Characterization of intact protein structures in the gas phase using electrospray ionization combined with ion mobility mass spectrometry has become an important tool of research. However, the biophysical properties that govern the structures of protein ions in the gas phase remain to be understood. Here, we investigated the impact of host-guest complexation of ubiquitin (Ubq) with macrocyclic host molecules, cucurbit[n]urils (CB[n]s, n = 6, 7), on its structure in the gas phase. We found that CB[n] complexation induces the formation of compact Ubq ions. Both CB[6] and CB[7] exhibited similar effects despite differences in their binding properties in solution. In addition, CB[n] attachment prevented Ubq from unfolding by collisional activation. Based on the experimental results, we suggest that CB[n]s prevent unfolding of Ubq during transfer to the gas phase to promote the formation of compact protein ions. Furthermore, interaction with positively charged residues per se is suggested to be the most important factor for the host-guest complexation effect.
The advancement of electrospray ionization (ESI) in combination with ion mobility mass spectrometry (IM-MS) enabled characterization of protein tertiary and quaternary structures using MS.1,2 In addition to mass-to-charge ratio (
Host-guest chemistry provides a unique means to investigate structural changes of molecules in response to specific interactions. A recent study by Pagel and coworkers showed that noncovalent attachment of a host molecule, 18-crown-6 (18C6), to protein ions caused their structural compaction.4 This observation was explained by microsolvation of the charged lysine side chains by 18C6 preventing their structural rearrangements in the gas phase. However, underlying mechanism for the observation has not been characterized in detail. Understanding structural changes of protein ions induced by specific interactions would provide deeper insights into the structural properties of protein ions in the gas phase.
Here, we investigated the influence of host-guest complexation on the structure of ubiquitin (Ubq) ions in the gas phase. Cucurbit[
>
Materials and sample preparation
Ubq from bovine erythrocytes, equine cytochrome c (Cyt c), equine myoglobin (Myb), CB[6], CB[7], and formic acid were purchased from Sigma-Aldrich (Saint Louis, MO, USA). HPLC-grade water and acetonitrile were purchased from Avantor Performance Materials, Inc. (Center Valley, PA, USA) and used as solvents. Protein solutions were prepared as 10 μM, and CB[6] or CB[7] was added to be 100 μM concentration.
ESI-IM-MS experiments were performed with a Synapt G2 HDMS quadrupole travelling wave ion mobility orthogonal time-of-flight mass spectrometer equipped with a Z-spray ionization source (Waters, Milford, MA, USA). The capillary, sampling cone, and extraction cone voltages were set as 2.00 kV, 40 V, and 2 V, respectively. The helium cell and drift cell gas flows were 180 mL min-1 and 30 mL min-1, respectively, with wave voltage and height of 450 m s-1 and 12 V, respectively. Calibration of measured arrival times from IM experiments into ΩD values was performed by measuring arrival times of Ubq, Cyt c, and Myb and creating a calibration curve using their previously reported ΩD values.10 Collisional activation experiments were performed in the trap cell of the Synapt G2 instrument with a gas (argon) flow of 2.0 mL min-1.
>
Structures of Ubq and its complex ions
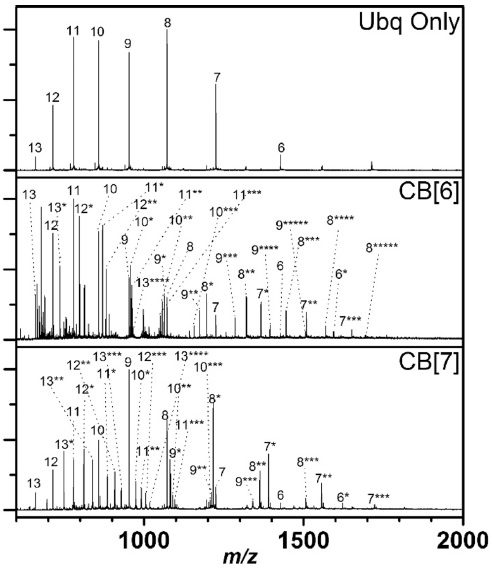
Figure 2 shows that Ubq ions with various charge states are formed upon ESI of a Ubq solution. With addition of CB[6] or CB[7] to a Ubq solution, Ubq ions complexed with CB[
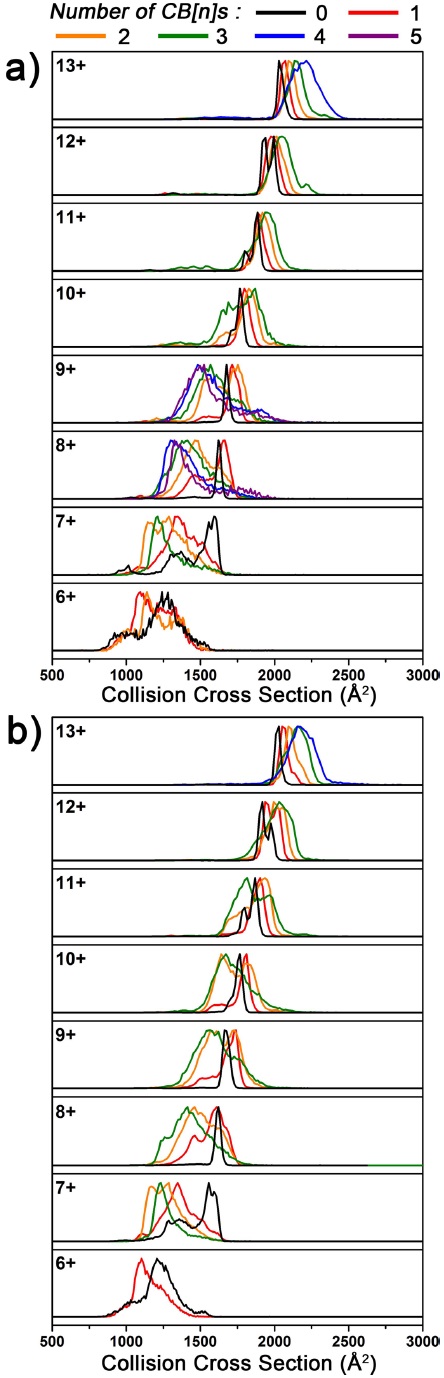
ΩD values of both Ubq and its complex ions generally increased with increasing charge states (Figure 3). This trend is well known, and can be attributed to strong electrostatic repulsion between positively charged sites resulting in expansion of highly charged ions.12 On the other hand, comparison between uncomplexed and complexed ions of identical charge state reveals previously unreported trend in their structures. Three trends in ΩD distributions of the ions were observed depending on their charge states, which are: 1) decrease in ΩD (low charge states), 2) increase in ΩD, followed by decrease in ΩD with further CB[
18C6 complexation has previously been shown to cause decreased ΩD values of protein ions.4 Still, ΩD decrease by CB[
Then, the structures of CB[6] and CB[7] complex ions were compared (Figure 3). It was observed that the structures of CB[6] and CB[7] complex ions are similar. Especially, the structures of 6+ to 9+ charged complexes are highly similar. This result implies that CB[6] and CB[7] share the most important factor driving the generation of compact complex ions. The capability to interact strongly with positively charged groups is the most probable candidate, because CB[6], CB[7], and 18C6 all share this property. Although CB[7] preferentially interacts with hydrophobic guests in solution, the binding property of CB[7] was reported to be altered during transfer to the gas phase to interact strongly with positively charged guests.13 In addition, even if CB[7] is bound to a hydrophobic guest through one of its two portals, the other portal can accommodate another positively charged group simultaneously.13,14
Some differences were observed between CB[6]and CB[7] complex ions at higher charge states (Figure 3). The ΩD values of 10+ to 12+ charged ions with three CB[7] attachments are smaller than those with three CB[6] attachments, suggesting that CB[7] is more effective in stabilizing compact structures of protein ions than CB[6], despite its larger size. A possible cause is the simultaneous interactions of CB[7] with multiple residues, as discussed above, which would allow CB[7] to further promote the generation of more compact Ubq ions.14
>
Collisional activation of Ubq ions
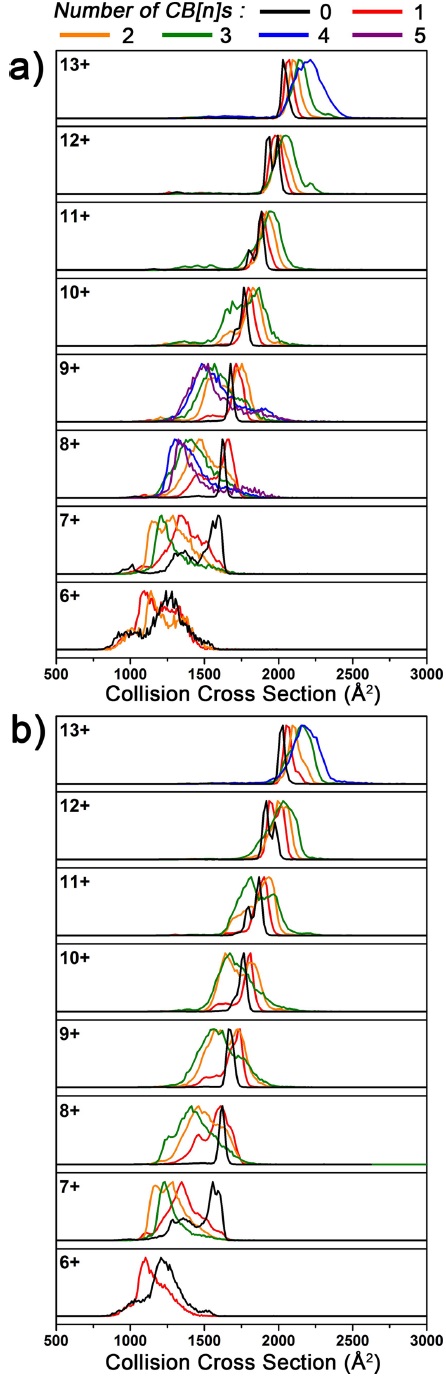
Noncovalent interactions between CB[
>
Mechanism of compact Ubq ion formation by CB[n]s
It is important to consider the stage during ionization and MS analysis at which CB[
A possible reason that CB[
Most reasonable explanation is that the interaction with positively charged sites is critical for the structural compaction, because this is the only common factor between CB[
We have studied the gas-phase structures of host-guest complex ions comprising a protein and macrocyclic hosts. It was found that CB[
![Schematic structure of CB[n] and three-dimensional structure of CB[7].](http://oak.go.kr/repository/journal/20506/E1MPSV_2016_v7n1_16_f001.jpg)
![Mass spectra of Ubq (10 μM), and Ubq with CB[6] (100 μM) or CB[7] (100 μM) in 50/50 water/acetonitrile solution with 1% formic acid (positive ion mode). Asterisks (*) denote the number of CB[n]s on the Ubq ions.](http://oak.go.kr/repository/journal/20506/E1MPSV_2016_v7n1_16_f002.jpg)
![Ion mobility spectra of Ubq and its CB[n] complex ions from 50/50 water/acetonitrile solution with 1% formic acid: a) with CB[6], and b) with CB[7].](http://oak.go.kr/repository/journal/20506/E1MPSV_2016_v7n1_16_f003.jpg)
![Ion mobility spectra of Ubq and its CB[n] complex ions after applying 20 or 40 V of collision energy (CE): a) CB[6], b) CB[7].](http://oak.go.kr/repository/journal/20506/E1MPSV_2016_v7n1_16_f004.jpg)