Zooplankton distribution is strongly influenced by water body environmental variables, such as abiotic factors (e.g., water temperature, salinity, stratification, and advection), biotic factors (e.g., food limitation, predation, and competition), or a combination of both (Roff et al. 1988, Escribano and Hidalgo 2000, Beyst et al. 2001). Therefore, recognition of environmental variables in the distribution of zooplankton has been central to limnological research. In temperate regions, environmental factors in water bodies change dramatically according to the season, which determines the seasonal ontogeny and population growth of zooplankton (Marneffe et al. 1996).
Among diverse environmental factors, water temperature is clearly altered by seasonality, and controls metabolic rates and circadian rhythms, such as mating and dormancy (Hirche 1987, Robinson et al. 1983). Low water temperatures decrease the metabolism of zooplankton, which reduces their life cycle and feeding. The aforementioned circumstances affect their predators (mainly fish) as well (Brandt 1993), and hence, winter is believed to be a unique period during which zooplankton are not interrupted by predators. Therefore, we hypothesize that zooplankton communities in winter are affected by different environmental variables when compared with other seasons (spring, summer, and autumn). In particular, withered vegetation does not reduce phytoplankton biomass through shading effects (Sand‐Jensen and Søndergaard 1981) and allelopathy (van Donk and van de Bund 2002), and should thus support a large abundance of not only food sources, but also zooplankton. Unfortunately, distribution patterns of zooplankton related to environmental variables during winter have been insufficiently studied.
The primary objective of our study was to investigate the distribution pattern of zooplankton during winter. We investigated the following: (1) the influence of diverse environmental variables on zooplankton distribution, and (2) the contribution of withered vegetation to support a large abundance of zooplankton.
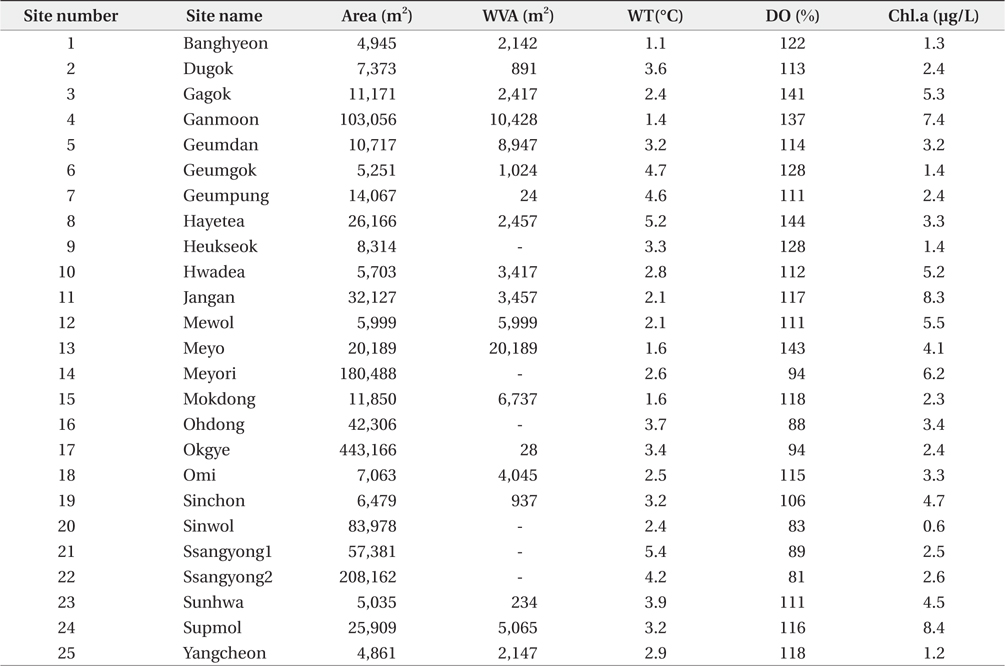
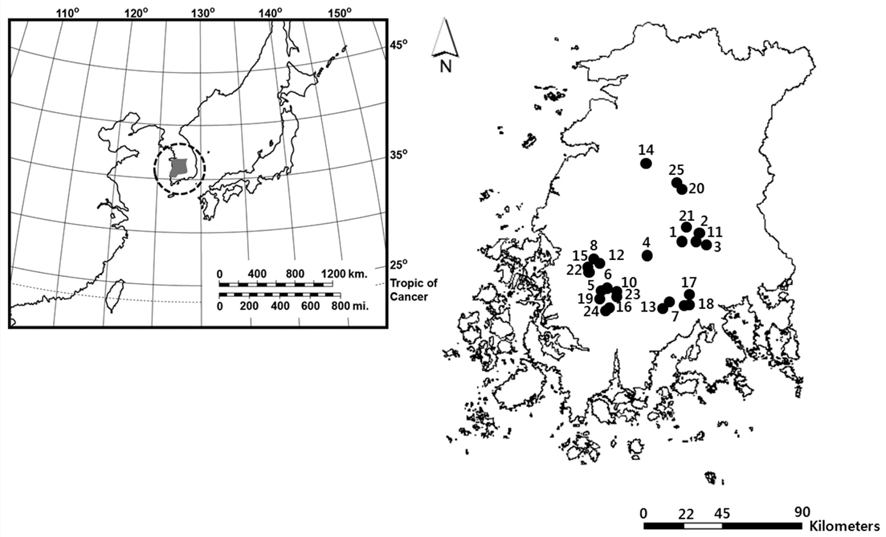
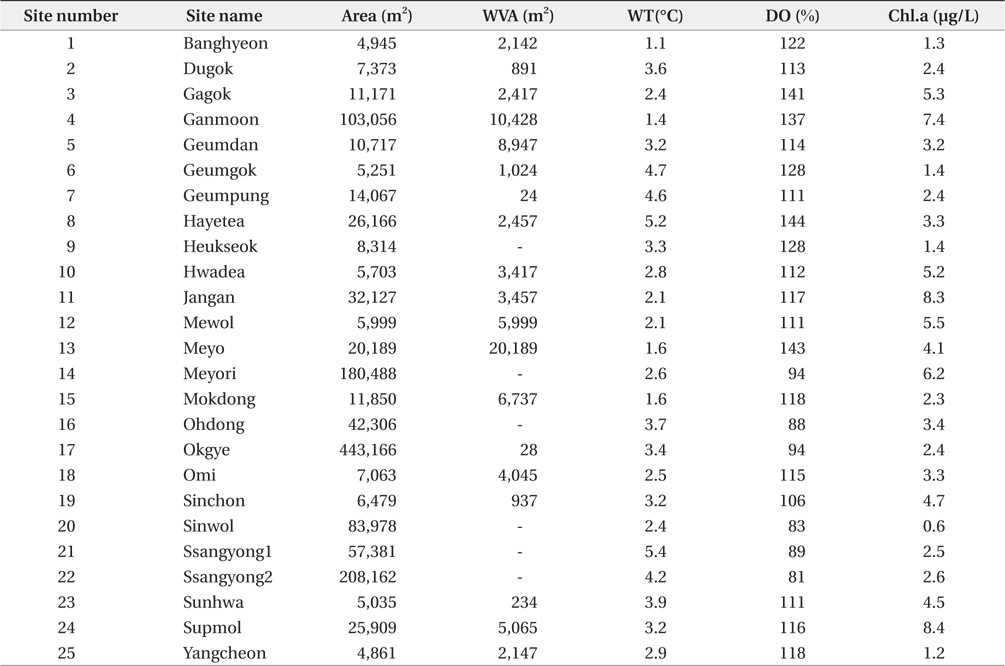
South Korea is located in East Asia, and possesses a temperate climate. Four distinct seasons lead to a dynamic succession of the biological community in Korean freshwater ecosystems. In particular, this country experiences an average temperature of −0.8°C during winter (from December to January). The reservoirs monitored in this study are located in southwestern Korea, within the central and lower reaches of the Yeongsan River (Fig. 1). We investigated environmental parameters (reservoir area, withered vegetation area, water temperature, dissolved oxygen, and chlorophyll a) and zooplankton communities in 25 reservoirs during winter (December 2010 to January 2011). We established three sampling points in littoral zone of each reservoir. The sampling points were randomly selected based on virtual grids constructed over maps of the reservoirs. We used a YSI 58 dissolved oxygen (DO) meter (YSI Inc., Yellow Springs, OH, USA) to detect water temperature and DO (% saturation). Chlorophyll a concentration was measured using a spectrophotometer in accordance with Wetzel and Likens (2000). Reservoir area and withered aquatic plant area were measured using ESRI’s ArcGIS ver. 9.3 (ESRI Inc., Redlands, CA, USA) and a digital map from National Geographic Information Institute (http://www.ngii.go.kr) with a scale of 1 to 25,000.
For zooplankton collection, we collected 5 L water samples using a 10 L column sampler at each sampling point. The sampled water was filtered through a plankton net (32-μm mesh net), and the filtrate was preserved in formaldehyde (final concentration: 5%). The zooplankton were identified and counted using an Axioskop microscope 40 (Carl Zeiss, Oberkohen, Germary) at ×200 magnification, based on the classification key published by Mizuno and Takahashi (1999).
Canonical correspondence analysis (CCA) was used to identify the relationship between the zooplankton communities and environmental parameters by using Canoco ver. 4.5 (Ter Braak and Smilauer 1998).
The observed levels for environmental factors (reservoir area, withered vegetation area, water temperature, dissolved oxygen level, and chlorophyll a level) at study sites are listed in Table 1. Even though some reservoirs showed exceptionally high or low relative values for these parameters, the coefficients of variation (CV) were less than 100%. The average water temperature ranged from 1.1 to 5.4°C, while DO (81% to 144%) and chlorophyll a (1.7 to 12.5 μg/L) showed wider ranges than that of the water temperature. The reservoir area and withered aquatic plant area showed the largest ranges of 4,861–443,166 ha and 0–20,189 ha, respectively.
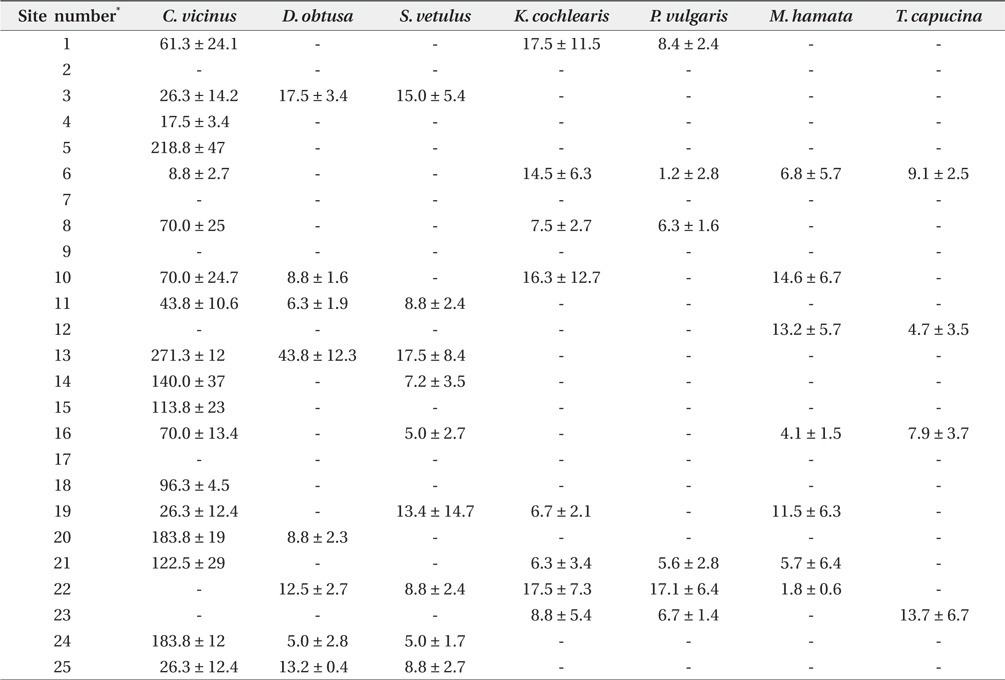
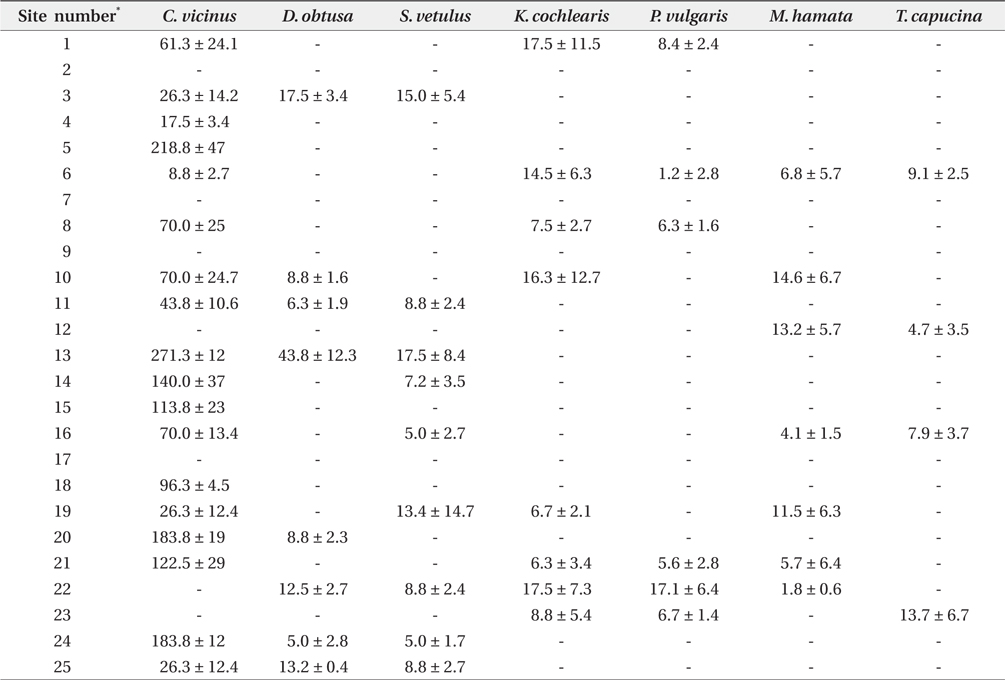
Zooplankton were collected across all investigated coreservoirs, but no zooplankton was found in the sites 2, 7, 9, and 17 (Table 2). Cyclops vicinus was predominant among the zooplankton groups, with densities of other zooplankton species (i.e., cladocerans and rotifers) being relatively low. The highest density of C. vicinus was found at site No. 13 (271 individual/L on average), followed by No. 5 (218 individual/L on average). The densities of other zooplankton did not differ between study sites
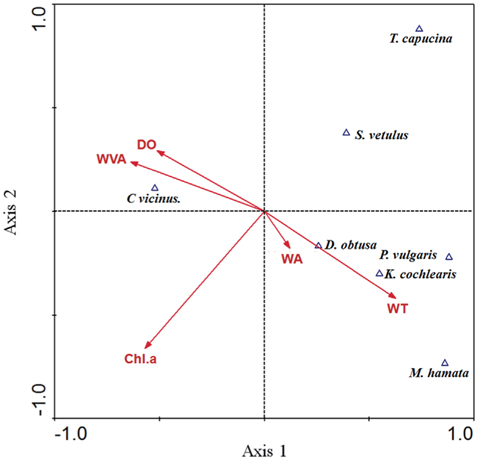
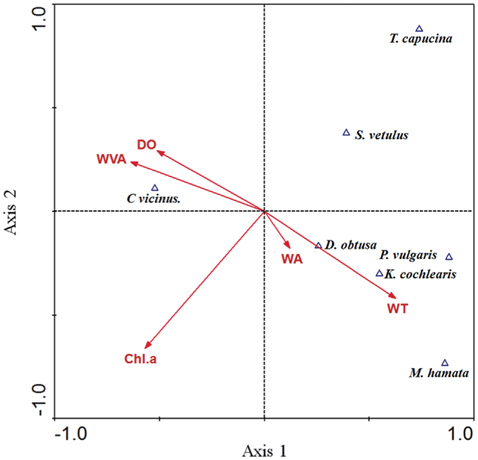
The results of CCA showed the relationship between environmental factors and zooplankton species (Fig. 2). The eigenvalues for CCA axis 1 and axis 2 were 0.73 and 0.46, respectively, and they accounted for 54.2% of the cumulative variance in the species data. The species–environment correlations for CCA axes 1 and 2 were high, and accounted for 68.2% of the variance in the species composition–environment relationship. Specifically, the withered vegetated area and DO had a strong effect on C. vicinus, Monostyla hamata, Keratella cochlearis, Polyarthra vulgaris, and Daphnia obtusa densities and compositions, which were associated with both the reservoir area and water temperature.
In this study, we found high densities of C. vicinus in shallow reservoirs during winter. Copepods are commonly abundant under high water temperature conditions from spring to autumn (Huq et al. 1984), and winter typically constitutes low densities. Low water temperature leads to decreased metabolic rates in copepods, and a decrease in their population growth. However, some studies have reported the presence and density pattern of copepods during winter (Hoffmeyer 1994, Dejen et al. 2004). Copepods present during winter are believed to be responsible for sustaining copepod abundance at certain levels in the following autumn seasons. Moreover, copepod abundance in winter can largely contribute to population growth in spring. Importantly, the increased number of copepods in spring can be utilized as food items for the growth of secondary consumers, such as juvenile fish and macroinvertebrates.
From results of the CCA analysis, we found that copepod density was related to withered vegetation area. Even though area of withered vegetation does not always be proportional to the occupied space by the vegetation, a strong positive relationship between C. vicinus and withered vegetation area was found (r2 = 0.76, P < 0.01), implying that the height of withered vegetation had relatively constant influence. Strong association of C. vicinus with withered vegetation area in CCA result would be explained by the circumstance. Generally, aquatic vegetation is utilized as a refuge to avoid predators such as fish, and thus can help to increase the density of zooplankton, include copepods (Manatunge et al. 2000, Kuczyńska- Kippen and Nagengast 2006). Although the foraging activity of predators is reduced in low water temperatures (Jang et al. 2006, Carey et al. 2010, Theel and Dibble 2008), some predators can consume prey species under such conditions, and in such a case, withered vegetation can be utilized as a habitat for copepods. Therefore, the presence of withered vegetation might be considered an important factor in determining C. vicinus distribution during winter. Furthermore, macrophytes, through nutrient competition and allelopathy with phytoplankton (Van Donk and Van de Bund 2002), can decrease algal food for zooplankton, but withered vegetation may exert a limited effect on the phytoplankton. Although chlorophyll a did not show a close relationship to copepod density in the present study, the possibility cannot be entirely excluded. This is one of reasons why withered vegetation could support a high density of copepods. The withered vegetation is not influenced by periphytic algae or phytoplankton; moreover, the leaves and stem surfaces are used as a habitat for periphytic algae. Based on this information, our results should indicate that the high concentrations of dissolved oxygen seen are due to the high amount of periphytic algae present.
Non-copepod zooplankton species were partly analyzed by environmental parameters in the CCA. The presence of Daphnia obtusa, Polyarthra vulgaris, Keratella cochlearis, and Monostyla hamata was related to both reservoir area and water temperature. However, the occurrence of other zooplankton species at the 25 study sites was relatively low (Table 2), indicating that the aforementioned relationship should be considered cautiously.
In the present study, we hypothesized two possibilities for copepod (i.e., C. vicinus) distribution under low water temperatures: (1) predator foraging is strongly dependent on water temperature (Byström et al. 2006), with copepods showing high densities, because the foraging activity of predators deceases under low water temperatures and (2) the resting eggs of copepods might exhibit hatching selection based on lower water temperatures. Copepods are always exposed to predation under higher water temperatures (Turner 1984), and to avoid predators, their hatching mechanisms might show a strong possibility for adaptation to low water temperatures. We found abundant egg-shells of copepod resting eggs in several reservoirs during the study period (sites 2, 12, and 16) which allows us to suspect the possibility, even though we cannot assure the hatching timing of the resting eggs.
We expect that withered vegetation in water body supports tolerance of Cyclops population to unfavorable growth condition (i.e., low temperature in winter), leading to persistence of population size at a certain level. In this study, we observed very low abundance of Cyclops in the area where withered vegetation was absent. On the contrary, their abundance was larger as withered vegetation was abundant. Although copepods prefer high water temperatures for increasing their density and population growth, if Cyclops can overcome low temperature stress that leads to disruption of population, their population growth initiation in the next growing season (i.e., next spring) is possibly propelled by the winter population. Also, the tolerance to winter season may advantage Cyclops population to avoid extensive predation impact in summer. Typically their predators have increased foraging activity under high water temperature, and possible temperature tolerance coupled with withered vegetation may allow to avoid summer predation. This hypothetical advantage in compliance with mechanism of Cyclops persistence in withered vegetation area should be elucidated in further in-depth approach.
Cyclops vicinus was dominant during winter, and this is significantly related to withered vegetation area and dissolved oxygen among diverse environmental factors. Therefore, the presence of withered vegetation might be considered as an important factor in determining C. vicinus distribution during winter. The withered vegetation in water body supports tolerance of Cyclops population to unfavorable growth condition (i.e., low temperature in winter), leading to persistence of population size at a certain level. Although copepods prefer high water temperatures for increasing their density and population growth, if Cyclops can overcome low temperature stress that leads to disruption of population, their population growth initiation in the next growing season (i.e., next spring) is possibly propelled by the winter population. Moreover, the tolerance to winter season may advantage Cyclops population to avoid extensive predation impact in summer.