Traditional Chinese medicine (TCM) has become an increasingly popular alternative for the treatment of a variety of physiological disorders [1]. Shengmaisan (SMS), one of the most famous Chinese herbal formula, consists of
Interstitial cells of Cajal (ICCs) play an important role in the generation of coordinated GI peristalsis [5-8]. The major function of the ICCs is the generation of slow waves that allow these cells to act as specialized pacemaker cells [6-8], and they have additional functions in the gastrointestinal tract as regulators of mechanical activity and neurotransmission [7-9]. Due to the central role of ICCs in GI motility, it has been suggested that defects of ICCs may contribute to motility dysfunction in several GI motility disorders [10]. Therefore, experimental observations of the ICCs should be a good tool to investigate the GI motility functions. However, the effects of SMS on small intestinal ICCs in a mouse have not yet been investigated. Therefore, we undertook this research to investigate the effects of SMS in mouse small intestinal ICCs.
2.1. Preparation of cells and cell cultures
Balb/c mice (3-7 days old) of either sex were anesthetized with ether and sacrificed by cervical dislocation. All animal procedures were approved by the Institutional Animal Care and Use Committee of Pusan National University (Busan, South Korea) and carried out in accordance with the Guide for the Care and Use of Laboratory Animals of Pusan National University. All efforts were undertaken according to the “3R principles” of the directive to reduce the number of animals used in this study and to optimize the experimental protocols to acquire the maximum amount of data from each tested animal. The small intestines from 1 cm below the pyloric ring to the cecum were removed and opened along the mesenteric border. Luminal contents were removed by washing with Krebs-Ringer bicarbonate solution. The tissues were pinned to the base of a Sylgard dish and the mucosa were removed by sharp dissection. Small tissue strips of the intestine muscle (consisting of both circular and longitudinal muscles) were equilibrated in Ca2+-free Hanks solution (containing in mmol/L: KCl 5.36, NaCl 125, NaOH 0.34, Na2HCO3 0.44, glucose 10, sucrose 2.9, and HEPES 11) for 30 min. Then, the cells were dispersed using an enzyme solution containing collagenase (Worthington Biochemical Co., Lakewood, NJ, USA) 1.3 mg/mL, bovine serum albumin (Sigma Chemical Co., St. Louis, MO, USA) 2 mg/mL, trypsin inhibitor (Sigma) 2 mg/mL and ATP 0.27 mg/mL. Cells were plated onto sterile glass coverslips coated with murine collagen (2.5 μg/mL, Falcon/BD, Franklin Lakes, NJ,USA) in a 35-mm culture dish and then cultured at 37 °C in a 95% O2, 50 mL/L CO2 incubator in a smooth muscle growth medium (Clonetics Corp., San Diego, CA, USA) supplemented with 2% antibiotics/antimycotics (Gibco,Grand Island, NY, USA) and murine stem cell factor (SCF, 5 ng/mL, Sigma). ICCs were identified immunologically with anti-c-kit antibody (phycoerythrin-conjugated rat anti-mouse c-kit monoclonal antibody; eBioscience, San Diego,CA, USA) at a dilution of 1:50 for 20 min. ICCs were morphologically distinct from other cell types in the culture; thus, it was possible to identify the cells by using phase contrast microscopy once they had been verified with the anti c-kit antibody.
The whole-cell patch-clamp configuration was used to record membrane potentials (current clamp) from cultured ICCs. An axopatch ID (Axon Instruments, Foster, CA, USA) was used to amplify membrane currents and potentials. The command pulse was applied using an IBM-compatible personal computer and pClamp software (version 6.1; Axon Instruments). Data obtained were filtered at 5 kHz and displayed on an oscilloscope, a computer monitor, and a pen recorder (Gould 2200, Gould, Valley View, OH, USA). Results were analyzed using pClamp and Origin (version 6.0) software. All experiments were performed at 30-32℃.
The physiological salt solution used to bathe cells (Na+-Tyrode) contained (mmol/L): KCl 5, NaCl 135, CaCl2 2, glucose 10, MgCl2 1.2 and 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) 10, adjusted to pH 7.4 with NaOH. The pipette solution contained (mmol/L): KCl 140, MgCl2 5, K2 Adenosine triphosphate (ATP) 2.7, Na Guanosine triphosphate (GTP) 0.1, creatine phosphate disodium 2.5, HEPES 5 and ethylene glycol tetraacetic acid (EGTA) 0.1, adjusted to pH 7.2 with KOH.
SMS was purchased from I-WORLD Pharmaceuticals (South Korea). The dosage for an adult human is 10-15 g (crude materials) per day. More information about SMS can be found on I-WORLD Pharmaceuticals Homepage (http://i-pharm.koreasme.com). The SMS was dissolved in distilled water at a concentration of 0.5 g (crude drug)/ml and stored in a refrigerator. SMS is a powder preparation and contains lactose and corn starch as an excipient. The lactose and corn starch can affect the ICCs and the GI motility. However, because SMS containing lactose and corn starch is eaten, we think that any investigation of its effects should use SMS containing that excipient.
All other drugs were obtained from Sigma (Sigma Chemical Co., USA). The drugs were dissolved in distilled water, and just prior to use were added to the bath solution to make the desired concentrations. Addition of these chemicals to the bath solution did not alter the pH of the solution. Thapsigargin, U-73122, U-73343, chelerythrine and calphostin C were dissolved in dimethyl sulfoxide (DMSO) for the 50 or 100 mmol/L stock solution and were added (1000 times dilution) to the bathing solution on the day of the experiment. The final concentration of DMSO in the bath solution was always < 0.1%, and we confirmed that this concentration of DMSO did not affect the results that were recorded.
All data are expressed as means ± S.E.’s. The student’s
3.1. Effect of SMS on pacemaker potentials in cultured ICCs
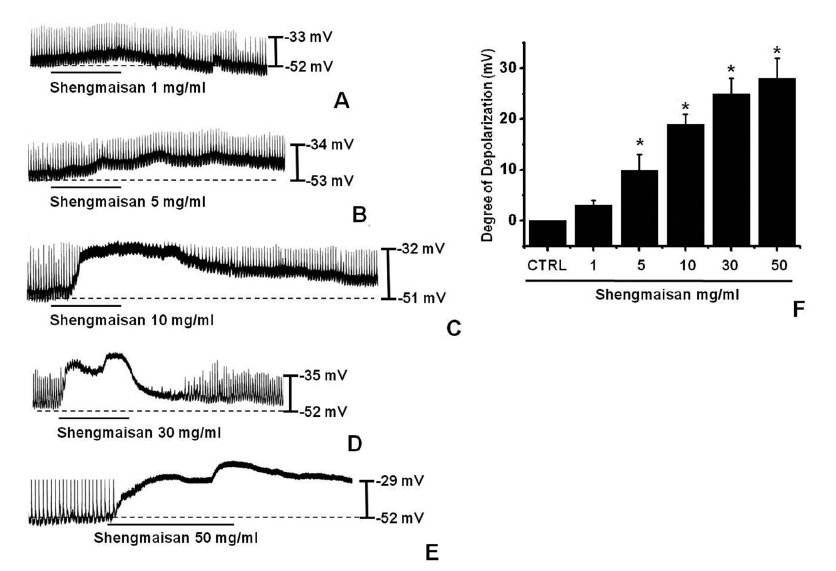
The patch-clamp technique was used to investigate the ICCs that formed network-like structures in culture (2-4 days). Spontaneous rhythms were routinely recorded from cultured ICCs under current and voltage-clamp conditions, and the ICCs within the networks displayed more robust electrical rhythms. Tissue-like spontaneous slow waves have been recorded from these cells [11]. To understand the relationship between SMS and the modulation of pacemaker activity in ICCs, we examined the effects of SMS on pacemaker potentials. Recordings from cultured ICCs under current clamp conditions (I = 0) showed spontaneous pacemaker potentials. The resting membrane potential was -52.1 ± 2.1 mV and the amplitude was 20 ± 4 mV. In the presence of SMS (1 - 50 mg/ml), the membrane potentials were depolarized to 3.0 ± 1.1 mV at 1 mg/ ml, 11.2 ± 3.2 mV at 5 mg/ml, 19.4 ± 2.4 mV at 10 mg/ml, 25.2 ± 3.2 mV at 30 mg/ml and 28.1 ± 4.1 mV at 50 mg/ml (Fig 1)A-1E). The summarized values and the bar graph of SMS’s effects on pacemaker potentials are indicated in Fig. 1F (n = 4).
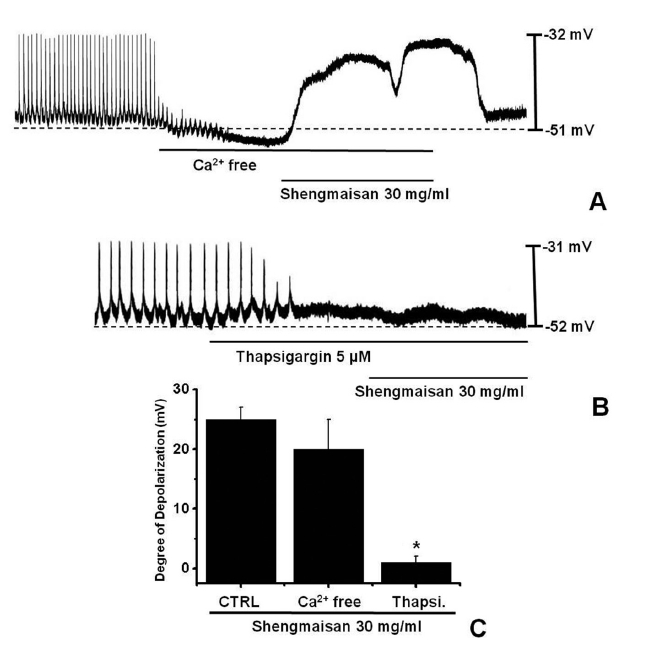
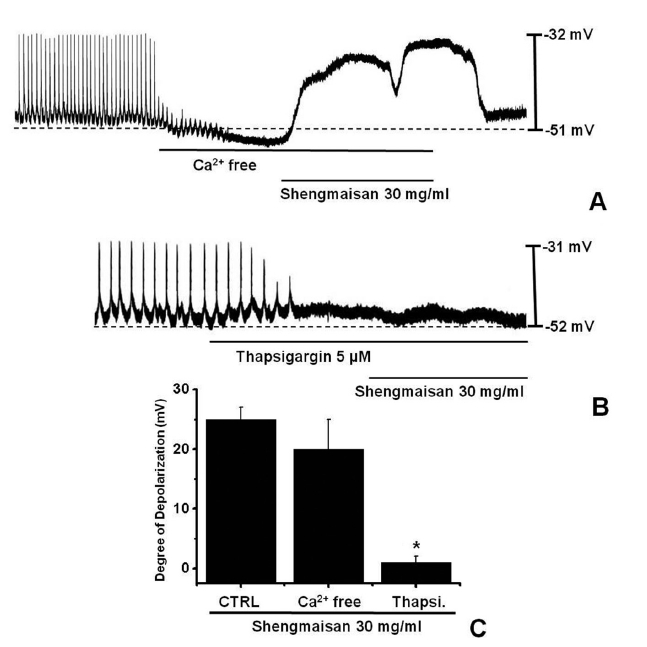
The external Ca2+ influx is necessary for GI smooth muscle contractions and is essential for generating pacemaker potentials in the ICCs. The generation of pacemaker currents has been shown to depend on the intracellular Ca2+ oscillation [12]. To investigate the role of external Ca2+ or internal Ca2+, we tested SMS under external Ca2+-free conditions and in the presence of thapsigargin, a Ca2+-ATPase inhibitor of the endoplasmic reticulum [11, 13]. The pacemaker potentials completely disappeared when an external Ca2+-free solution was used. In this condition, SMS induced membrane depolarizations (n = 5; (Fig 2)A). Under external Ca2+-free conditions, the membrane depolarizations with SMS (30 mg/ml) were not significantly different from those with SMS (30 mg/ml) in normal Ca2+ solution (n = 5, (Fig 2)C). In addition, SMS-induced membrane depolarizations were inhibited by pretreatment with thapsigargin (n = 4, (Fig 2)B). In the presence of thapsigargin (5 μM), the membrane depolarizations with SMS were significantly different from those with SMS in the absence of thapsigargin (n = 4, (Fig 2)C).
3.3. Effects of a phospholipase C inhibitor on SMS-induced
pacemaker potentials in cultured ICCs
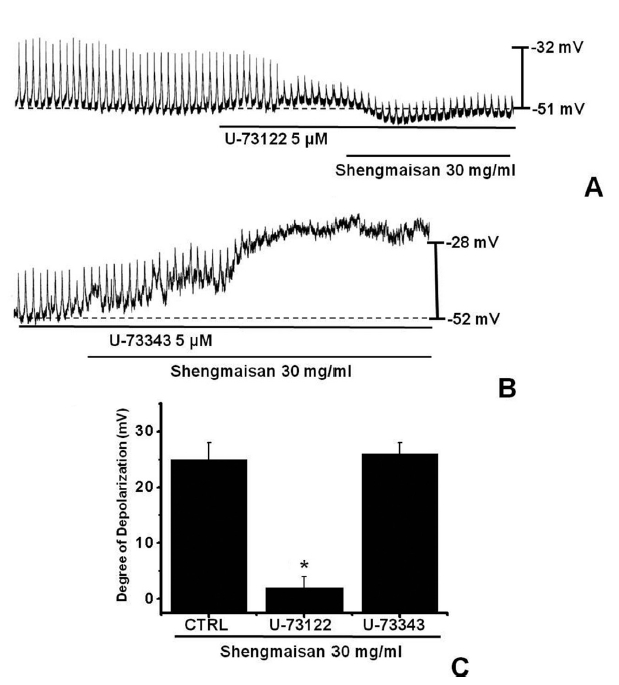
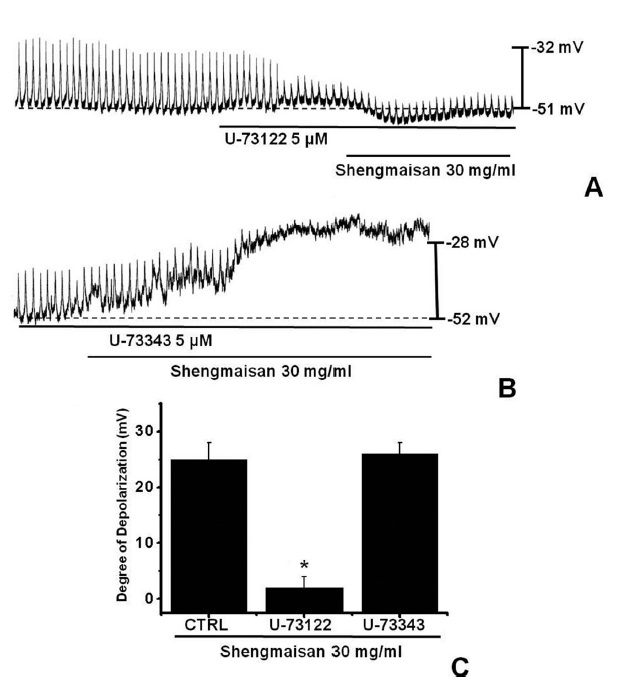
Because the membrane depolarizations cause by SMS were related to intracellular Ca2+ mobilization, we examined whether its effects on pacemaker potentials required phospholipase C (PLC) activation. To test this possibility, we measured SMS-induced membrane depolarizations in the absence and the presence of U-73122, an active PLC inhibitor [14]. The pacemaker membrane depolarizations completely disappeared when U-73122 (5 μM) was used. Under these conditions, SMS (30 mg/ml)-induced membrane depolarizations were not produced (n = 4; (Fig 3)A). In the presence of U-73122, the membrane depolarization produced by SMS was 2.3 ± 2.1 mV. The value of membrane depolarization caused by SMS in the presence of U-73122 was significantly different from that caused by SMS in the absence of U-73122 (n = 4, (Fig 3)C). The treatment with U-73343 (5 μM), an inactive analog of U-73122, had no influence on the SMS-induced pacemaker potentials. Under these conditions, SMS (30 mg/ml)-induced membrane depolarizations were not suppressed by using U-73343 (n = 4, (Fig 3)B).
3.4. Effect of protein kinase C inhibitor on SMS-induced
pacemaker potentials in cultured ICCs
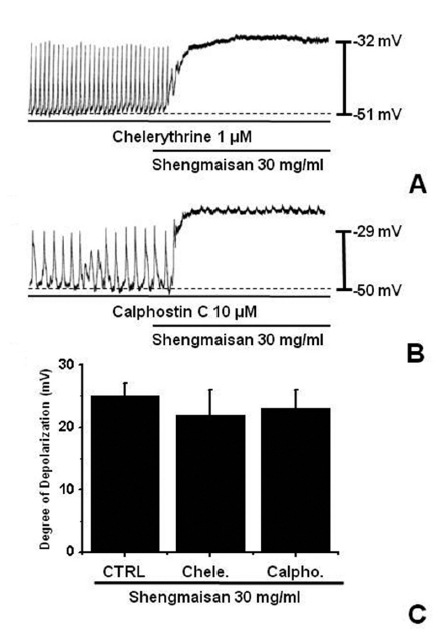
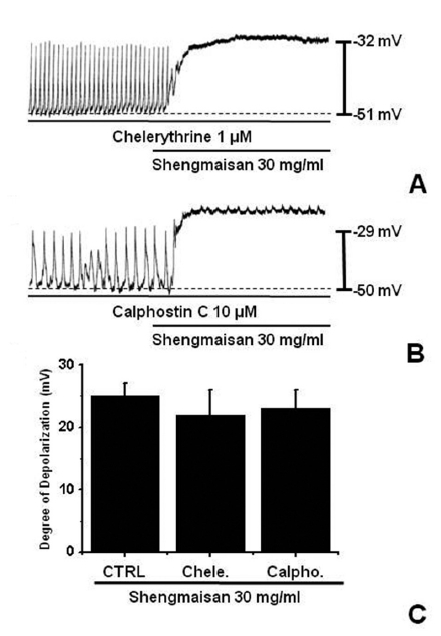
We tested the effects of chelerythrine or calphostin C, an inhibitor of protein kinase C (PKC) [13, 15], to investigate whether the SMS-induced responses of the pacemaker potentials were mediated by the activation of PKC. Chelerythrine (1 μM) or calphostin C (10 μM), did not have any effect on the membrane depolarizations caused by SMS (30 mg/ml; (Fig 4) and the value was not significantly different from the membrane depolarization caused by SMS in the absence of chelerythrine or calphostin C (n = 4; (Fig 4)C).
SMS is a traditional Chinese herbal medicine that has long been used for the treatment of heart failure and ischemic heart disease. It is a mixture of three herbal components. Constituent herbs of SMS include
ICCs are the pacemaking cells in the GI muscles that generate the rhythmic oscillations in the membrane potential known as slow waves [5-8]. c-kit is a transmembrane protein kinase receptor that is essential for the development and function of ICCs. In the gut, c-kit is expressed only in ICCs and mast cells [5, 8]. ICCs are distributed throughout the digestive tract from the upper oesophageal sphincter to the internal sphincter of the anus [22-24]. ICCs contribute to important functions in the digestive tract due to their generation, coordination and propagation of electrical slow wave activity, their transduction of motor neural inputs from the enteric nervous system and their role in the mechanosensation of smooth muscle cells [8].
In this study, SMS produced membrane depolarization in ICCs. Pretreatment with a Ca2+-free solution and a thapsigargin, a Ca2+-ATPase inhibitor in the endoplasmic reticulum, stopped the generation of pacemaker potentials. Under Ca2+-free solution conditions, SMS showed membrane depolarizations. However, under thapsigargin solution conditions, SMS had no effects on membrane depolarizations. Also, the pacemaker membrane depolarizations were inhibited by U-73122, an active PLC inhibitor. Furthermore, the PKC inhibitors, chelerythrine and calphostin C, did not block the SMS-induced membrane depolarizations. These results suggest that SMS might affect GI motility by the modulation of pacemaker activity in the ICCs, and that the activation takes place in an internal Ca2+ and PLC-dependent and an external Ca2+ and PKC-independent manners.
Our data suggest that the gastroprokinetic effects of SMS might be mediated by the induction of pacemaker potentials in the ICCs. When the effects of this drug on the ICCs are considered, further research, including finding active compound(s) and examining their action mechanisms, is clearly needed.