In general, the deep sea is referred to as the watershed below 200 m at the end point of the continental shelf. The average depth of East Sea in Korea is about 1,700 m (Sohn et al., 2010). Deep sea water (beginning at a depth of 200 m) has stable environmental characteristics, including a low temperature, high salinity, and an absence of penetration by natural light. The species diversity and number of inhabitants in the East Sea have been shown to be very low (Barnes and Mann, 1991; Park et al., 2007).
Several recent studies have raised awareness of the need to develop the marine resources that inhabit the deep sea. Coastal marine resources have become depleted as the number of fish catches has increased worldwide, and overfishing, promoted by technological developments in fishing boats and gear, is threatening future supplies.
Species known to inhabite the eastern deep sea in Korea include the snow crab
yama
The fishery characteristics, population variation, ecological characteristics, and geographical distribution of the aforementioned species have been studies (Kim et al., 2007).
However, around 50% of deep sea animals abandoned on fishing vessels because of a lack of consumer demand, likely because of their appearance and unfamiliar name. Thus, it is necessary to develop effective processing technologies and to ascertain the biochemical composition of the animals that inhabit the deep sea in order to expand the utilization of these resources as food (Park et al., 2007).
In this study, we examined the biochemical composition of muscle from Tanaka’s eelpout
Tanaka’s eelpout
>
Proximate composition analysis
The moisture content of the fishes was determined by drying the meat in an oven at 105℃ until a constant weight was obtained (Association of Official Analytical Chemists, 1990). The crude protein content was calculated by converting the nitrogen content, determined by Kjeldahl’s method (6.25 × N) (Association of Official Analytical Chemists, 1990). The crude lipid content was determined described by the Association of Official Analytical Chemists (1990) using the Soxhlet system. The ash content was determined by dry ashing in a furnace oven at 525℃ for 24 h. The carbohydrate content was determined as 100 minus the moisture, crude protein, crude lipid, and ash contents.
The total amino acid composition was determined using an amino acid analyzer (S43000; Sykam, Eresing, Germany). All samples were hydrolyzed in 6 N HCl in vacuum-sealed tubes at 110℃ for 24 h.
Fifteen grams each of animals was placed in a Corning tube (Corning, NY, USA) containing 30 mL of extraction solvent (100% ethanol) and blended for 1 h. The extract was then transferred to a 100 mL volumetric flask and the tube was rinsed once with 30 mL of 70% ethanol then blended for 2 min using a sonicator (D78224; Singen, Elma, Germany). The filtrate was enriched using a rotary evaporator (N-1000; Eyela, Tokyo, Japan) and combined with 25 mL of the lithium citrate loading buffer (pH 2.2). The sample was passed through an Advantec cellulose acetate 0.20 μm filter membrane and the free amino acid composition was determined using an amino acid analyzer (S43000; Sykam).
Total lipids were extracted according to the method of Folch et al. (1957), using chloroform/methanol (2:1). Aliquots
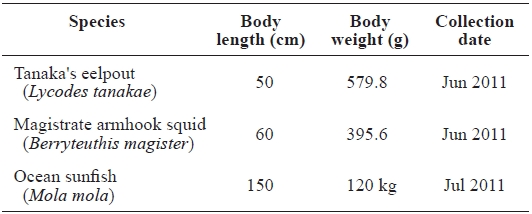
[Table 1.] The profiles of samples caught in the East Sea, Korea
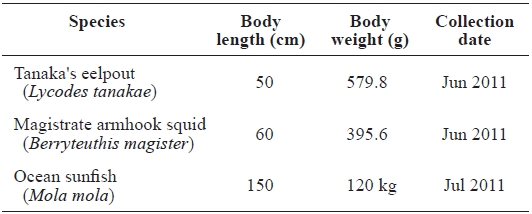
The profiles of samples caught in the East Sea, Korea
of the chloroform layer extract were evaporated to dryness under nitrogen and lipids were quantified gravimetrically.
Fatty acid methyl esters (FAMEs) were obtained as described by the American Oil Chemists’ Society (1997). A fraction of the lipid extract was saponified with 0.5 N NaOH in methanol followed by methylation in 14% boron trifluoride in methanol. The resulting methyl esters were analyzed using GC-17A gas chromatograph (Shimazu, Tokyo, Japan) equipped with a flame ionization detector, a split ratio (1:50) injector, and an Omegawax-320 capillary column (film thickness: 30mⅹ0.32 mm; i.dⅹ 0.25 μm). The temperatures of the injector and detector were 250 and 260℃, respectively. Helium was used as the carrier gas (flow rate, 1.03 mL/min). Peaks were identified by comparison of their retention times with a 37 component FAMEs mixed standard (Supelco SA, Gland, Switzerland).
The data were analyzed using analysis of variance through the general linear model procedure (SAS Institute, Cary, NC, USA). Duncan’s multiple-range test was applied to determine the significance of differences between means (
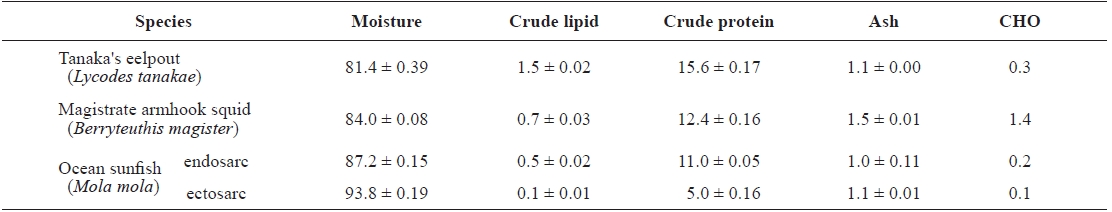
The proximate compositions of Tanaka’s eelpout, magistrate armhook squid and ocean sunfish are shown in Table 2.
The moisture contents were 81.4 ± 0.39, 84.0 ± 0.08, 87.2 ± 0.15 and 93.8 ± 0.19 g/100 g for Tanaka’s eelpout, magistrate armhook squid, ocean sunfish’s endosarc and sunfish’s ectosarc, respectively. The crude lipid contents were 1.5 ± 0.02, 0.7 ± 0.03, 0.5 ± 0.02 and 0.1 ± 0.01 g/100 g. The crude protein contents were 15.6 ± 0.17, 12.4 ± 0.16, 11.0 ± 0.05 and 5.0 ± 0.16 g/100 g. The ash contents were 1.1 ± 0.00, 1.5 ± 0.01, 1.0 ± 0.11 and 1.1 ± 0.01 g/100 g. The carbohydrate contents were 0.3, 1.4, 0.2 and 0.1 g/100 g. Finally, the energy contents
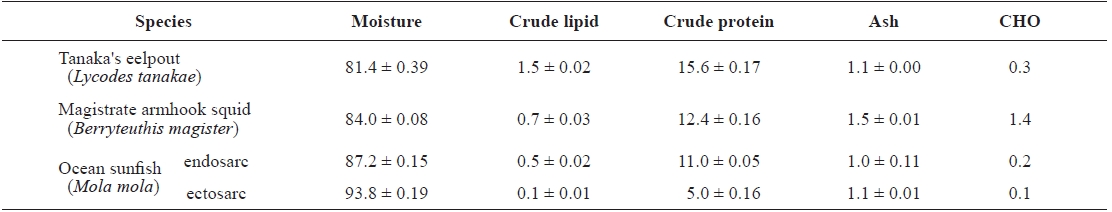
Proximate composition (g/100 g) of muscle from Tanaka's eelpout, magistrate armhook squid, and ocean sunfish
of the animals were 77.1, 61.5, 49.3 and 21.3 kcal, respectively; the highest content was in Tanaka’s eelpout whilelowest was in sunfish’s ectosarc.
These results show a clear inverse relationship between the lipid and moisture contents, with high lipid values being matched by a low moisture content and vice versa (Hernandez et al., 2003). The lipid stored in muscle is largely neutral lipids in the form of oil droplets (Shindo et al., 1986). These oil droplets were replaced by interstitial water; thus, the moisture content in the muscle was reduced while the crude protein content remained unchanged. An inverse relationship between these values has been shown (Ackman, 1989; Mok et al., 2007).
The reported average moisture content of 51 species of Korean coastal water fishes, including Japanese anchovy
The levels of moisture (88.2-92.5%), crude protein (7.0- 9.9%), crude lipid (0-0.1%), ash (0.9-1.1%), and carbohydrate (0-1.2%) in cubed snailfish
With regard to demersal fish, most settlement fish, which exhibit low mobility owing to the face that they do not need to expend much energy were shown to have low lipid contents (Jeong et al., 1998a).
Deep sea fishes live longer than organisms on the continental shelf, and have a low growth rate (Park and Bae, 2011). The average moisture content in Tanaka’s eelpout and sunfish was >80 g/100 g which is higher than that in coastal water pathfishes. The average crude protein content in these species was <15.6 g/100 g while the average crude lipid content was <1.5 g/100 g, which is less than the values reported for coastal water fishes.
The levels of moisture (77.3-88.1 g/100 g), crude protein (7.7-18.3 g/100 g), crude lipid (<0.1-1.4 g/100 g), ash (1.3-2.5 g/100 g) and carbohydrate (<0.1-2.0 g/100 g) in six species of cephalopods found in the coastal waters, including cuttlefish
The water layer inhabited by squids is approximately 200 m from the surface. The water layer inhabited by the Japanese hooked squid
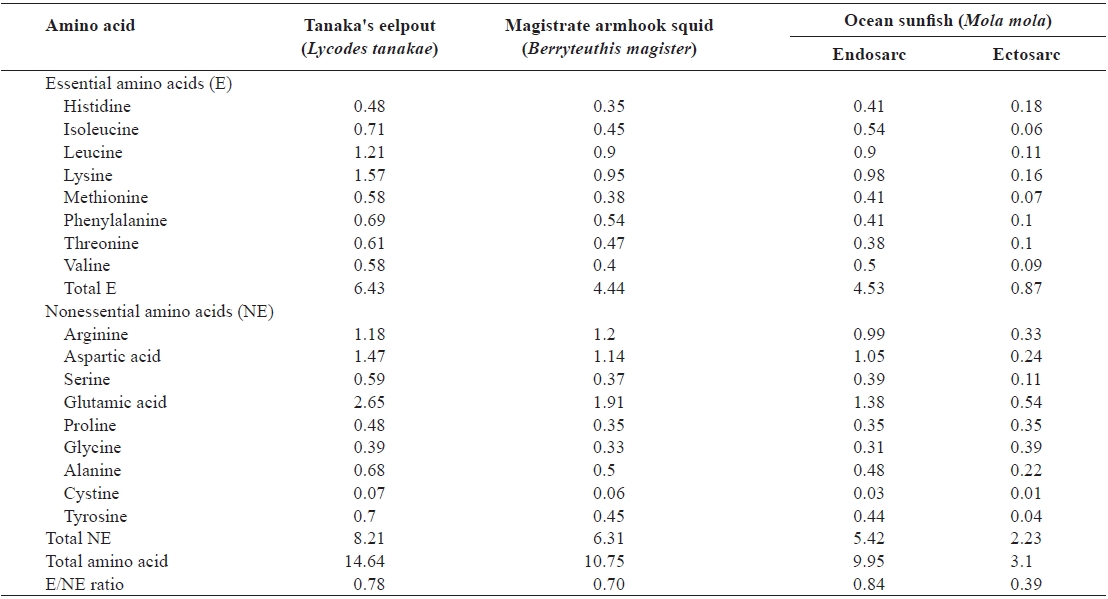
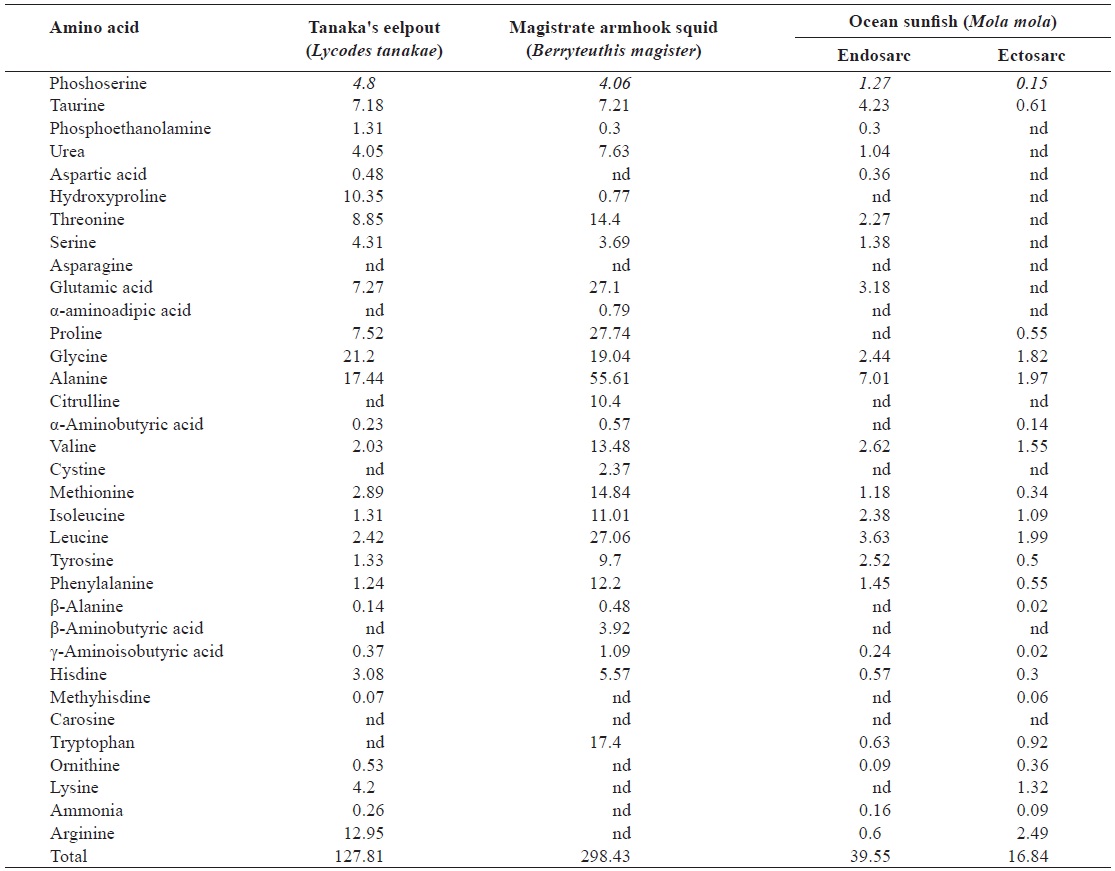
The total amino acid contents of these deep-sea animals are shown in Table 3. Seventeen amino acids other than tryptophan were detected in the samples; tryptophan was damaged by hydrolysis using HCl.
The total amino acid contents of Tanaka’s eelpout, magis-
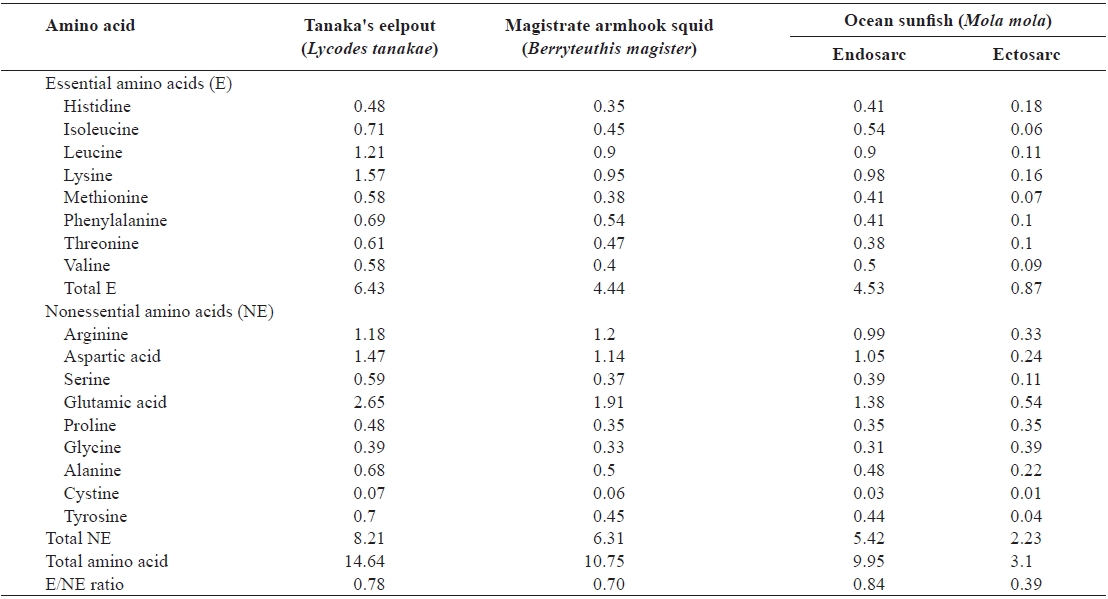
The total amino acid content (g/100g) of muscle from Tanaka's eelpout, magistrate armhook squid, and ocean sunfish
trate armhook squid, and ocean sunfish endosarc, and ectosarc were 14.64, 10.75, 9.95 and 3.10 g/100 g, respectively. Tanaka’s eelpout and ocean sunfish’s endosarc contained significantly higher amounts of total amino acids than magistrate armhook squid and ocean sunfish’s ectosarc. High contents of glutamic acid, aspartic acid, lysine, leucine and arginine were detected in these animals, while those of the histidine, glycine and cysteins were low. The animals were also found to be rich in glutamic acid (2.65, 1.91, and 0.54-1.38 g/100 g, respectively, Tanaka’s eelpout, magistrate armhook squid, and ocean sunfish). The glutamic acid contents of coastal sea fishes were analyzed and previously found to be similar to or greater than those in the present samples (1.71-1.83, 1.60-1.77 and 1.91-2.18 g/100g, respectively, in red seabream
The essential amino acids ratio in Tanaka’s eelpout (44%), magistrate armhook squid (41%), and ocean sunfish (28-46%) was also calculated. The essential amino acids/nonessential amino acids ratios in Tanaka’s eelpout, magistrate armhook squid, ocean sunfish’s endosarc, and ectosarc were 0.78, 0.70, 0.84 and 0.39%, respectively. The E/NE ratios of magistrate armhook squid and ocean sunfish’s ectosarc were lower than the common level (0.74%) in most of fishery products (Iwasaki and Harada, 1985; Kang et al., 2009). Sulfur, mainly in the form of sulfur-containing amino acids such as taurine, methionine and cysteine are essential for the detoxification path-ways in the liver. This is one of the major roles of sulfur in the body (Kang et al., 2007). The values of sulfur-containing amino acids in Tanaka’s eelpout, magistrate armhook squid, and ocean sunfish were 0.65, 0.44 and 0.08-0.44 g/100 g. Aromatic amino acids such as phenylalanine and tyrosine function as substance as neurotransmitters in the brain (Kang et al., 2007). The amounts of aromatic amino acids in Tanaka’s eelpout (1.39 g/100 g), magistrate armhook squid (0.99 g/100 g) and ocean sunfish (0.14-0.85 g/100 g) were analyzed. There was very a little appreciable difference in the total amino acid composition of aquatic animals, depending on the species (Lee et al., 2000).
Free amino acids, which are a major component of active biological substances, have been shown to have distinct flavors and have been classified accordingly (Ohta, 1976). For example, glycine, alanine, threonine, proline and serine are described as sweet tasting while leucine, isoleucine, methio-
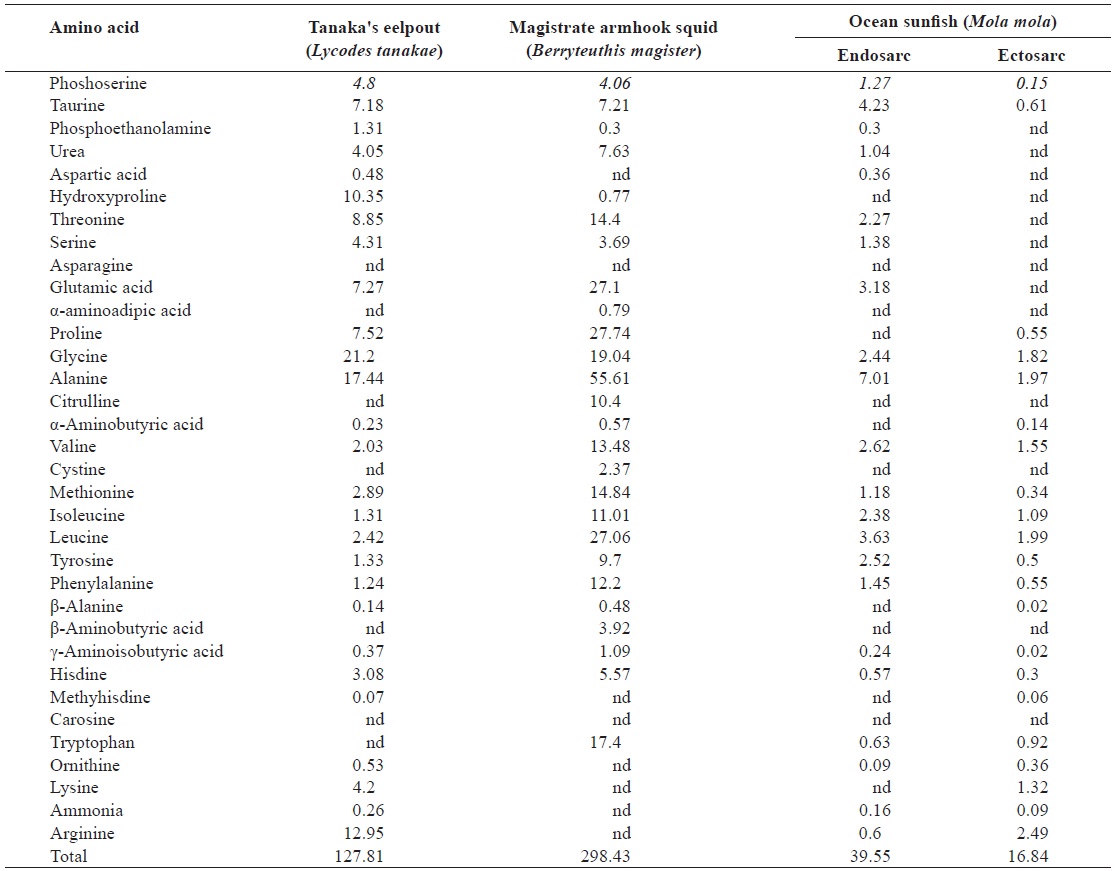
The free amino acid content (mg/100 g) of muscle from Tanaka's eelpout, magistrate armhook squid, and ocean ocean sunfish
nine, phenylalanine, lysine, valine and arginine are bitter tasting amino acids. Moreover, aspartic acid has a sour taste while glutamic acid is considered umami taste (Shou, 1969). The free amino acid contents of our samples are shown in Table 4. The total free amino acid values for Tanaka’s eelpout, magistrate armhook squid, and ocean sunfish were 127.81, 298.43 and 16.84-39.55 mg/100 g, respectively. The amounts of free amino acids in red seabream, brassblotched rockfish and olive flounder were previously shown to be 563.39-601.57, 451.89-530.30, and 300.18-473.75 mg/100 g, respectively, with taurine as the dominant amino acid (Kim et al., 2000). The free amino acid content in deep-sea aquatic animals was 16.84-298.43 mg/100 g and the taurine content was 0.61-7.21 mg/100 g. These values are lower than those of coastal organisms. Taurine represented 39-65% of the free amino acid content, followed in abundance by hydroxyproline, lysine, alanine, and glycine (Kim et al., 2000). The prominent amino acids in Tanaka’s eelpout were glycine, alanine, and arginine.
Magistrate armhook squid had high contents of proline, glycine, alanine and leucine whereas ocean sunfish contained only minor amounts of free amino acids. In general, the alanine and glycine contents were highest in deep-sea aquatic animals and coastal organisms; however, there was no significant similarity between them.
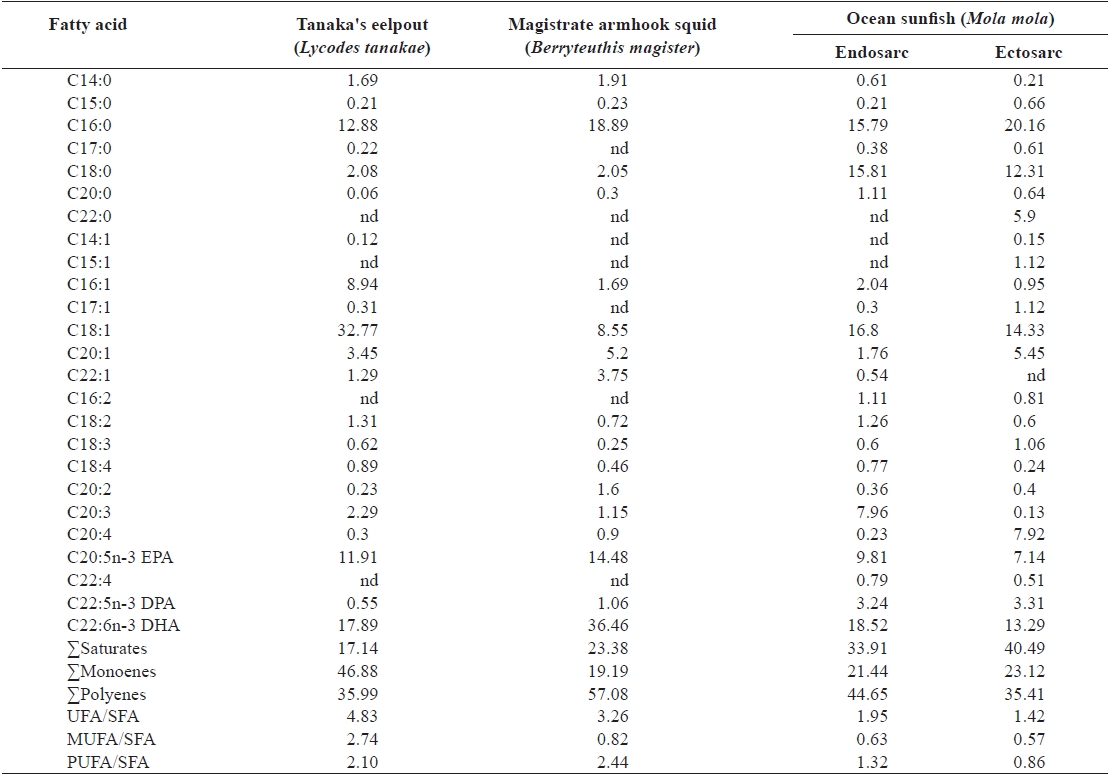
The total fatty acid compositions of the deep-sea animals are shown in Table 5. The prominent saturated fatty acids (SFAs) were C16:0 in Tanaka’s eelpout and magistrate armhook squid and C18:0 in ocean sunfish. The content of monounsaturated fatty acids such as palmitoleic acid (C16:1) and oleic acid (18:1) were not different in these samples except for Tanaka’s eelpout.
The prominent fatty acids in coastal fishes are C16:0,
The fatty acid composition (area%) of muscle from Tanaka's eelpout, magistrate armhook squid, and ocean sunfish
C20:0, C22:6, C18:1, C20:5, and C16:1 (Kang et al., 2007). Similar to coastal fishes, the major fatty acids in deep-sea aquatic animals were C16:0, C18:0, C16:1, C18:1, C20:5, and C22:6. In contrast, Tanaka’s eelpout had a high C18:1 content while C22:6 was the most abundant fatty acid in magistrate armhook squid and ocean sunfish.
The fatty acid contents in five species of migratory fish (Japanese anchovy
The major unsaturated fatty acids in twelve species of coastal fishes were previously shown to be DHA, EPA, C20:4n-6, and DPA (C22:5n-3) (Jeong et al., 1998b). The DHA content (7.26-28.4%) was highest in black scraper and lowest in flathed grey mullet. The EPA content (4.21-17.9%) was highest in ridge-eye flounder and lowest in Japanese amberjack (Jeong et al., 1999).
Flathead grey mullet, dotted gizzard shad, and ridge-eye flounder had high levels EPA while the other species had abundant DHA. Black scraper possessed >5% C20:4n-6, while flathead grey mullet, blackhead seabream, and ridgeeye flounder possessed <5% DPA (Jeong et al., 1999). All of the deep-sea animal samples had circa twice as much DHA as EPA. Moreover, these samples contained trace DPA. The total fatty acid compositions of the deep-sea animals were similar to those of coastal organisms.