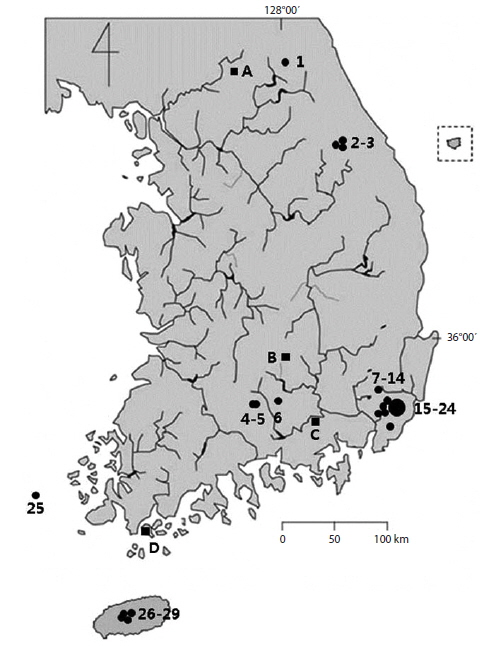
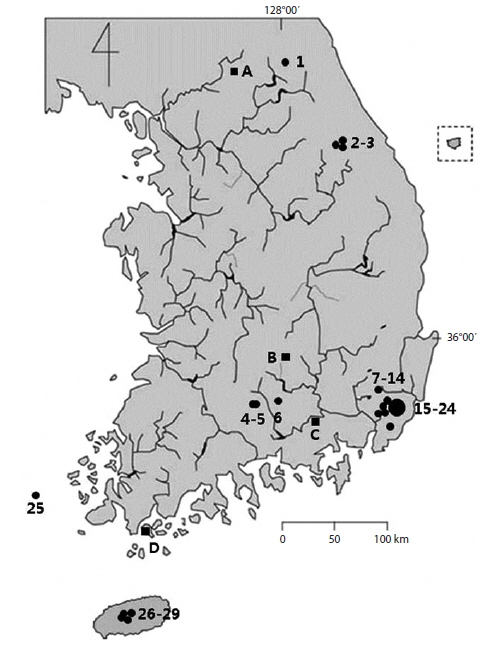
Stauroneis diatoms were collected from the 38 sites in 32 freshwater areas (Fig. 1). A total of twenty five Stauroneis species were found in these local areas, but mainly in the peatlands, mountain areas, and oligotrophic freshwaters. Some highly contaminated freshwaters were chosen for this study, but the Stauroneis were not found at all or scarce, and these areas were ignored. Important morphological characters in the classification of Stauroneis diatoms are the outline of the valve, the form of the valve ends, the density of striae and areola on a stria, and the shape of hyaline stauros in the central area (Van de Vijver et al. 2004). The characteristics of each species are given as follow and the morphological difference among all neighboring species are emphasized. The dimensions of the valve and strial density are presented as the local data of each species. The distributional patterns are briefly summarized. Among twenty five species, sixteen are newly reported to Korea and are marked with asterisks (*) in front of the scientific name.
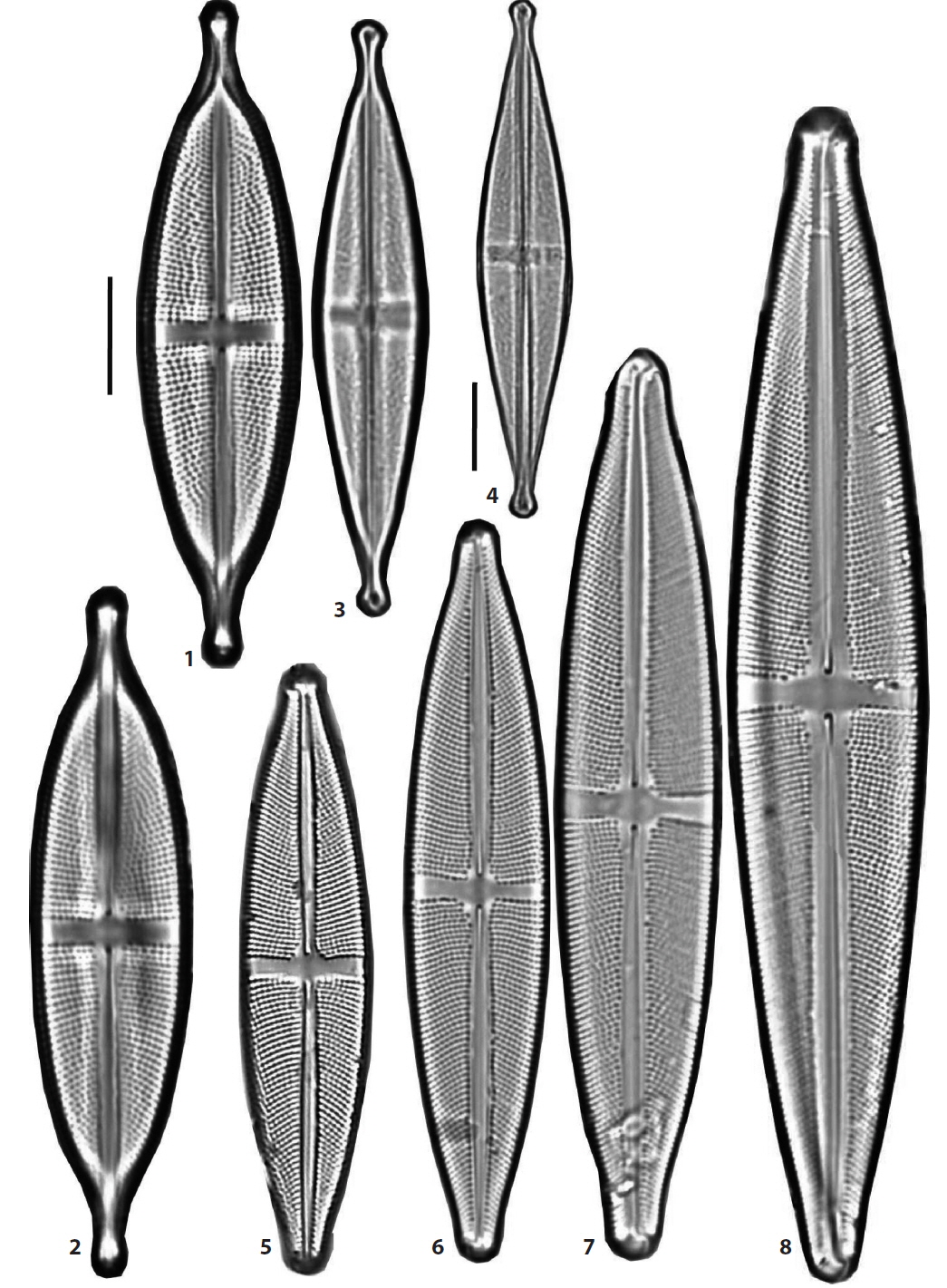
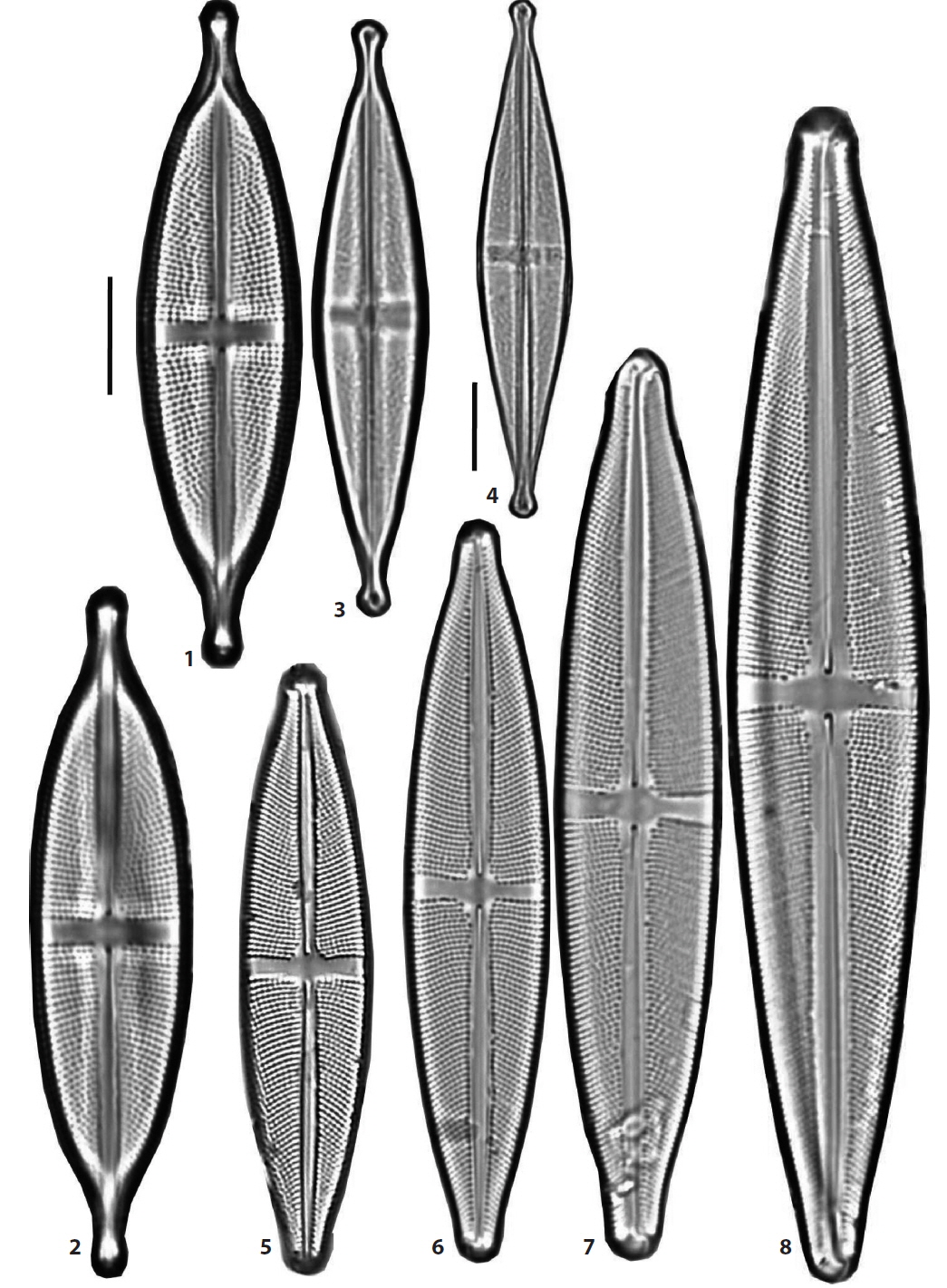
(1) Stauroneis alabamae Heiden in Schmidt et al. 1903 (Pl. 4, Fig. 6) (Schmidt 1903, pl. 242: 2; Hustedt 1930, p. 257, f. 412). Valves broadly elliptical-lanceolate in outline, the breadth of the valve abruptly narrow towards the ends. The hyaline stauros of the central area narrow rectangular, hardly expanded towards the ends. The ends of the valve not protracted, obtusely round. Valves 168–170 μm in length, 22.5–23.5 μm in breadth, striae coarse 21–23 rows in 10 μm, puncta in a stria distantly spaced, 18–19 in 10 μm. Occurring rarely in lowland wetlands in Haman (Chung and Noh 1987) and recently in Dongsong Reservoir in Cheolwon.
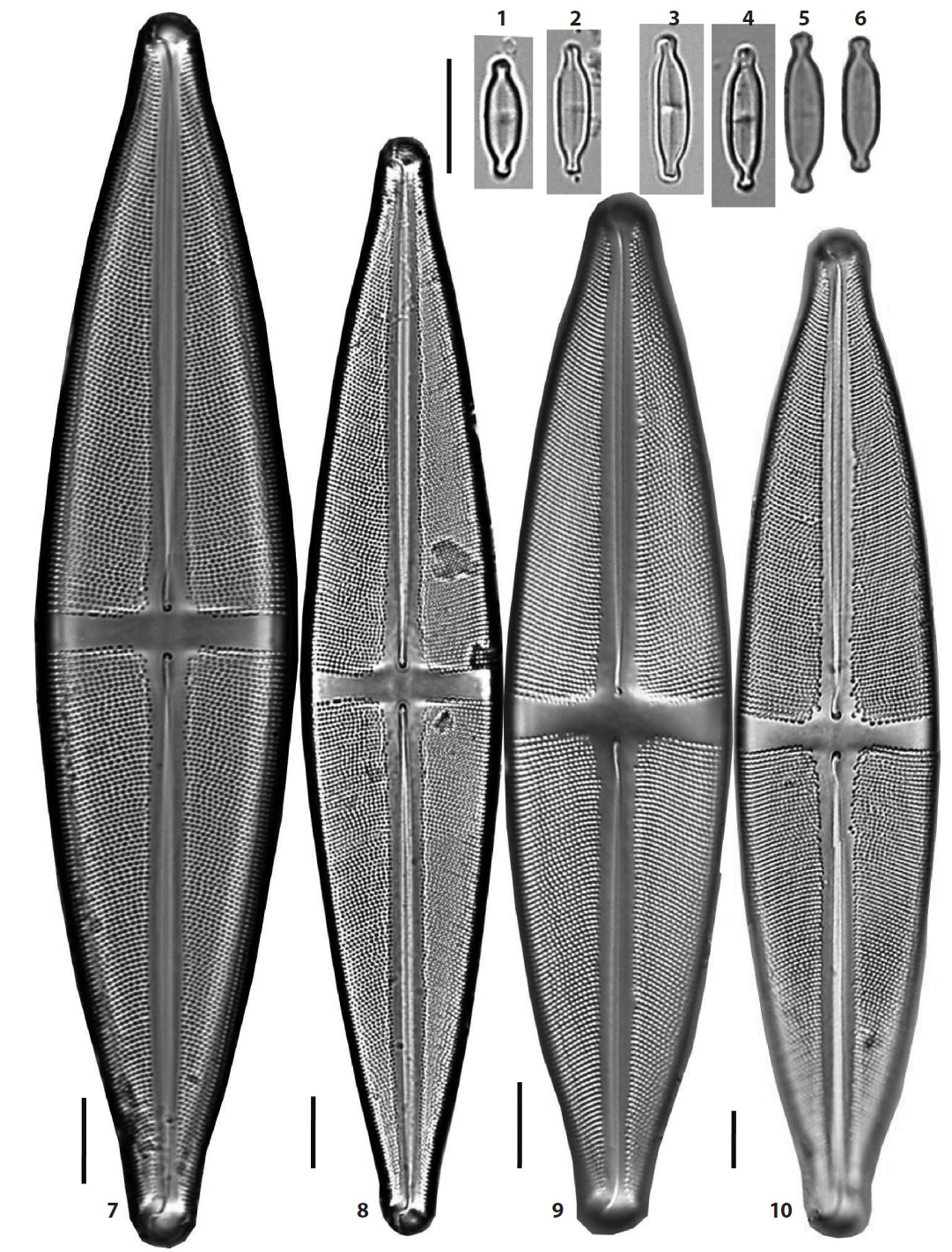
(2) *Stauroneis amphicephala Kützing 1844 (Pl. 1, Figs. 4–6) (Van de Vijver et al. 2004, p. 20, pl. 51: 1–10, pl. 52: 1–5, pl. 53: 1–4, pl. 54: 1–5). Valves elliptical-lanceolate in outline, the ends of the valve protracted and rostrate. The stauros more expanded towards the ends. Valves 45–55 μm in length, 11–13 μm in breadth and striae 19–22 rows in 10 μm. Occurring rarely in a rice paddy in Wando, in June 2012, and newly reported to Korea.
(3) *Stauroneis ancepsopsis Lange-Bertalot, Cavacini, Tagliaventi & Alfinito 2003 (Pl. 2, Figs. 1 and 2) (Lange-Bertalot et al. 2003, p. 132, pl. 38: 15, pl. 125: 2–7). Valves elliptical-lanceolate to elliptical in outline, the ends of the valve abruptly long protracted and rostrate. The stauros rectangular and not expanded towards the margins. Valves 55–60 μm in length, 13.5–14 μm in breadth, striae 18–20 rows and puncta of the striae 17–19 in 10 μm. This species is differentiated with S. anceps Ehrenberg sensu stricto from the form of the valve ends. Occurring rarely in rice paddy in Wando, and newly reported to Korea.
(4) Stauroneis anceps Ehrenberg 1843 sensu stricto (Pl. 1, Figs. 7 and 8) (Lange-Bertalot et al. 2003, p. 132, pl. 35: 1-12, pl. 38: 14; Van de Vijver et al. 2004, pl. 43: 1, 2; Bahls 2011a). Valves lanceolate to linear-lanceolate in outline, the ends of the valve moderately protracted and rostrate. The stauros rectangular and slightly expanded towards the margins. Valves 32.5–43 μm in length, 9.5–10 μm in breadth, striae 22–24 rows and puncta of the striae 20–28 in 10 μm. Reichardt (1995) designated the lectotype for this species from a specimen in Ehrenberg’s type material collected from Cayenne, French Guyana (Van de Vijver et al. 2004). This species is the most popular and common in Korean freshwaters and was reported 22 times before 1997 (Lee 1997). However, S. anceps sensu stricto is found less frequently, occurring rarely in lowland wetlands in Haman.
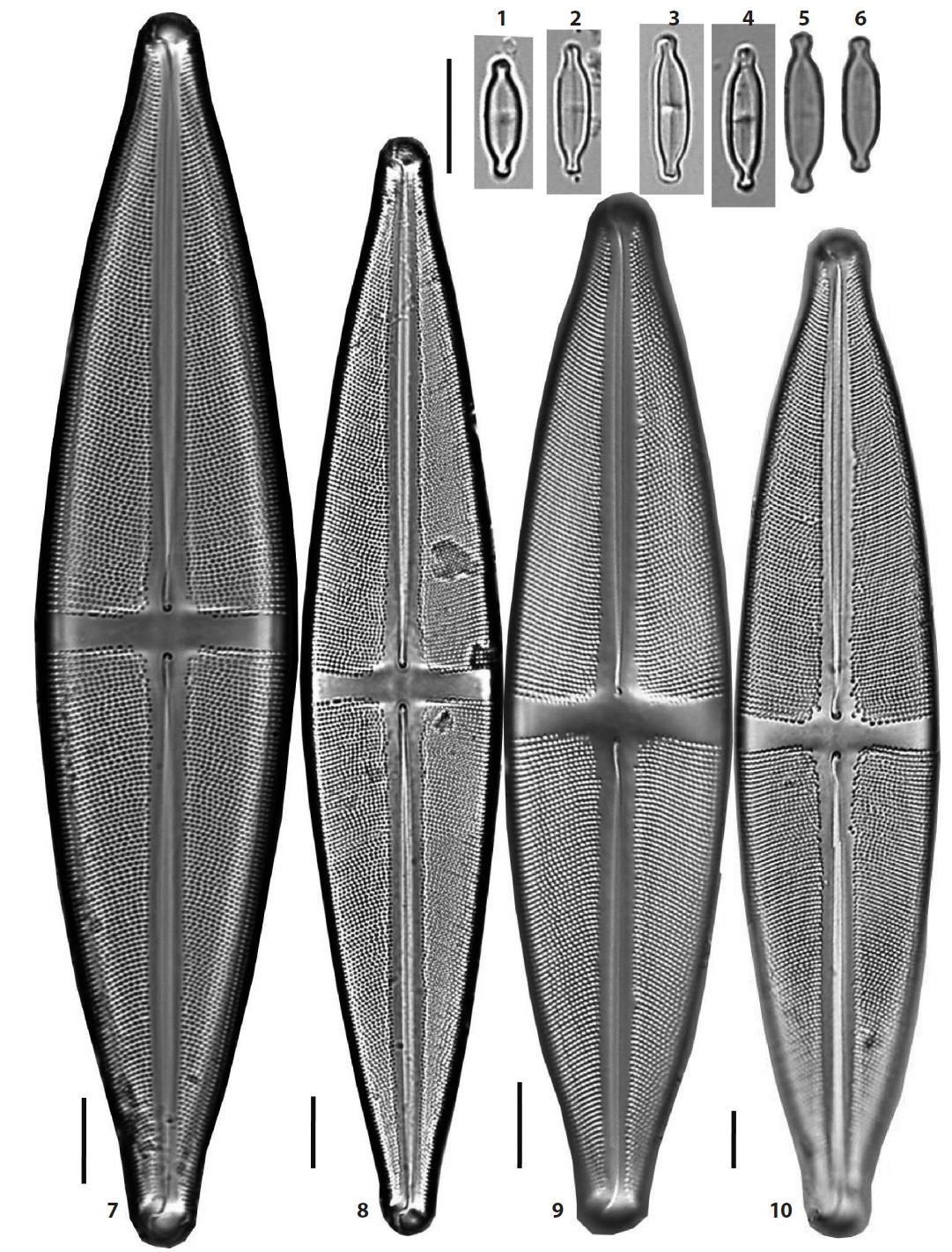
(5) *Stauroneis borgei Manguin 1941 (Pl. 5, Fig. 8). Synonym: Stauroneis smithii var. borgei (Manguin) Hustedt 1959 (Hustedt 1959, p. 811, f. 1157h–k). Valves elliptical to rhombic-lanceolate with convex margins, the ends of the valve not protracted, and pseudosepta developed near the ends. Axial area narrow and stauros in the central area rectangular, extending to the margins of the valve. Valves 20 μm in length, 7.6 μm in breadth and striae fine and not resolved in light microscopy. Occurring only in a lake, Dongsong Reservoir in Cheolwon, and newly reported to Korea.
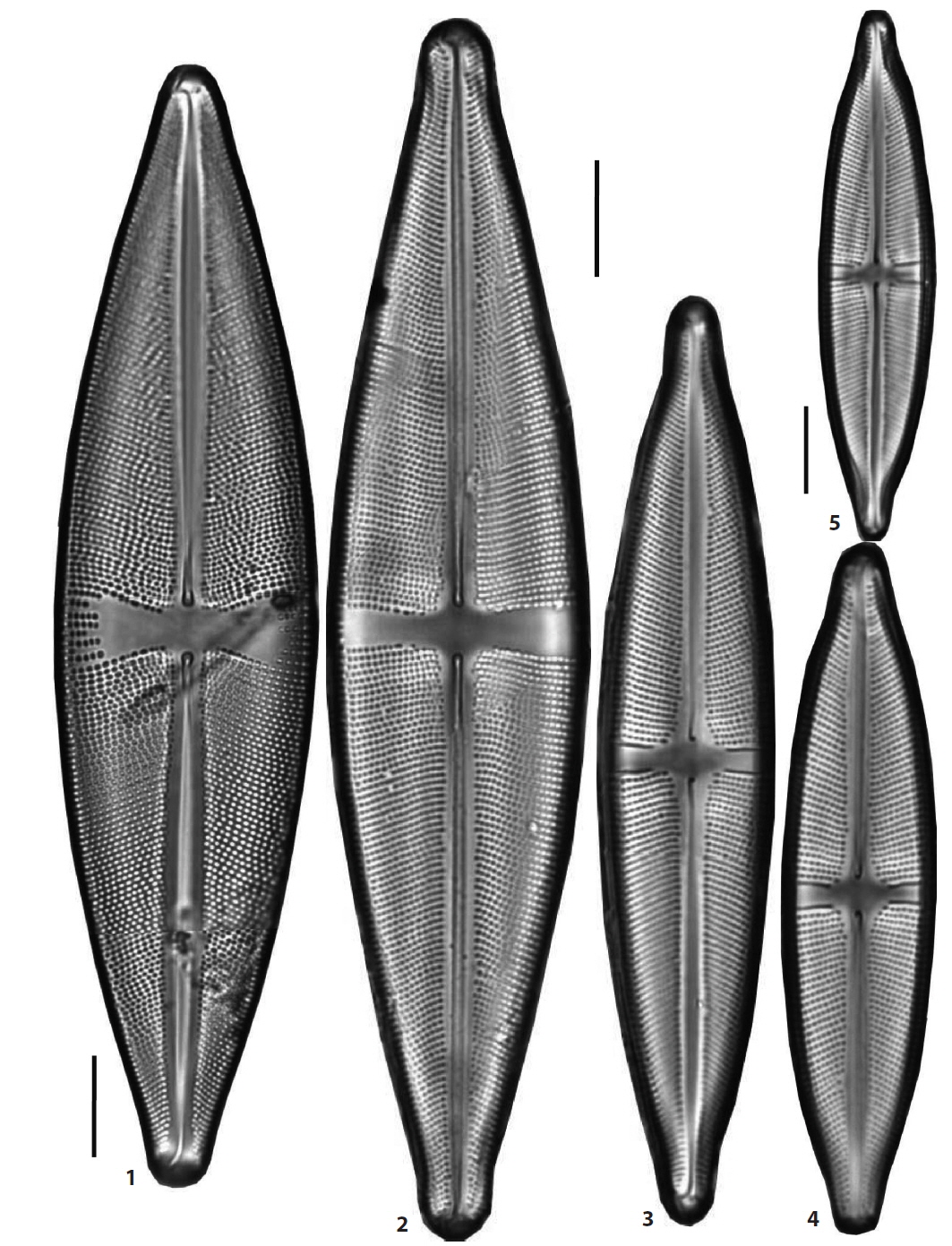
[Plate 5.] Figs. 1, 2. Stauroneis prominula, Fig. 3. S. smithii, Figs. 4?7. S. pseudosmithii, Fig. 8. S. borgei, Figs. 9, 10. S. sagitta, Fig. 11. S. kriegeri, Figs. 12, 13. S. sturolineata var. japonica. Scale bars, 10 μm (×2000 magnification).
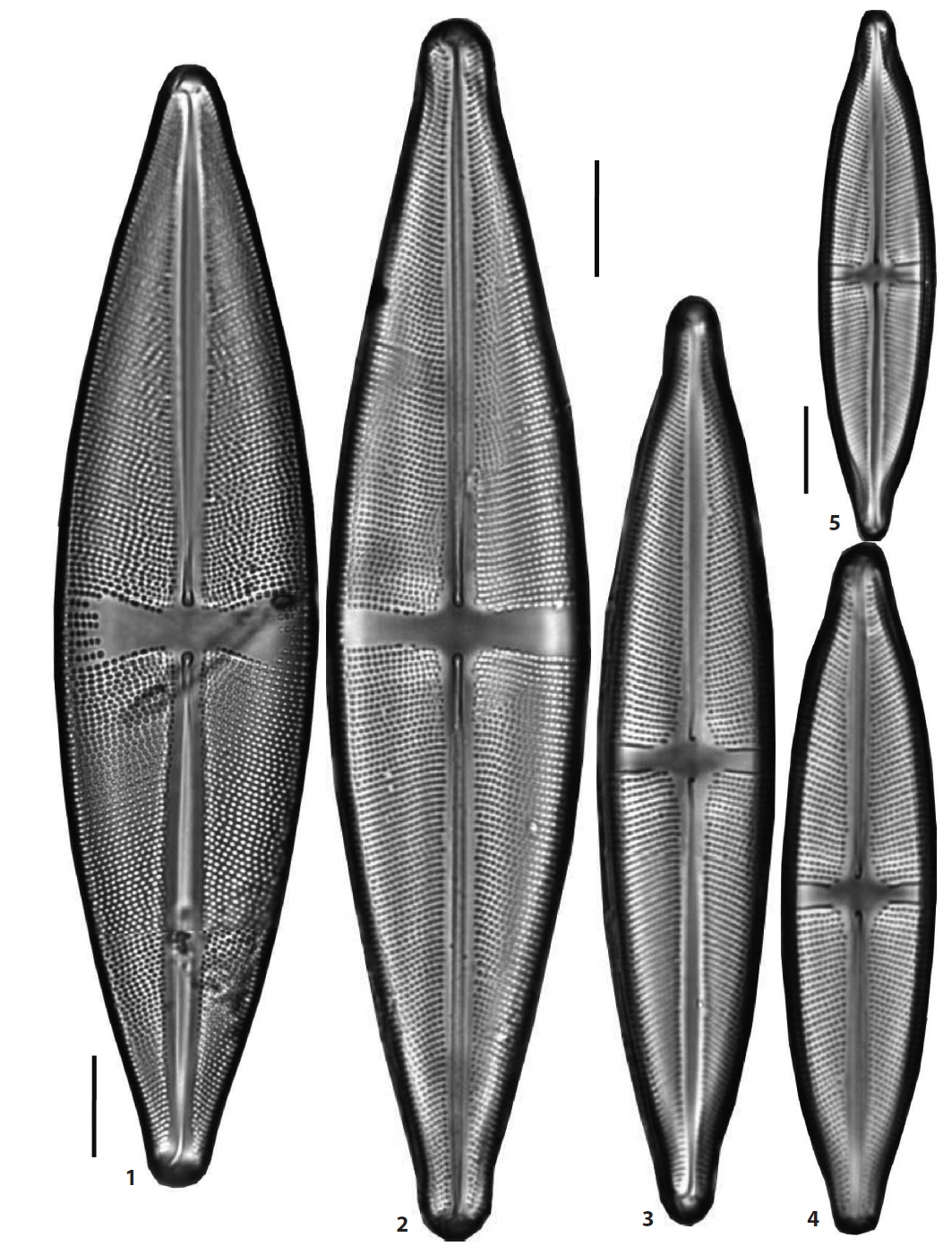
(6) *Stauroneis elena Kulikovskiy, Lange-Bertalot & Witkowski 2010 (Pl. 3, Figs. 9 and 10) (Kulikovskiy et al. 2010, p. 59, pl. 92: 1, 2). Valves elliptical-lanceolate in outline, the ends of the valve protracted and broadly rostrate. The stauros in central area expanded towards the margins. Valves 121–133 μm in length, 26–26.5 μm in breadth and striae 17–18 rows in 10 μm. Occurring rarely in mountaion peatlands, YN Peatland of Daeam Mountain in Inje and WDJ Peatland of Jiri Mountain in Sancheong, and newly reported to Korea.
(7) *Stauroneis circumborealis Lange-Bertalot & Krammer 1999 (Pl. 6, Fig. 1) (Van de Vijver et al. 2004, p. 31, pl. 10: 1-4, pl. 11: 1-3). Valves broadly elliptic-lanceolate in outline, the ends of the valve slightly protracted and rounded. The central ends of the raphe strongly curved and the terminal pores of the raphe drop-like expanded. The hyaline stauros of the central area rectangular, strongly expanded towards the margins, and some shortened striae in the margins. Valves 101-111 μm in length, 24-25.5 μm in breadth, striae 17-18 rows in 10 μm, and puncta in a stria 14-16 in 10 μm. Occurring very rarely in a lowland wetland in Haman and newly reported to Korea. The striae of the valve in the local specimens are sparsely arranged, 13-16 rows in 10 μm, less than 18-20 rows in 10 μm in the original description.
(8) Stauroneis gracilior Reichardt 1995 (Pl. 2, Figs. 3 and 4) (Lange-Bertalot et al. 2003. pl. 55: 1-11, pl. 56: 1, 2; Van de Vijver et al. 2004. p. 37. pl. 76: 1-10, pl. 77: 1-4). Replaced synonym: Stauroneis anceps f. gracilis Rabenhorst 1864, p. 247 (Krammer and Lange-Bertalot 1986, pl. 87: 9). Valves typically lanceolate, the breadth of the valve sharply narrow towards the ends, the ends of the valve long protracted and rostrate to capitate. The stauros rectangular and not expanded towards the ends. Valves 51–59 μm in length, 10–10.5 μm in breadth, and striae very fine and unresolved under light microscopy. Stauroneis gracilior Reichardt, S. schroederi Hustedt and S. subgracilior Lange-Bertalot et al., are closely related in the valve morphology, and make a group of species within the genus (Van de Vijver et al. 2004). Very rare in Dongsong Reservoir in Cheolwon.
(9) Stauroneis gracilis Ehrenberg 1843 (Pl. 2, Figs. 7 and 8) (Van de Vijver et al. 2004, p. 38, pl. 17: 1–4, pl. 18: 1–4, pl. 19: 1–4, pl. 20: 1–4). Valves lanceolate to somewhat linear-lanceolate in outline, the ends of the valve slightly protracted and broadly subrostrate. The stauros in central area weakly expanded towards the margins. Valves 66–100 μm in length, 14–17 μm in breadth and striae 18–19 rows in 10 μm. In a bibliotheca of genus Stauroneis written by Van de Vijver et al. (2004), S. gracilis and S. subgracilis were closely related in the valve morphology, and it was hard to discriminate two species each other. The dimensions or breadth of the valve and the density of striae are used to differentiate the species, and the valve of S. gracilis is wider than 14 μm in breadth and striae are less than 19 rows in 10 μm. Frequent in an unnamed peatland in DGR Pasture in Pyeongchang.
(10) *Stauroneis gracillima Hustedt 1943 (Pl. 3, Figs. 1–6) (Krammer and Lange-Bertalot 1986, p. 248, pl. 90: 28–30; Simonsen 1987, p. 311, pl. 469: 7–10). Valves linear with parallel or slightly convex margins, the ends of the valve protracted and capitate. The stauros in central area extended to the margins. Valves 10–14 μm in length, 2.5–3 μm in breadth and striae unresolved in light microscopy. Widely distributed in mountain peatlands and oligotrophic freshwaters in Korea, and newly reported to Korea.
(11) *Stauroneis heinii Lange-Bertalot & Krammer 1999 (Pl. 3, Figs. 7 and 8) (Van de Vijver et al. 2004, p. 40, pl. 1: 1). Valves lanceolate in outline, the ends of the valve slightly protracted and broadly subrostrate. The stauros in central area rectangular or weakly expanded towards the margins. Valves 99.5–168 μm in length, 20–33.5 μm in breadth and striae 16–17 rows in 10 μm. Frequently occurring in mountain wetlands, MJC3 Peatland in Ulsan and an unnamed peatland of DGR Pasture in Pyeongchang, and newly reported to Korea.
(12) *Stauroneis jarensis Lange-Bertalot, Cavacini, Tagliaventi & Alfinito 2003 (Pl. 1, Figs. 9 and 10) (Van de Vijver et al. 2004, p. 44, pl. 31: 1–10, pl. 32: 1–3, pl. 33: 1–6). Valves lanceolate to linear-lanceolate in outline, the ends of the valve protracted and rostrate. The stauros in central area expanded towards the margins. Valves 45–59.5 μm in length, 9–12 μm in breadth and striae 23–24 rows in 10 μm. Rare in YN Peatland of Daeam Mountain in Inje, and newly reported to Korea.
(13) Stauroneis kriegeri Patrick 1945 (Pl. 4, Figs. 1–5; Pl. 5, Fig. 11) (Van de Vijver et al. 2004, p. 45, pl. 59: 1–17). Valves linear with parallel or slightly convex margins, the ends of the valve broadly protracted and more or less capitate. The stauros in central rectangular extending to the margins. Valves 19–25 μm in length, 4–5 μm in breadth and striae unresolved in light microscopy. Widely distributed in mountain peatlands and some oligotrophic freshwaters, and more frequent in two mountain wetlands, DSA Peatland near Jeonjok Mountain in Yangsan and JD Peatland of Jang Island in Sinan.
(14) *Stauroneis kuelbsii Lange-Bertalot 2004 (Pl. 6. Figs. 3–5) (Van de Vijver et al. 2004, p. 45, pl. 40: 17). Valves lanceolate to linear-lanceolate in outine, the ends of the valve more or less abruptly protracted, subrostrate or subcapitate. The central ends of the valve curved. The hyaline stauros of the central area rectangular and slightly expanded towards the margins. Valves 59-80 μm in length, 13-16 μm in breadth, and striae 16-17 rows in 10 μm. Occurring rarely in lowland wetlands in Haman and newly reported to Korea.
(15) Stauroneis nobilis Schumann 1867 (Pl. 4, Fig. 7) (Krammer and Lange-Bertalot 1986, p. 242, pl. 87: 1, 2). Valves rhombic-lanceolate in outline, the ends of the valve long protracted and capitate. The stauros in central area rectangular and narrow towards the margins. Valves 80–108 μm in length, 16.5–25 μm in breadth, striae 16–17 rows in central area, but 19–21 rows in 10 μm in the terminal ends. Occurring frequently in Korean freshwaters, the lowland wetlands in Haman (Chung and Noh 1987), in Gwang River of Uljin (Lee et al. 1994), and recently rare in Dongsong Reservoir in Cheolwon.
(16) *Stauroneis prominula (Grunow) Hustedt 1959 (Pl. 5, Figs. 1 and 2) (Krammer and Lange-Bertalot 1986, p. 247, pl. 90: 16–20). Valves linear with slightly triundulate margins, the ends of the valve narrow protracted and rostrate, and pseudosepta developed. Axial area narrow. Axial area narrow and the stauros in the central area rectangular, extending to the margins of the valve. Valves 27–30 μm in length, 4–5 μm in breadth, and striae fine and unresolved in light microscopy. The species was found in two wetlands, JD Peatland of Jang Island in Sinan, and OGG Peatland of Jiri Mountain in Sancheon and is new to Korea.
(17) *Stauroneis pseudosmithii Van de Vijver & Lange-Bertalot 2004 (Pl. 5, Figs. 4–7) (Van de Vijver et al. 2004, p. 58, pl. 103: 1–11, pl. 104: 1–14). Valves narrow linear-lanceolate with triundulate margins, the ends of the valve narrow protracted and rostrate, and pseudosepta developed near the ends. Axial area narrow and the stauros in the central area rectangular, extending to the margins of the valve. Axial area narrow. Valves 27–30 μm in length, 5–5.5 μm in breadth and striae fine and unresolved in light microscopy. Very rare in a small pool of Baekyang Mountain in Busan and JD Peatland of Jang Island in Sinan, and recently occurring in mosses of Daebong Reservoir in Changnyeong, and newly reported to Korea.
(18) *Stauroneis reichardtii Lange-Bertalot, Cavacini, Tagliaventi & Alfinito 2003 (Pl. 1, Figs. 1–3) (Lange-Bertalot et al. 2003, p. 142, pl. 36: 1–8, pl. 37: 1–14, pl. 61: 6, 7; Van de Vijver et al. 2004, p. 60, pl. 54: 6–12, pl. 55: 1–13, pl. 56: 1–5). Valves linear to slightly linear-lanceolate, the ends of the valve protracted and capitate. The stauros of the central area almost rectangular, hardly expanded towards the ends. Valves 41–48 μm in length, 8–9 μm in breadth and striae 23–26 rows in 10 μm. Stauroneis reichardtii is closely similar to S. acidoclinatopsis Van de Vijver & Lange-Bertalot, and the former has more lanceolate outline of the valve and the bigger dimensions. It was found in some mountain peatlands, especially in DGR peatland in Pyeongchang and is new to Korea.
(19) *Stauroneis respectabilis Lange-Bertalot, Cavacini, Tagliaventi & Alfinito 2003 (Pl. 6, Fig. 2) (Lange-Bertalot et al. 2003, p. 144, pl. 31: 3, 4, pl. 32: 1-5, pl. 33: 1, 2). Valves lanceolate in outline, the ends of the valve weakly protracted or not protracted. The central ends of the raphe almost straight and the central pores of the raphe expanded. The hyaline stauros of the central area rectangular and more or less expanded towards the margins. Valves 102-133 μm in length, 21.5-25.3 μm in breadth, striae 16 rows in 10 μm, and puncta in a stria 16-18 in 10 μm. Occurring very rarely in lowland wetlands in Haman and newly reported to Korea. This species is very close to S. phoenicenteron (Nitzsch) Ehrenberg sensu stricto in the valve morphology, but differs in the dimensions, the lower valve length and breadth. S. phoenicenteron has large dimensions in valve, larger than 150 μm in length and 28 μm in breadth, though it has not been observed in this study.
(20) Stauroneis sagitta Cleve 1881 (Pl. 5, Figs. 9 and 10) (Van de Vijver et al. 2004, p. 63, pl. 101: 1–12, pl. 102: 1–4). Synonym: Stauroneis smithii var. sagitta (Cleve) Hustedt 1959 (Hustedt 1959, p. 810, f. 1158). Valves elliptical-lanceolate to rhombic-lanceolate with triundulate and convex margins, the ends of the valve narrow protracted and rostrate, and pseudosepta developed near the ends. Axial area narrow and the stauros in the central area rectangular, extending to the margins of the valve. Valves 31–41 μm in length, 8–9 μm in breadth and striae 23–24 rows in 10 μm. Occurring only in a lake, Dongsong Reservoir in Cheolwon.
(21) Stauroneis siberica (Grunow) Lange-Bertalot & Krammer in Lange-Bertalot and Metzeltin 1996 (Pl. 1, Figs. 11–13) (Lange-Bertalot and Metzeltin 1996, p. 104, pl. 35: 1, 2, pl. 104: 43, 44. Bahls 2011b). Basionym: Stauroneis anceps var. siberica Grunow in Cleve and Grunow 1880. Valves elliptical-lanceolate in outline, the ends of the valve protracted and rostrate to subcapitate. Axial area narrow. The stauros in central area narrow toward valve margins and areolae bordering the stauros coarse. Valves 55.5–62 μm in length, 13 μm in breadth and striae very fine, 26–27 rows in 10 μm. Transapical striae are very fine. This species was reported in the Kwang River in Uljin (Lee et al. 1994) and was rare in a pool of Odo Mountain in Hapcheon in this study.
(22) Stauroneis smithii Grunow 1860 (Pl. 5, Fig. 3) (Hustedt 1959, p. 810, f. 1157a–c). Valves lanceolate with triundulate and convex margins, the ends of the valve narrow protracted and rostrate, and pseudosepta developed near the ends. Axial area narrow and the stauros in the central area rectangular, extending to the margins of the valve. Valves 25 μm in length, 5.5 μm in breadth, and striae fine and unresolved in light microscopy. Very rare in the JD Peatland of Jang Island in Sinan.
(23) *Stauroneis sonya Kulkovskiy, Lange-Bertalot & Dorofeyuke 2010 (Pl. 4, Figs. 8 and 9) (Kulikovskiy et al. 2010, p. 62, pl. 93: 1–3, pl. 94: 1–3). Valves lanceolate to elliptical-lanceolate in outline, the ends of the valve not protracted and obtusely rounded. Axial area wide, the stauros in the central area rectangular, expanding towards the margins, and irregularly shortened striae in the margins. The central ends of the raphe reversely curved. Valves 121–142 μm in length, 26.5–34.5 μm in breadth, and striae 15–17 rows in 10 μm. Occurring more frequently in a mountain peatland of Jang Island in Sinan and an unnamed peatland of DGR Pasture in Pyeongchang and newly reported to Korea.
(24) *Stauroneis staurolineata var. japonica Kobayasi & Ando 1978 (Pl. 5, Figs. 12 and 13) (Kobayasi and Ando 1978, p. 15–16, pl. 2: 18–20). Valves lanceolate in outline, the ends of the valve not protracted and acute. The stauros in central area rectangular extending to the margins. Valves 99–114.5 μm in length, 16–18 μm in breadth and striae 15–18 rows in 10 μm. Only occurring in TH Peatland of Halla Mountain in Jeju Island and newly reported to Korea.
(25) *Stauroneis subgracilis Lange-Bertalot & Krammer in Lange-Bertalot and Genkal 1999 (Pl. 2, Figs. 5 and 6) (Van de Vijver et al. 2004, p. 71, pl. 27: 1–6, pl. 28: 1–6, pl. 29: 1–6). Valves lanceolate to elliptical-lanceolate in outline, the ends of the valve weakly protracted and broadly subrostrate. The stauros in central area rectangular extending to the margins. Valves 43–71 μm in length, 11–14 μm in breadth and striae 20–24 rows in 10 μm. This species is most similar in morphology of S. pergracilis Van de Vijver & Lange-Bertalot but distinguished by the lanceolate form of the valve rather than the linear to linear-lanceolate form. Rare in mountain wetlands, DJ Peatland of Yeongchuk Mountain in Yangsan and SMBD Peatland of Halla Mountain in Jeju Island, and newly reported to Korea.
Stauroneis diatoms occurred in most samples collected from peatlands and oligotrophic bodies of water, whereas they were scarcely found in eutrophic or polluted habitats. The relative abundance of Stauroneis diatoms in peatlands ranged from 0.1% to 3.4% of all diatom frustules, while two sites showed an exceptional abundance with about 7.6% and 10%, respectively. It was found that individual peatlands are typically inhabited by 2–6 Stauroneis species. On the other hands, two sampling areas, the mountain peatland of Jang Island in Sinan and an unnamed peatland of DGR Pasture in Pyeongchang showed high species richness of genus Staurones. The influence of pH and alkalinity on the abundance and richness of Stauroneis diatoms was evident, despite the relatively low abundance of this genus’s diatoms. More strongly acidic areas are characterized by low abundance and richness.
Throughout the whole sampling area, the most frequent and abundant species in the peatlands were S. kriegeri Patrick in the small forms and S. elena Kulikovskiy et al. in the large forms. Many species occurred in only one or two localities, including S. sonya Kulikovskiy et al., S. elena Kulikovskiy et al., S. ancepsopsis Lange-Bertalot et al. and S. gracilior Reichardt in the large forms, and S. smithii and its infraspecific taxa in the small forms.
This survey of the Staurones diatoms was restricted to the mountain peatlands, the headwaters of the streams, upstream waters, pools and reservoirs, and oligotrophic waters. Through this survey, it was evident that Stauroneis diatoms prefer acidic or oligotrophic freshwaters. Stauroneis was scarce in rivers and lakes, while a variety of Stauroneis taxa were found in the mountain peatlands or oligotrophic habitats.
Small forms of Stauroneis diatoms, Stauroneis gracillima (Grunow) Hustedt and S. kriegeri Patrick, were more widely distributed over the sampling area than large forms, as large forms tend to be discretely dispersed. Large-sized taxa were found to be restricted to only one or sometimes two areas or one island, for example, S. ancepsopsis Lange-Bertalot et al. only in a rice paddy, S. staurolineata var. japonica Kobayasi & Ando in TH Peatland of Halla mountain, and S. sonya Kulkovskiy, Lange-Bertalot & Dorofeyuke in JD peatland of Jang Island and DGR Pasture in Pyeongchang. The high degree of endemism of genus Stauroneis is shown in the Antarctic regions and northern Rocky Mountains (Van de Vijver et al. 2005, Bahls 2012).
Peatlands in mountainous area are likely a kind of isolated and undisturbed habitats, differing from the homogeneous surrounding areas. A large number of Stauroneis taxa have also been newly discovered in Antarctica and the Arctic, the Rocky Mountains, Sardinia and tropical regions (Lange-Bertalot et al. 2003, Van de Vijver et al. 2004, Metzeltin and Lange-Bertlaot 2007, Bahls 2010). The local endemics of Stauroneis species can be applied to this work. In Korea, a total of 22 taxa have been reported from a variety of freshwaters according to Lee et al. (1995) and Lee (1997). They accounted for 12 taxa mainly S. anceps Ehremberg sensu lato and S. phoenicenteron (Nitzsch) Ehrenberg sensu lato and their infraspecies, variety or forma. The two species of Stauroneis are well known to be widely distributed over the freshwaters. However, S. anceps and S. phenicenteron sensu lato taxon have been revised on the basis of the narrow species concept, thus being separated from the existing taxa, with a number of new species described in the Antarctic regions, Sardinia in the Mediterranean Sea and some other regions (Lange-Bertalot et al. 2003, Van de Vijver et al. 2004). In the current study, of the twenty five species are reported, sixteen are described for the first time in Korea.
Regardless of the endemics of Stauroneis diatoms, they are less abundant and diverse in particular various habitats. The algal assemblages in peatlands are largely dominated by diatoms and Stauroneis diatoms that are large in size compared with other diatoms. The largeness of Staurones can be correlated with their biomass. Species diversity has significant correlation with algal biomass. The significant negative linear correlation between largeness and algal biomass exists and algal biomass is dominated by the smaller diatoms (Eloranta 1993). Besides of the genus Stauroneis, other species-rich genera are Pinnularia, Eunotia, Neidium, and Frustulia, and the mountain peatlands support the abundance and species diversity of these diatoms. Thus, the protection of the habitat for the diatom biodiversity should be very important.