white. Hindwing dark yellowish brown; marginal shade dark grayish brown. Abdomen dark brown, intermixed with pale yellow scales. Male genitalia with uncus elongate, curved; valvae elongate, round apically; harpe straight, at distal end of sacculus; vinculum narrowly diverged, narrowly round anteriorly; phallus curved; vesica with two spinulate cornutal zones. See Kononenko (2010) for the female genitalia.
Distribution. South Korea, China (Heilongjiang, Inner Mongolia), Mongolia, and Russia (South Siberia).
Notes. Kononenko (2010) included ‘S Korea’ in the distribution of this species without giving any specimen record. This species is known as xerophilous (Kononenko, 2010). In Korea,