Sacoglossan mollusks are suctorial herbivores that live on the cell sap of coenocytic macroalgae (Jensen 1993).As an adaptation for suctorial feeding, these sea slugs have uniserial radula and show narrow specificity for their algal diet (Handeler and Wagele 2007). With few exceptions,algae that have been identified as food for sacoglossan sea slugs belong to the class Ulvophyceae sensu Floyd and O’Kelly (1990). However, food organisms are not known for many sacoglossans yet.
Some sacoglossan mollusks incorporate algal chloroplasts inside their cells, and the incorporated chloroplasts are functional for days to months, depending on the species. Trench et al. (1972) provided conclusive evidence that photosynthetic assimilates are released from the chloroplasts in the sea slug cells. This incorporation and maintenance of foreign chloroplasts is known as kleptoplasty (e.g., Clark et al. 1990). It has long been suspected that genes have been transferred from algae to the sea slugs, because many components of photosystems in active algal plastids are very short-lived, with turnover times especially under bright light condition (Schuster et al. 1988, Vass et al. 1992, Warner et al. 1999). Thus, if the plastids are maintained in an active state for weeks and months, it is logical to assume that the mollusks could produce necessary proteins to keep the photosynthetic activity because the photosynthetic proteins in chloroplasts are turned over quickly (often in less than a day). Recently, accumulating evidences show that algal nuclear genes have been laterally transferred to these sacoglossan species (Rumpho et al. 2001, 2008, Pierce et al. 2007, Schwartz et al. 2010).
Sacoglossan mollusks bind avidly to their food algae. In most cases the sacoglossan mollusks eat less than ten species of algae throughout their life cycle, and the chloroplasts from only one or two species could be sequestered and maintained in the sea slug cells. This feeding specificity is also necessary for the development of successful lateral gene transfer which might render the acquired chloroplast photosynthetically active for long time.
Here, we screened for Korean sacoglossans containing functional chloroplasts using an
[Table1.] Sacoglossans identified in this study

Sacoglossans identified in this study
lation, PAM). The feeding specificity of the sea slugs was tested using 37 species of cultured algae.
>
Sea slug collection, identification and culture
The sacoglossans (Table 1, Fig. 1) were collected monthly from Wando and Gangneung, Korea from July 2009 until October 2010. They were identified following Koh (2006) and Internet resources on the subject (Sea slug forum 2010). The animals were kept in IMR medium (Klochkova et al. 2006) in 9 × 5 cm Petri dishes at 15-20°C in 12 : 12 h L : D cycles with 15 ㎛ol photons m-2 s-1 provided by cool-white fluorescent bulbs.
The algae used in the feeding experiment, their collection site or source and conditions for laboratory culture are given in Table 2. All algae were grown in IMR medium as described above. While performing feeding experi-
ments for
>
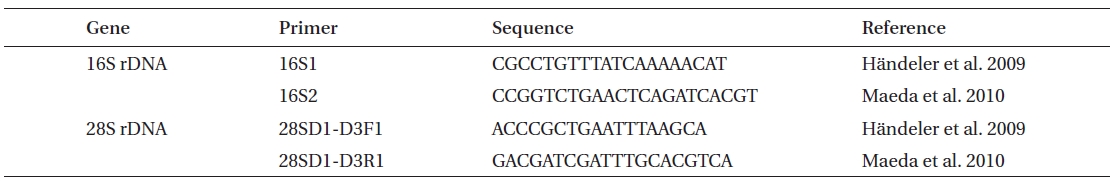
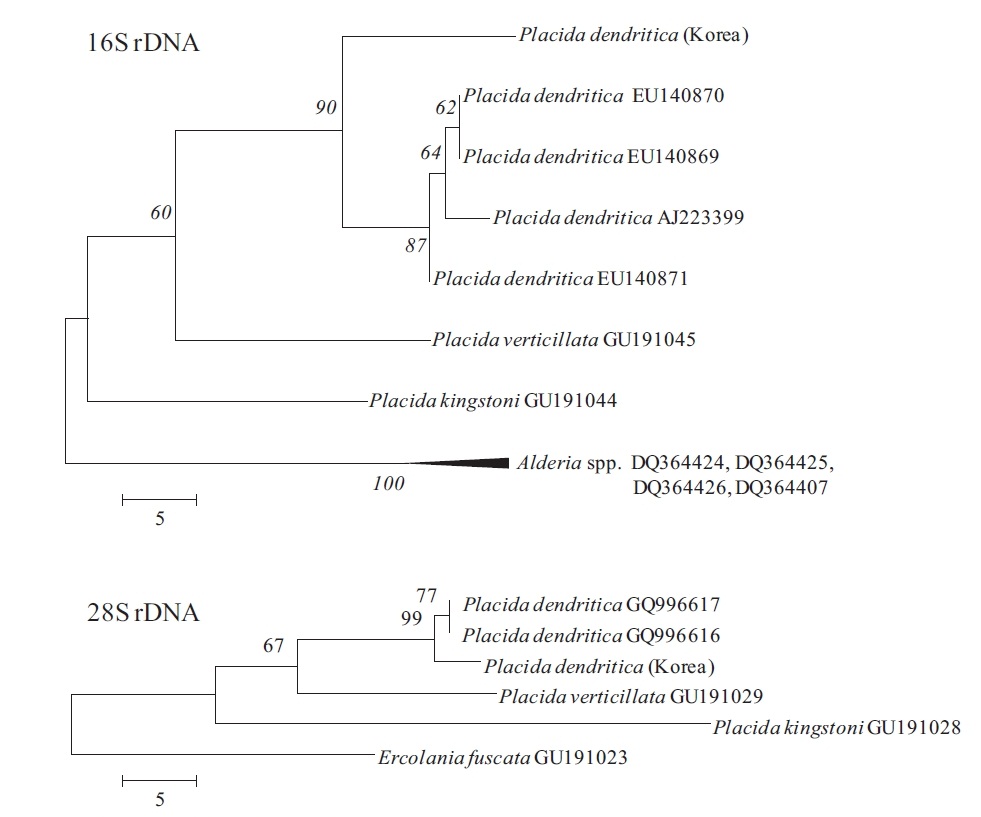
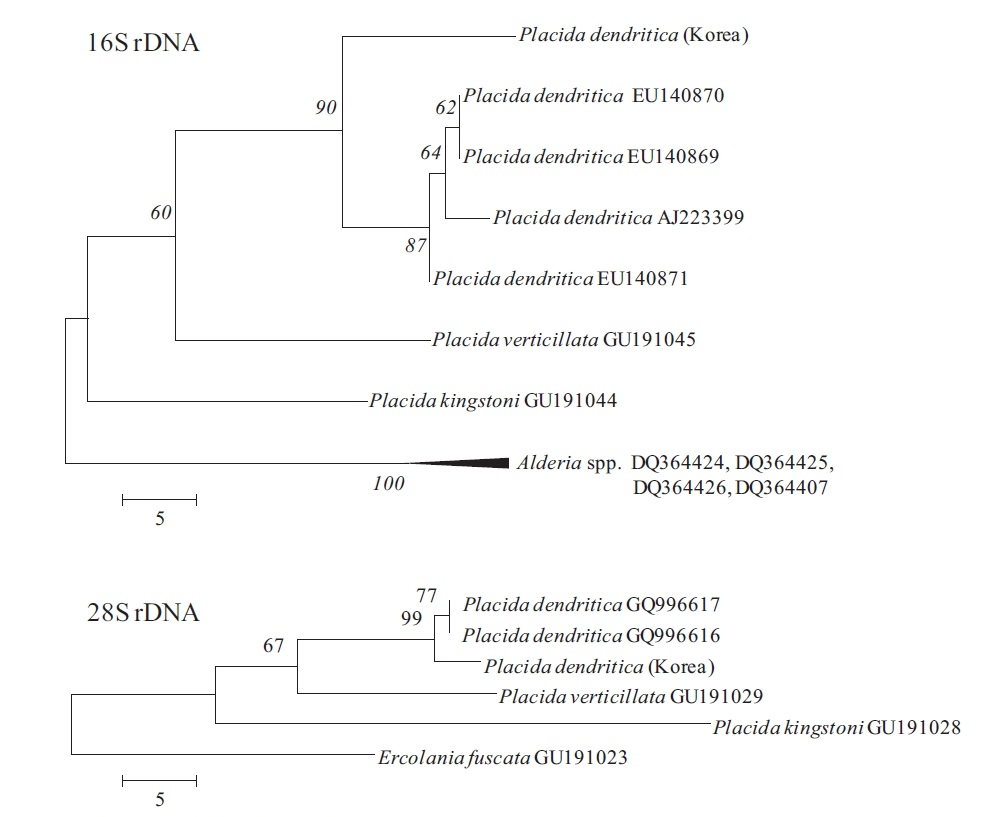
Phylogenetic analyses in Placida dendritica
Genomic DNA was isolated using a genomic DNA isolation kit following the manufacturer’s instructions (Intron Biotech, Seoul, Korea). Polymerase chain reaction (PCR) was performed using primers displayed in Table 3, with an initial denaturation at 95ºC for 3 min, followed by 35 cycles of amplification (denaturation at 95ºC for 20 s, annealing at 55ºC for 30 s and extension at 72ºC for 1 min and 1.5 min for 16S and 28S rDNA, respectively) with a final extension at 72ºC for 10 min. Amplified PCR products were purified using a gel extraction kit (Qiagen, Valencia, CA, USA) and cloned into a T-easy vector (Promega, Madison, WI, USA) to determine DNA sequences. The phylogenetic analyses were performed using sequences for

List of algae used in the slug feeding experiment their collection site and conditions for laboratory culture and results of sacoglossan feeding experiment
version 4.1 (MEGA 2007, Tamura et al. 2007, Kumar et al. 2008). The MP analyses were tested for robustness by bootstrapping with 1,000 replications.
Elysia atroviridis (Fig. 1A)
This species was reported from Korea by Koh (2006) under the name
[Table 3.] Oligonucleotide primers used for amplification of 16S and 28S rDNA
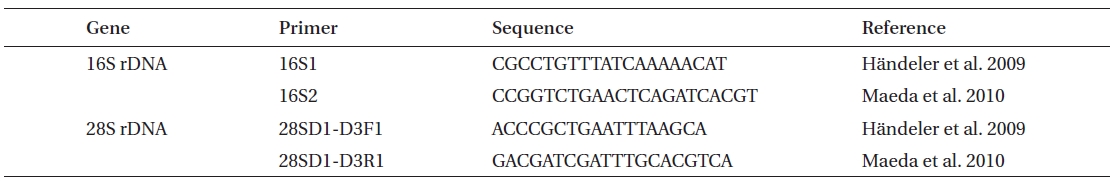
Oligonucleotide primers used for amplification of 16S and 28S rDNA
however, was regarded as a synonym of
Diet. The animals ate little but continuously when food was available. In the field, they were always found on
Seasonality. This species appeared in Wando in December and had peak of population from January to mid
March. During this period, it was possible to collect over 40 animals during one survey. In other months of the year, this species was not detected at this site.
>
Elysia nigrocapitata (
This species was reported from Japan and Hong Kong (Sea slug forum 2010). This is the first report of this species in Korea. Our specimens ranged from several mm to 1.7 cm in length, and ranged in color from light to very dark green when fed and pale grayish-yellow when starved. Parapodia were leaf-like when flattened, with a wide black band along the reverse side edges and thin white band along the upper side edges. The head was black, while the rhinophores and areas around eyespots were white.
Diet. In the field, the animals fed on
Seasonality. In October, 58-214 animals were easily collected from a sampling site in Wando. However, from late November to mid March, only 2-6 animals were collected from the same site during each survey. In other months of the year, this species was absent from this site.
This species was reported from Jeju Island by Koh (2006). We herein extend knowledge on its distribution on the southern coast of Korea.
Diet. The animal ate continuously when food was available. In the field it was found on
Seasonality. During the course of this study, we found 1 animal from Wando site on October 4, 2009 and were unable to find more specimens from the same sampling site at other times during that year.
>
Ercolania boodleae (
Ten animals were found on
Diet.The animals were fed on
Seasonality. In Gangneung, this species was more abundant in spring and the early summer months, correlating with the abundance of
>
Placida dendritica (
Since a number of very similar-looking taxa around the world have been identified as
This species is a new record for Korean sacoglossan fauna, although very common and distributed all around the Korean peninsula. In north Atlantic waters,
Diet.The animals ate continuously when
Seasonality.This species had peak of population in summer and early autumn. From July to mid October, it was possible to collect 50-100 animals from several plants of
This species is a new record for Korean sacoglossan fauna. It is known from Japan and Peter the Great Bay, Russia (Sea slug forum 2010). We herein extend knowledge on its distribution on the southern coast of Korea. Our specimens were aeolid-like in shape; the anterior surface was covered with irregularly arranged and irregularly sized cerata down each side of the body. The body was slightly yellowish to dirty white, fairly transparent, and the color of food in the digestive system showed through the body wall. The rhinophores were tapering and cylindrical, with a dark band about one-third of the way down from the tip.
Diet.This species has been reported to live and feed on the red algae, especially
Seasonality. During the course of this study, we found 5 animals from Wando on August 10, 2009 and were unable to find more specimens from the same sampling site in other months of the year.
This species is a new record for Korean sacoglossan fauna. Three animals were found on
Diet. In the field, the animals fed on
Seasonality. During the course of this study, we found 3 animals from Wando on October 4, 2009. No other specimens were recovered from the same sampling site in other months of the year.
During the course of this study, we collected 7 sacoglossan species from the same site in Wando where
Kawaguti and Yamasu (1965) described functional chloroplasts of
In the case of
Also, Yamamoto et al. (2009) reported a high Fv/Fm value of 0.6 in
In sea slugs feeding on green algae, the diet was restricted to either siphonous or siphonocladous species, especially
This work was supported by Korea Research Foundation grant No. 20100020701 to GHK. We thank Dr. John A. West (University of Melbourne, Australia) for providing many algal strains used in slug feeding experiments and Dr. Nina G. Klochkova (Kamchatka State Technical University, Russia) for her useful consultations on the subject. Special thanks to Ms. Kyong Hwa Cha (Kongju National University) for assistance during sample collection.