The genus Cryptonemia was established by Agardh (1842) based on Cryptonemia lactuca (C. Agardh) J. Agardh, which is currently regarded as a taxonomic synonym of the type species Cryptonemia lomation (Bertoloni) J. Agardh (1851) from Italy (Guiry and Guiry 2014). Cryptonemia lomation has the following characteristics: dichotomously branched or palmately lobed blade with an evanescent midrib at the base and a perennial stalk, and entwined filaments with highly refractive stellate cells in the medulla (Irvine 1983). Cryptonemia lomation appears to be closely related to Atlantic Cryptonemia seminervis (C. Ag.) J. Agardh (1846) (Athanasiadis 1987). Cryptonemia seminervis was originally described by Agardh (1846) based on Atlantic material collected from Cádiz, Spain, and it can be distinguished from other species that have a similar habit by the thickening (evanescent midrib) at the base of the blade and the stellate cells in the medulla (Irvine 1983). Dixon (1961) recognized the entity collected in Herm (British Channel Isles) as C. lactuca, but Irvine (1983) suggested that this taxon was conspecific with C. seminervis.
Forty-three Cryptonemia species are currently recognized worldwide; they occur in warm temperate to tropical regions of the Atlantic and Pacific Oceans (Guiry and Guiry 2014). Six species of Cryptonemia have been reported in Korea and Japan (Kang 1966, Yoshida 1998, Lee et al. 2005, Kim et al. 2012b). Among these, only Cryptonemia rotunda (Okamura) Kawaguchi has been reported from both countries (Kim et al. 2012b). Cryptonemia rotunda was recently transferred to the new genus Galene (D’Archino et al. 2014). Cryptonemia luxurians (C. Agardh) J. Agardh (1851) was first collected from Brazil, but the name is a taxonomic synonym of C. seminervis (Price et al. 1986, John et al. 2004). It has been reported from many national waters, including those of Japan and China, which are far from the type locality in Brazil (Silva et al. 1996, Yoshida 1998, Xia 2004). Asian C. luxurians specimens have a small discoid holdfast, a short stipe in the basal region, a conspicuous midrib through to the mid part of the thallus, and dichotomous branching with marginal leaflets (Yoshida 1998, Xia 2004).
Three Cryptonemia species have been listed in Korean literature: C. lactuca reported by Lee et al. (2005), C. tuniformis rarely collected by Kang (1966), and C. rotunda recently identified by Kim et al. (2012b). Cryptonemia lactuca was collected for the first time in Korea as summer beach cast. Specimens had a reniform, palmately lobed, and strongly undulate blade with ribs, which were proximally prominent but divided and vanished distally (Lee et al. 2005). Our recent collections from Jeju Island, Korea, contained specimens provisionally assigned to Cryptonemia; however, the specific identity of the specimens was not determined initially because the genus has not been studied in detail in Korea. In addition, several synonymies of the species in Cryptonemia have been suggested due to the difficulty of identification caused by simple foliose blades (Guiry and Guiry 2014).
Molecular taxonomy is especially useful for species that are wide-ranging and have few clear-cut morphological diagnostic criteria (Kim et al. 2012a, Yang et al. 2013). We assembled a collection of specimens of (i) C. lactuca from the subtidal zone of Jeju Island, Korea, (ii) C. luxurians from intertidal zone of Chiba, Japan, and (iii) C. seminervis from the type locality in Spain. To define the taxonomic entity of three species, we analyzed the plastid large subunit of the ribulose-1,5-bisphosphate carboxylase/oxygenase (rbcL) gene and compared them to the several Cryptonemia rbcL sequences available in GenBank (http://www.ncbi.nlm.nih.gov/genbank).
Samples were collected subtidally by SCUBA diving to 25-m depth in the subtidal zone or gathered as intertidal beach cast at a range of locations in Korea and Japan (Table 1). A sample of C. seminervis was collected from the type locality, Cádiz, Spain. Specimens were mounted on herbarium sheets from which we removed two subsamples; one was preserved in 5% formalin-seawater for morphological observations, and the second was desiccated in silica gel for molecular analysis. Sections were obtained by hand or with the aid of a NK-101-II freezing microtome (Nippon Optical Works Co. Ltd, Tokyo, Japan). The sectioned preparations were stained with 1% aqueous aniline blue acidified with a drop of 1% HCl and mounted in 30% Karo corn syrup (ACH Foods, Summit, IL, USA). Photomicrographs were captured with a QIMAGING 1394 camera (QImaging, Surrey, BC, Canada) attached to a microscope (BX50; Olympus, Tokyo, Japan). All images were imported into the PhotoShop ver. 5.5 software (Adobe Systems Inc., San Jose, CA, USA) for plate assembly. The voucher specimens were deposited in the herbarium of Jeju National University (JNUB), Jeju, and the National Institute of Biological Resources (NIBR), Incheon, Korea.
Total genomic DNA samples were extracted using the DNeasy plant mini kit (Qiagen, Hilden, Germany) following the manufacturer’s instructions. For amplification and sequencing reaction, we used the following primer pairs: rbcLF7-rbcLR898 and rbcLF762-rbcLR1442 (Gavio and Fredericq 2002, Kim et al. 2010). PCR amplification was performed with a Swift MaxPro Thermal Cycler (Esco, Singapore) using AccuPower PCR Premix (Bioneer, Daejeon, Korea). PCR was performed with an initial denaturation step at 94℃ for 4 min, followed by 35 cycles of 1 min at 94℃, 1 min at 50℃, and 2 min at 72℃, with a final 7-min extension at 72℃. The PCR products were purified with an AccuPrep PCR Purification Kit (Bioneer) and then sequenced commercially by Macrogen (Seoul, Korea). Two electropherogram outputs from each sample were edited using Chromas ver. 1.45 software (Technelysium Pty Ltd., Helensvale, QLD, Australia).
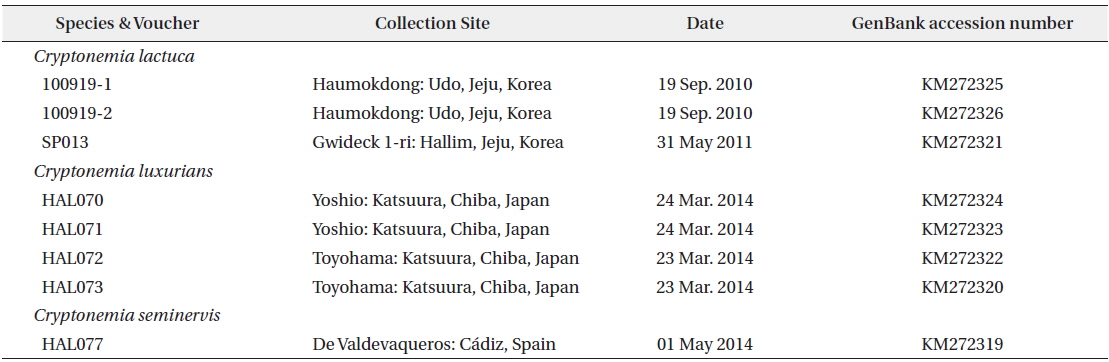
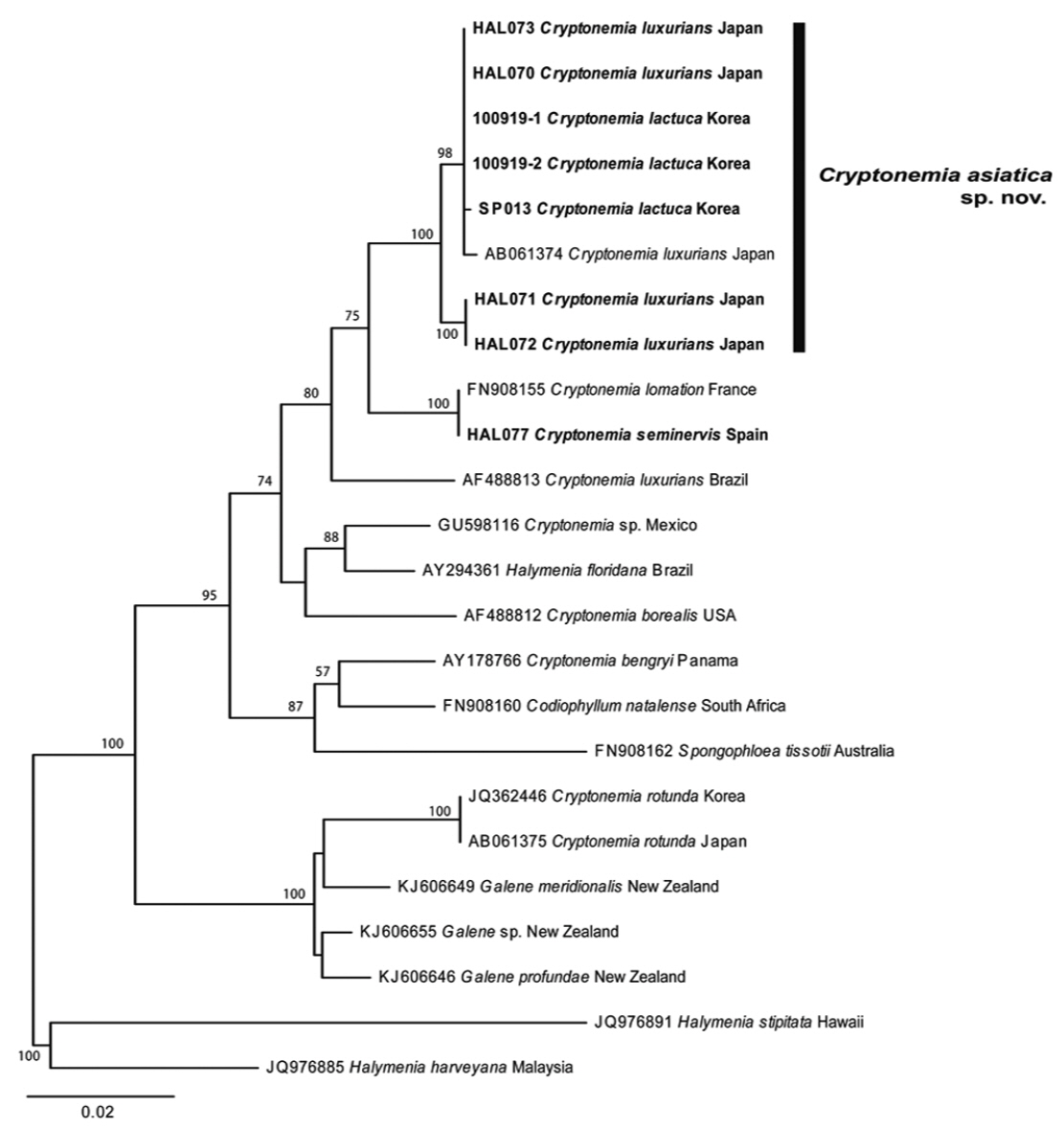
We collated a total of 24 rbcL sequences (8 new) with multiple-sequence alignment BioEdit software (Hall 1999) and aligned them visually. Collection site information and the GenBank accession numbers of the eight specimens are listed in Table 1. Plastid rbcL sequences of two Halymenia species (H. harveyana and H. stipitata) were included as outgroups. Phylogenetic analyses were performed using the ML (Maximum likelihood) routine in RAxML software (Stamatakis 2006). We assembled 200 independent tree inferences, and applied the “-#” (with default –I: automatically optimized Subtree PruningRegrafting rearrangement) and “-c” (25 distinct rate categories) software options to identify the best tree. We performed 1000 iterations with the same software and settings to generate bootstrap values for these phylogenies.
We aligned a total of 1,347 bp in 24 rbcL sequences from species of Cryptonemia, related genera, and two Halymenia species (as outgroups) (Fig. 1). Three specimens of C. lactuca from Korea and five of C. luxurians from Japan fell in a single clade with 0.7% divergence (100% bootstrap value). Members of the clade clearly differed from C. luxurians collected from the type locality in Brazil (AF488813) with 3.3–3.5% interspecific sequence divergence. The sequences for C. seminervis specimens collected from the type locality in Spain (HAL077) were identical to those of specimens of C. lomation (generic type) from France (FN908155). Our specimens from Korea and Japan formed a sister group to the specimens of C. lomation from France and C. seminervis from Spain with 2.1–2.3% interspecific divergence in 75% bootstrap value. The sequence divergence within the Cryptonemia clade that included the type species of the genus, C. lomation, ranged from 1.9% between Cryptonemia sp. (GU598116) and Halymenia floridana (AY294361) to 5.9% between H. floridana (AY294361) and Spongophloea tissotii (FN908162). The recently established related genus Galene differed by 5.4–6.5% from our Asian specimens of Cryptonemia. The genus Cryptonemia was not a monophyletic group in our phylogenetic tree (Fig. 1) because of the presence of Halymenia floridana from Brazil (AY294361) and Cryptonemia bengryi from Panama (AY178766). On the basis of our molecular analyses, we propose Cryptonemia asiatica sp. nov. from Korea and Japan, which is included in recent collections that were previously assigned to C. lactuca in Korea and C. luxurians in Japan.
[Fig. 2.] Cryptonemia asiatica M.Y. Yang et M.S. Kim, sp. nov. (a) Holotype specimen (JNUB-100919-2, vegetative plant, 19 September 2010, Udo in Jeju Island) deposited in the Herbarium of Jeju National University (JNUB), Jeju, Korea. (b) Vegetative plant with young blades collected from Gwideok in Jeju Island (SP013, 31 May 2011). (c) Vegetative plant collected from Chiba in Japan (HAL071, 24 March 2014). (d) Midrib divided several times in the blade. (e) Globular or angular cells on the surface of thallus. (f) Cross section of terete stipe at the base of plant. (g) Cross section of middle part of stipe becoming flattened. (h) Cross section of blade with midrib. (i, j) Cross sections of blade showing refractive ganglionic cells (arrowheads). Scale bars represent 5 cm (a-c), 50 mm (d), 50 μm (e, i, j), 200 μm (f, g), and 500 μm (h).
Description: Thallus comprising foliose blades, solitary, epilithic, medium red to red-brown fading greenish-yellow, membranous and cartilaginous, undulate margins, terete stipe becoming flattened and alate, holdfast discoid to conical, irregularly branched with flabellate blade, prominent ribs in the mid to lower portion of blade, vanishing in the distal portion, sometimes dividing into 2–5 branches, rounded without midrib and smooth margins in young blades, globular or angular cells at the blade surface, cortex 4–6 cells thick, relatively narrow outer cells, inner cells ovoid, medulla of filaments longitudinally elongated, with prominent refractive ganglionic cells. Gametophyte and tetrasporophyte unknown.
Holotype: JNUB-100919-2, vegetative plant, collected on 19 September 2010 (Fig. 2a).
Isotypes: JNUB-100919-1, 100919-3; NIBRAL0000143 239-40.
Type locality: Udo, Jeju Island, Korea (33°33´28.01˝ N, 126°45´29.77˝ E).
Etymology: The specific epithet refers to Asia (Korean and Japanese collection sites were off the Asian coastline).
Korean name: 상추말
Distribution: Jeju Island, Korea and the Pacific coast of Honshu Island, Japan.
Habitat: Cryptonemia asiatica grows in the subtidal zone to 25-m depth and is attached to bedrock.
Other specimens examined. JN-SP013 (vegetative thallus, Gwideok, Jeju Island, Korea collected on 31 May 2011); JN-HAL070-073 (vegetative thallus, Katsuura, Chiba Prefecture, Japan collected on 24 March 2014).
Morphology: Plants are solitary, epilithic, and growing up to 35 cm tall. Thalli have the clearly visible stems with irregular branches, medium red to red-brown, fading to greenish-yellow, membranous and cartilaginous foliose blades with undulate margins. Holdfasts are discoid to conical, and stipes are terete (Fig. 2f-2h) becoming flattened and alate, extending into broad leaf-like blades. Blades are flabellate, irregularly linear-oblong, 1–2 cm broad, 2–7 cm long, 60–150 µm thick (Fig. 2a-2c). Young blades are rounded without midrib, more or less smooth margins (Fig. 2b). Blades have prominent ribs in lower to mid portions, ribs are gradually vanished distally. Midribs of blades are sometimes branched 2–5 times (Fig. 2d). Blade surface cells are globular or angular (Fig. 2e). Tissue is comprised of cortex and medulla (Fig. 2i). Cortex is compact with 4–6 layers of cells. Outer cortical cells are relatively narrow, ellipsoid, 4–5 µm in diameter. Cortical inner cells are ovoid, loosely aggregated, 8–15 µm in diameter. Medulla is comprised of longitudinally elongated filaments (Fig. 2j) and prominent refractive ganglionic cells (Fig. 2i).
Cryptonemia lactuca was first reported in Korea by Lee et al. (2005), and C. luxurians was recorded as a component of the Japanese flora by Okamura (1936) and Tanaka (1963). Lee et al. (2005) indicated that two species were differentiated by the shapes of stipes and blades: only C. luxurians had sublinear bladelets on the midrib (Yoshida 1998). Although two species are different from one another in morphology, their taxonomic statuses are debatable. Our surveys of the subtidal area in Jeju Island, Korea, have been continued to discover the marine red macroalgal species diversity. Consequently, we collected samples of C. lactuca from Korea and also collected samples of C. luxurians from Japan that was cast ashore. Our molecular analyses have now revealed the presence of a new species in a separate clade containing Korean C. lactuca and Japanese C. luxurians specimens with 100% bootstrap support. Specimens of the new species are clearly distinguished from other species of Cryptonemia, including the generic type, C. lomation from France, and C. luxurians from Brazil, the type locality of this species. The binomial C. lactuca is currently regarded as a taxonomic synonym of C. lomation; we therefore assign the new name Cryptonemia asiatica to a cryptic species occurring in Korea and Japan.
According to Guiry and Irvine (1974), Cryptonemia is characterized by medullary filaments, cortical filaments that are three or four cells in length, and secondary thickening at the bases of blades that sometimes extends distally as a midrib. The new species Cryptonemia asiatica is epilithic and occurs to 25 m depth in the subtidal zone. It has the following characteristics: prominent midribs in the mid to lower portions of the blade, a cortex 4–6 cells thick, and longitudinally elongated filaments in the medulla. Cryptonemia asiatica resembles C. luxurians from Brazil in midrib morphology, a linear-oblong thallus shape, and a blade margin that is entirely undulate. However, C. luxurians from Brazil occurs intertidally, (Pinheiro-Joventino 1977), unlike the new species. Tetrasporangia of C. luxurians from the Atlantic are restricted to marginal leaflets (Taylor 1960), but in C. seminervis they are scattered over the frond (Guiry and Irvine 1974). Although we have no new information on tetrasporangial distribution in C. asiatica because fertile plants were not found in Korean and Japanese collecting sites in this study, Lee et al. (2005) reported that tetrasporangia were formed from cortical cells of thallus segments, and that small segments appeared to be special appendages for tetrasporangial formation.
We included a sample of C. seminervis from the Atlantic type locality in Cádiz, Spain; the sample was identical to C. lomation from France. Haroun et al. (1993) collected C. lomation from depths of 60–80 m and described thalli with small discoidal holdfasts, short terete stipes in the basal parts as evanescent midribs, and irregular margins with some proliferations. The binomial C. luxurians is currently regarded as a taxonomic synonym of C. seminervis (Dawes and Mathieson 2008). Our study showed that C. seminervis from Spain might be conspecific with C. lomation from France, an opinion shared by Irvine (1983) and Athanasiadis (1987). Cryptonemia luxurians from Brazil should be regarded as a separate species within the genus Cryptonemia.
D’Archino et al. (2014) established the new genus Galene, which is morphologically very similar to Cryptonemia in both vegetative and reproductive structure. Kawaguchi (1993) transferred Halymenia rotunda Okamura to Cryptonemia based on vegetative anatomy and the shape of the auxiliary ampulla. Kim et al. (2012b) showed that Korean specimens of C. rotunda are only distantly related to other species of Cryptonemia. In the present molecular analysis, our C. rotunda specimens from Korea and Japan fell together in a clade with the genus Galene with 100% bootstrap value. However, the genus Cryptonemia was not resolved as a monophyletic group in our plastid rbcL-based phylogenetic tree (Fig. 1). The generic type C. lomation fell in a clade with the new species C. asiatica, but with only 75% bootstrap support. The distant relationships between the clade of C. lomation/C. asiatica and their putative relatives raise questions concerning divergence pathways, which will be re-investigated through additional taxon sampling within the genus. Our present analysis has also clarified the distinction between C. seminervis from Spain and C. luxurians from Brazil with 2.7% sequence divergence, even though binomials have been suggested as a synonym by Price et al. (1986). In conclusion, specimens from Korea and Japan previously assigned to C. lactuca (Lee et al. 2005) and C. luxurians (Yoshida 1998) should be reassigned to Cryptonemia asiatica based on our molecular analysis of rbcL gene.