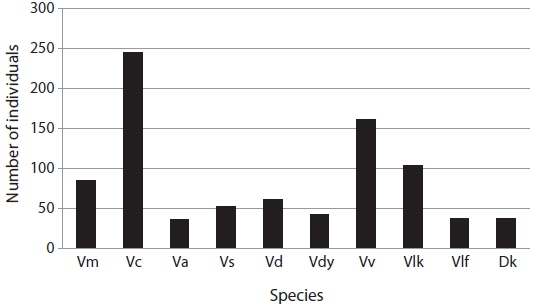
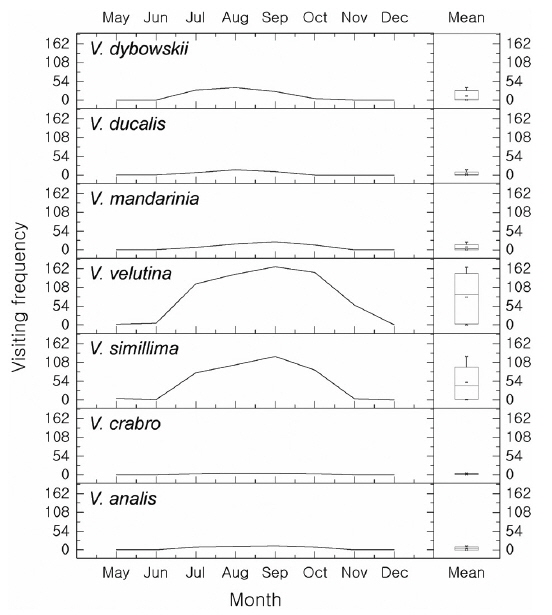
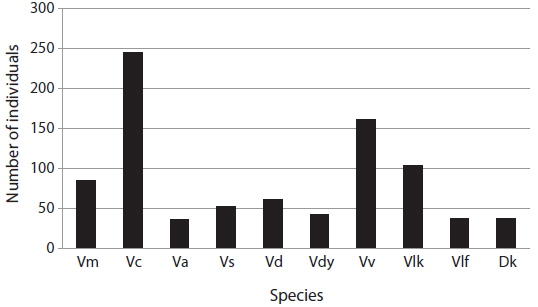
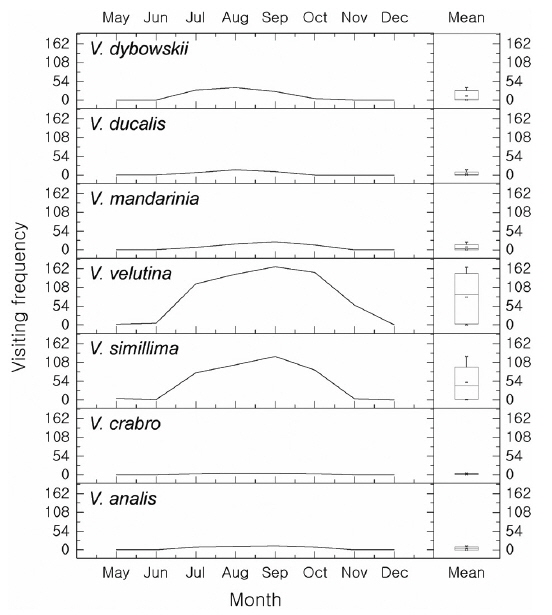
The aim of this study was to assess the occurrence and foraging of social and other wasps and bees in Jirisan National Park (JNP, South Korea), in particular in an apiary. Sixty one traps were placed in the southwestern part of JNP to identify social wasps from July to September 2014, and the damage to the apiary caused by wasps or bees was observed once a month from May to December 2014 between 10 a.m. and 5 p.m. In total, 10 species of Vespidae were collected by trapping. Vespa crabro was the most abundant (245 individuals, 28.3%), followed by Vespa velutina (162 individuals, 18.7%). In the apiary, however, V. velutina was the most frequent species. V. velutina visited a maximum of 167 times a day in September, which corresponded to one visit in 2.5 min. Accordingly, these data are in line with the most serious impact of V. velutina on the apiaries in South Korea. V. simillima was the second most frequent species; both Vespa species hawked honeybees. Even though the occurrence of V. mandarinia was low, this species caused serious damage by mass slaughter of honeybees. The occurrence of V. crabro, V. analis and V. ducalis was quite low and their impact on honeybees was negligible. There have been few reports of V. dybowskii foraging for honeybees, but they are considered to be a new pest because their impact on apiaries is considerable. Most Vespa species attacked the apiary from June to October, with a maximum in September. However, V. velutina visited until November to early December. Vespula species are not more serious pests than Vespa species, but many adults were observed stealing honey from beehives. Polistes, Orancistrocerus, and Bombus species had no impact on honeybee colonies in the apiary.
Pollination in terrestrial ecosystems is an important service supported by various pollinators. Their economic value is estimated at about 120 billion dollars a year (Costanza et al. 1997). The major insect pollinators are honeybees; their economic benefit is 11.7 billion dollars (77% of the total pollination value) in the United States (Calderone 2012) and 5.8 billion dollars in South Korea (Jung 2008). Honeybees have been cultivated since prehistoric times. They are one of the top three livestock resources in Europe (Tautz 2008).
However, Colony Collapse Disorder (CCD) has seriously affected honeybees all over the world since 2006 and caused enormous economic damage to agricultural crops (Johnson 2010). The number of apiaries and hives had constantly increased in Korea since 1970s, but a downward trend has continued after a peak in 2005 (Lee et al. 2010). This phenomenon seems to be caused by a decline in honey productivity because of sacbrood virus disease and an increase in the abundance of natural honeybee enemies (Kim et al. 2006, Kim et al. 2008, Jung et al. 2008). The best-known natural enemies of honeybees are hornets (
Recently, the invasive yellow-legged hornet
Jirisan National Park (JNP) is the largest national park (total area: 483.022 km2) and one of the densest apicultural areas in South Korea (Lee et al. 2010). Various social wasps have damaged many apiaries within this region and a recent
In this study, to identify the vespid wasps in JNP, we collected them by trapping in the southwestern region of JNP. To investigate their foraging behaviors and to assess their impact on honeybees, we also determined the number of wasps and bees visiting an apiary in JNP. These data will be useful for protection of honeybee colonies.
To identify the species of social wasps, we placed 61 traps (the same as in the study by Choi et al. 2012a) at different sites in the southwestern part of JNP (Fig. 1) and collected trapped wasps once a month from July to September 2014.
>
Observation of foraging behavior
To inspect the damage caused by wasps or bees, their foraging and predation behavior was recorded by observing the hive entrance. The apiary site (35°15′18.37″127°35′53.88″, 402 m above sea level) is a small farm with 6
One-way PERMANOVA (permutation multivariate analysis of variance) was used to test the independent effects of month and species. The non-parametric PERMANOVA test calculates a pseudo-
>
Social wasps collected by trapping
In total, 10 Vespidae species were identified (Fig. 2). Among them,
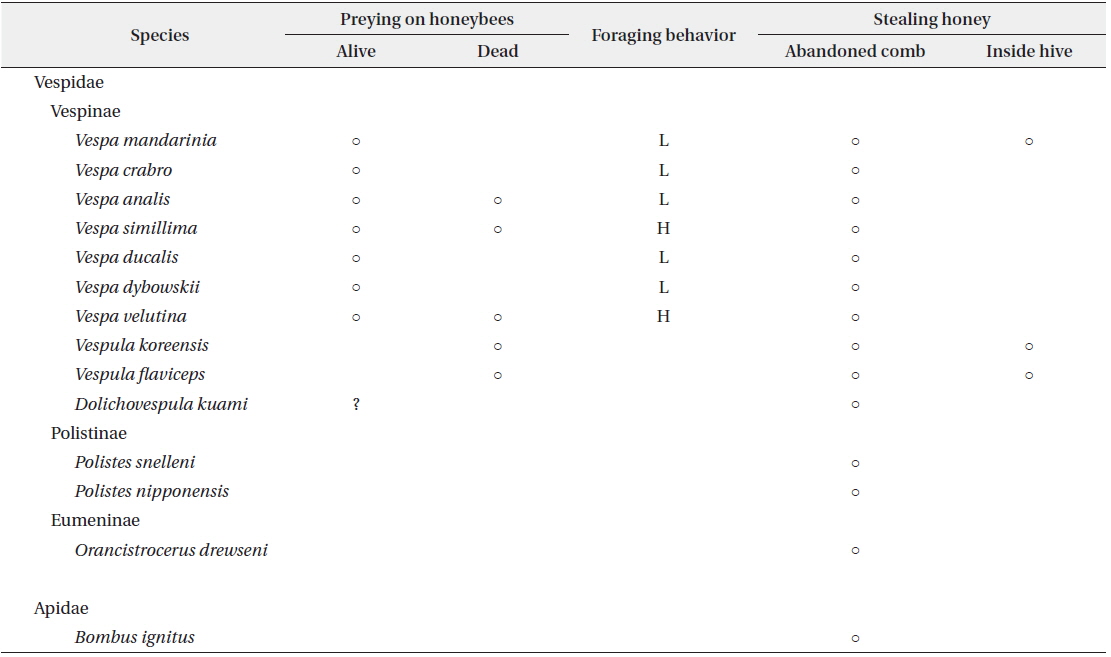
At the apiary site, 14 wasp and bee species were collected, including 13 species of Vespidae and one species of Apidae (Table 1). These included all species of Vespinae that we collected at other JNP sites. The species of wasps and bees in the JNP apiary were very similar to those reported in other domestic apiaries (Chang et al. 1993, Jung et al. 2007).
[Table 1.] Foraging behavior of wasps and a bumble bee in the apiary
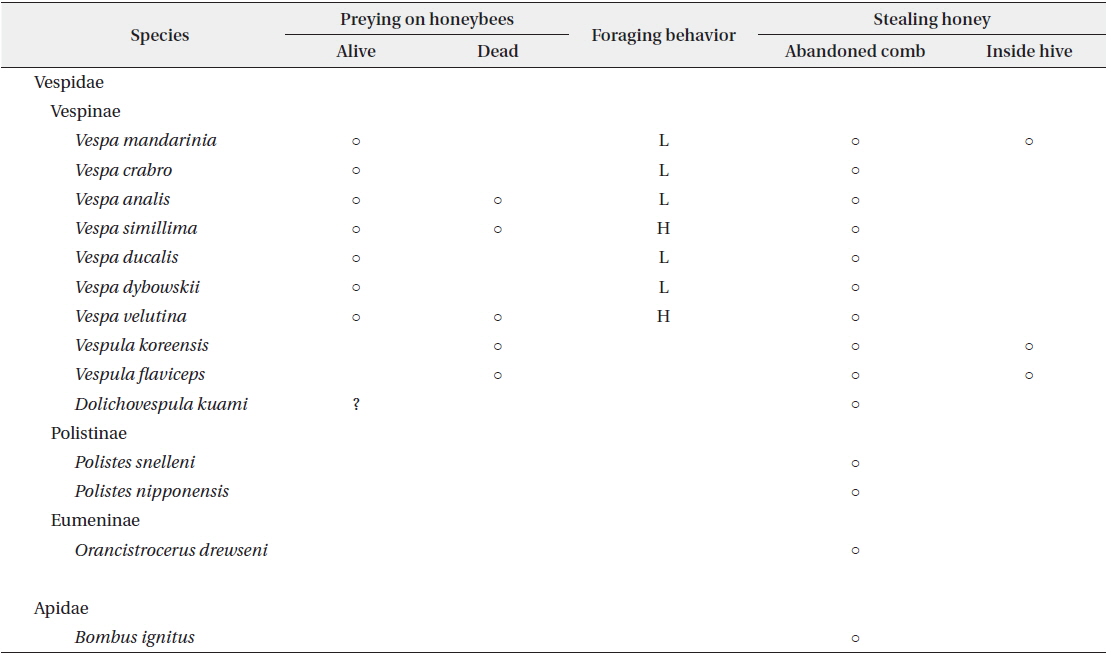
Foraging behavior of wasps and a bumble bee in the apiary
>
Foraging of Vespidae in the apiary
Genus Vespa
The hornets have the largest and most continuous impact on apiaries (Matsuura and Yamane 1990). In total, 7 species were found in the apiary and all of them attacked beehives and predated on honeybees (Figs. 3 and 4). The visiting frequency of
Although the number of trapped
According to Jung et al. (2008), soon after its introduction,
Genera Vespula and Dolichovespula
Others species
Polistes species and