Studies on the crossing of
Suto (1963) attempted artificial cross experiments between 5 species including two monoecious (e.g.,
I have been conducting cross experiments between
Mature foliose thalli of
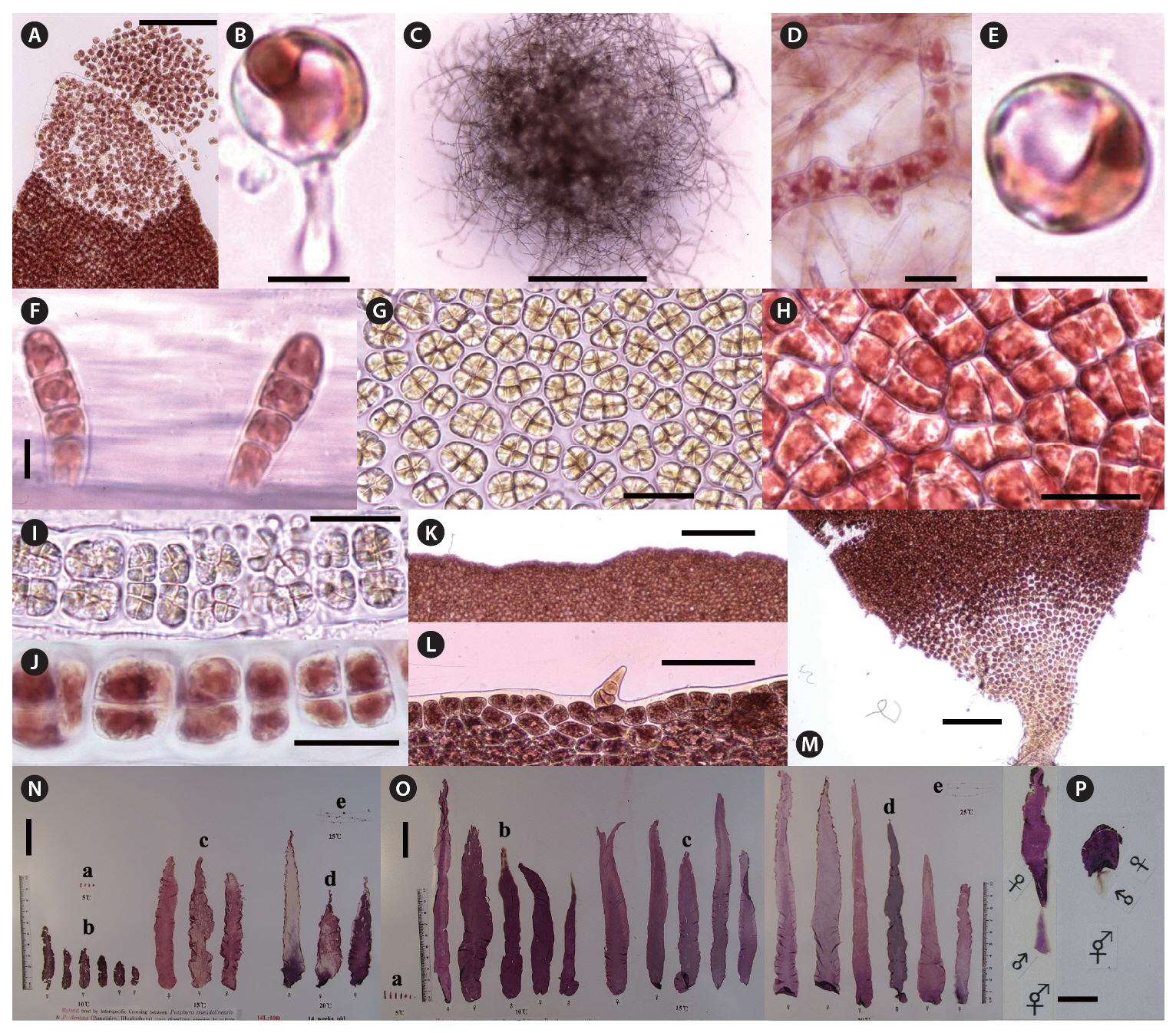
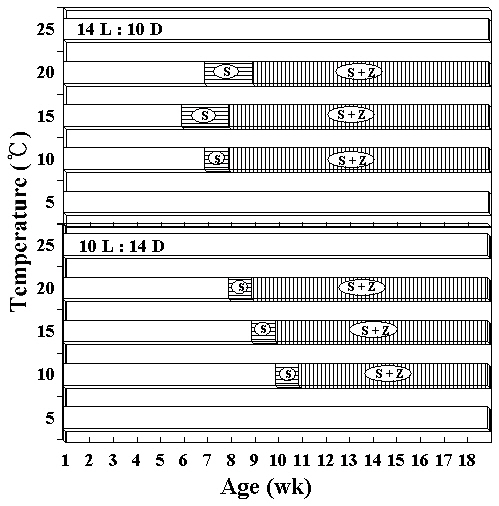
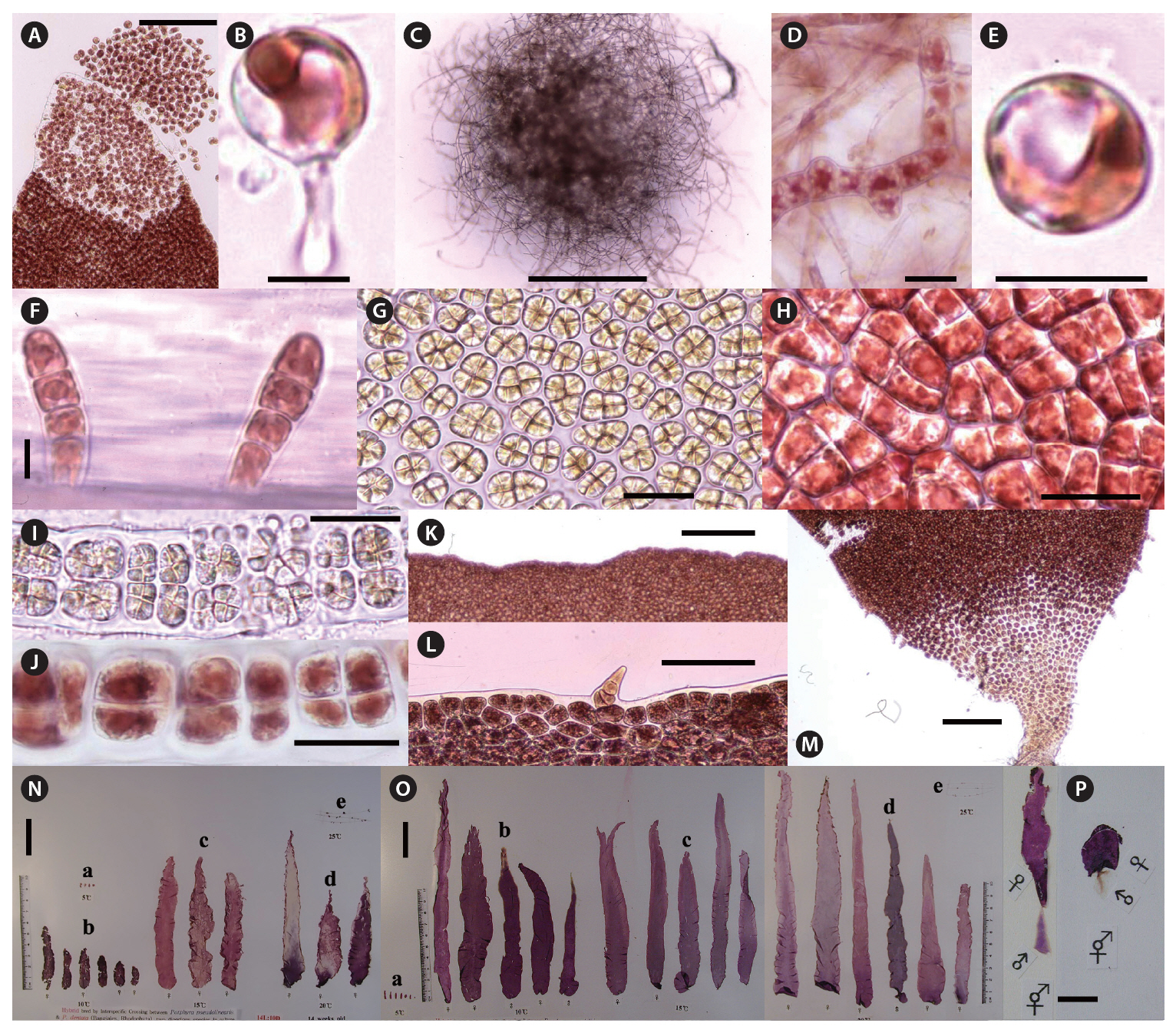
Zygotospores from the hybrid by interspecific crossing between both species were germinated to the conchocelis filaments at 5-25°C (Fig. 1A & B). Conchocelis filaments grew to a colony of about 1.5 mm diameter at 20°C after 3 weeks (Fig. 1C). The conchosporangial branches were produced only from the conchocelis colonies grown at 25°C under both photoperiods (Fig. 1D). Conchospores liberated from the conchosporangial branch were well developed to four cells sporeling (ca. 40 ㎛ long) after 3 days (Fig. 1E & F). Archeospores did not liberate from foliose thalli. Male and female reproductive organs began to occur in the marginal parts of plants, about 10 mm long after 8 weeks at 15°C and 40 ㎛ol m-2 s-1 under 14 L : 10 D (Fig. 1G & H). Mature spermatangia had a Hus’s (1902) fomula 64, (a/4, b/4, c/4) or 128, (a/4, b/4, c/8) (Fig. 1G & I). The zygotosporangia contained 8 zygotospores formed by the division pattern (a/2, b/2, c/2) (Fig. 1H & J). The marginal portions of fronds were entire or micro and macroscopic spinulate processes (Fig. 1K & L). The basal portion of frond has the more clear spinulate processes (Fig. 1M). The largest thalli were obtained at 10-20°C under both photoperiods after 18 weeks in culture. These foliose thalli reached to a blade length of about 130 mm under the long photoperiod, and of about 190 mm under the short photoperiod after 14 weeks. The shape of foliose thalli were linear or lanceolate types (Fig. 1N & O). The zygotospores produced either monoecious thalli (Fig. 1P) or dioecious thalli.
>
Growth of the Sporophyte (conchocelis) phase
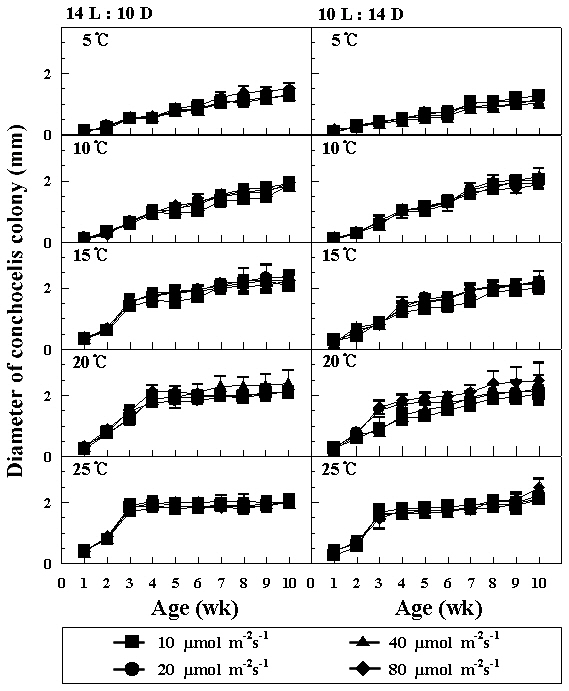
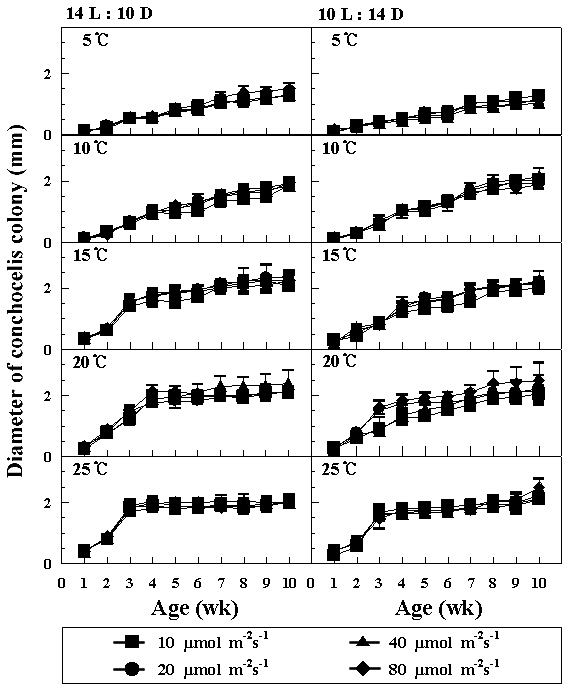
The zygotospores of hybrid were germinated into the conchocelis colonies at 5-25°C and four photon flux densities under both photoperiods. Higher growth of conchocelis was observed at 15 and 20°C under both photoperiods (Fig. 2). There were no remarkable differences in the growth of conchocelis according to photon flux densities. The maximum diameter of conchocelis colonies was about 2.5 mm within 10 weeks.
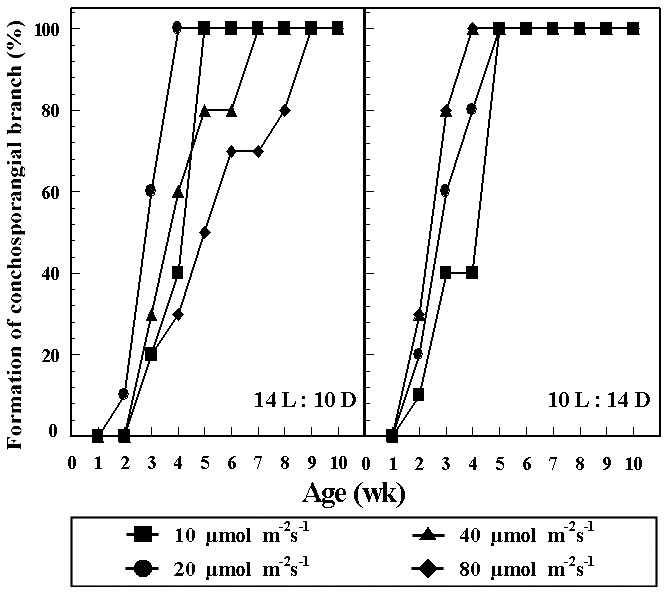
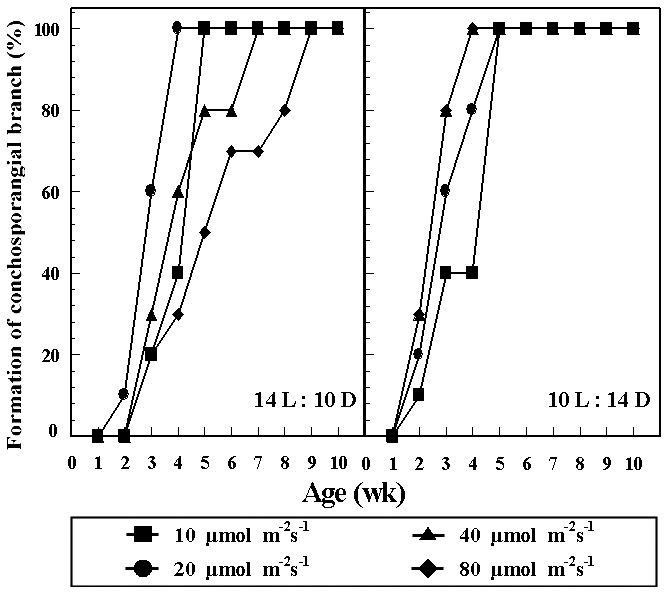
Formation rates of conchosporangial branches at different combinations of temperature and photon flux density are presented in Fig. 3. Conchocelis colonies with conchosporangial branches were produced only at 25°C
and 10-80 ㎛ol m-2 s-1 under both photoperiods within 3 weeks. Formation rates of conchosporangial branches reached 100% within 10 weeks. Conchospores did not liberate at any culture conditions.
>
Growth and maturation of the gametophyte phase
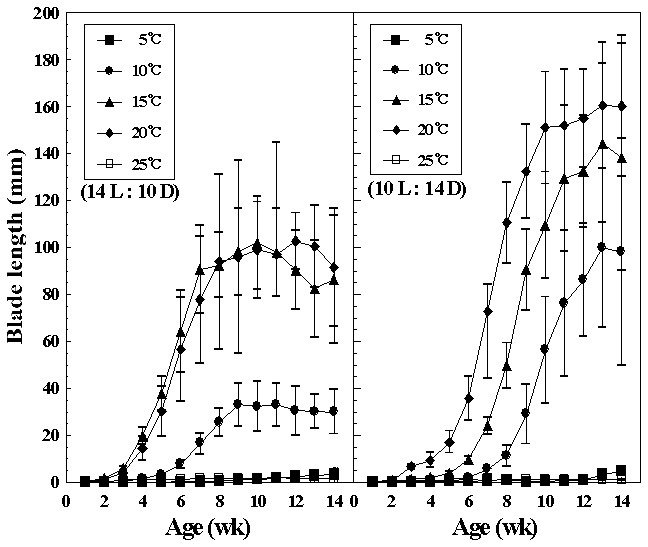
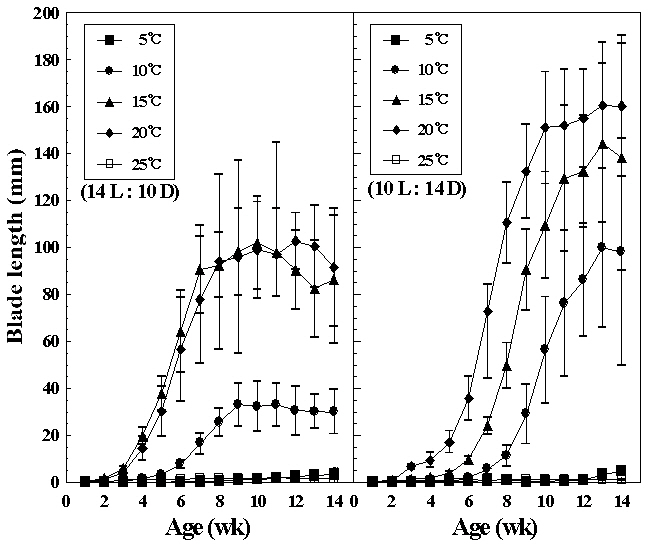
Liberation of conchospores in hybrid species was obtained by reducing the temperature from 25 to 15°C. Growth of foliose thalli at various temperatures under both photoperiods are shown in Fig. 4. Germlings of the hybrid reached their largest size, about 19 cm long at 20°C under short photoperiod after 14 weeks in cul-
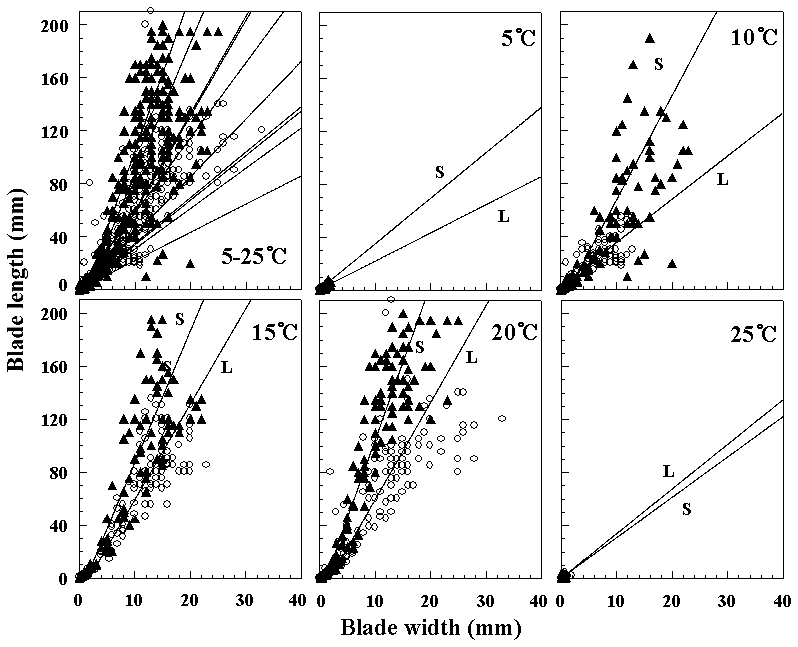
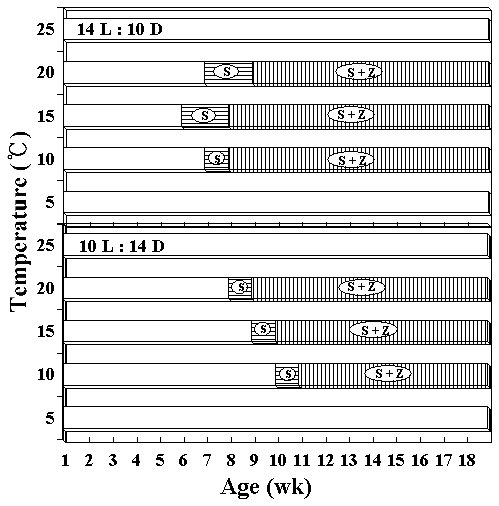
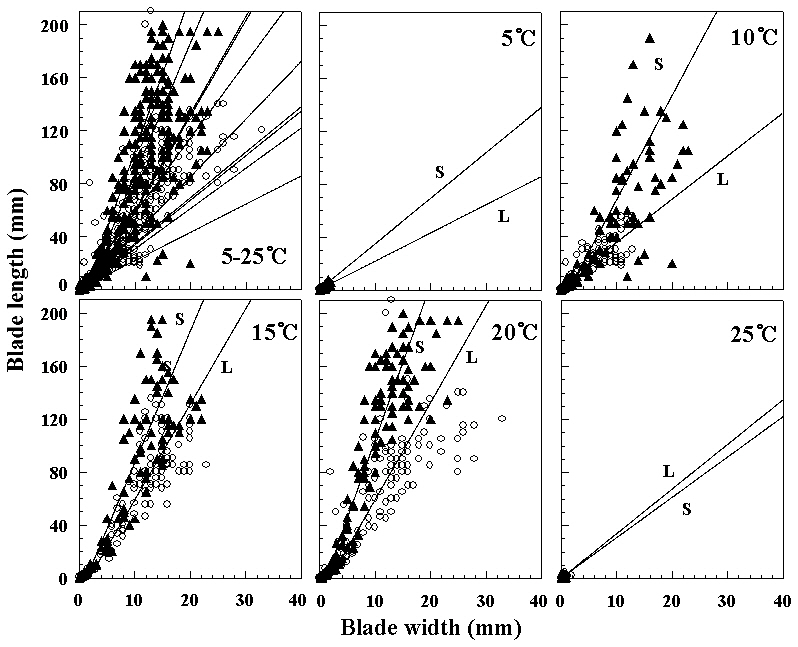
ture, whereas initially thalli grew faster 15°C under long photoperiods. At 5 and 25°C, foliose thalli showed almost no growth. They grew to 10 mm or under in blade length during the culture period. The liberation of spermatia and zygotospores at various temperatures and both photoperiods is presented in Fig. 5. In this hybrid species, archeospores did not liberate in all culture conditions. Their liberation occured at 10 and 20°C under both photoperiods within 11 weeks. The linear or lanceolate shape of fronds, similar to that in nature, was observed under culture conditions that produced rapid growth (Fig. 6).
Hybrid shows a typical biphasic life history of the
sporangia in two species were higher at high temperatures than at low temperatures. This result suggests that the abundance of conchosporangial branches in hybrid were also affected by high temperature (Kurogi and Akiyama 1966). Conchospore liberation in this species was induced by reducing temperature from 25 to 15°C (Kurogi and Akiyama 1966, Iwasaki and Sasaki 1971) and changing photoperiod from 14 L : 10 D to 10 L : 14 D (Kim 1999). In other species for
Notoya et al. (1993) studied four species of Japanese
Kim (1999) reported that the growth of foliose thalli, in
He also reported that
The young thallus with denticulate marginal cell rows was observed at 10°C at first, and it was quickly produced at low temperatures than high temperatures. In this study, the monoecious foliose thalli sectored by horizontal line were produced at 10°C and 10 L : 14 D. Hitherto, some authors (Chang and Zheng 1960, Lindstrom and Cole 1992, 1993) reported dioecious
No archeospores were produced under any culture conditions in hybrid species, and neither monospores nor protothalli were produced from the conchocelis phase.
In the anatomical examination of the hybrid, the divisional formulas of spermatangia according to Hus (1902) well agree with those of Tanaka (1952) but zygotosporangia did not.
Based on the results of this study and previous reports of two species in culture (Kurogi and Akiyama 1966, Notoya et al. 1993) and field investigation (Kang 1966, Wynne 1972, Scagel et al. 1989, Lindstrom and Cole 1992), the temperature needed for the growth and maturation of both phases of hybrid falls between low and high temperatures.
In conclusion, these culture studies indicate that the growth and maturation of both the foliose and conchocelis phase of the hybrid are more influenced by temperature than photon irradiance. Furthermore, it is apparent that the hybrid has monoecious and dioecious thalli, and suggests that hybrid more can be eurythermal cultivar than two cultivars,