조류는 하천 및 호소 생태계의 1차 생산자로서 먹이사슬의 기반을 형성하여 생태계 내 에너지 순환에서 중요한 역할을 한다(Reynolds, 1984). 하천과 호소에서 조류는 식물플랑크톤을 비롯하여 부착 및 저서성 조류로 대표된다. 조류는 서식 환경의 물리·화학적 변화에 따라 민감하게 반응하여 시·공간적 분포 특성이 뚜렷하므로 환경오염의 지표성이 뛰어나 하천의 오염 지표종으로서 수환경의 변화를 모니터링하는데 중요하게 이용된다(Harper, 1992; Palmer, 1980).
우리나라와 같이 사계절이 뚜렷한 온대기후에 서식하는 식물플랑크톤은 크게 수온, 광도 등 계절적 변화에 따라 천이하며(Sommer et al., 1986), 남조류의 경우 겨울에는 휴면세포 혹은 휴면포자의 형태로 퇴적층에 침전되어 수온이 높아질 때 까지 휴지기(resting stage) 상태로 존재하게 된다(Kang et al., 2014). 이후 계절 변화로 수온이 상승하게 되면 퇴적층에 침전되어 있던 조류세포들이 휴지기를 마치고 영양세포로 성장하면서 표층으로 부유하여(Yamamoto and Nakahara, 2009), 식물플랑크톤 군집을 형성하고, 조건에 따라 대발생을 일으키기도 한다(Li et al., 2013). 이러한 이유로 퇴적층에서의 조류발생과 식물플랑크톤 군집형성과정은 수자원관리에 있어서 중요하지만, 현재까지는 많은 연구가 이루어지지 않고 있는 실정이다.
특히 북한강 녹조현상을 유발하는 대표 종으로 알려진
따라서 본 연구에서는 조류발생 및 이취미 문제가 발생했던 의암호 내 춘천하수처리장 방류구 부근과 공지천 하류의 퇴적층 2지점을 채집·배양하여, 퇴적층으로부터 발생되는 조류군집 발생특성 및 그에 의한 이취미물질 발생여부를 파악하고자 하였다.
퇴적층 배양실험에 사용된 시료는 2014년 4월 15일 의암호 내에 위치한 춘천하수처리장 방류지점(US)과 의암호 유입지천인 공지천의 하류(GS, 공지천교)에서 grab sampler를 사용하여 5~10 cm 깊이의 퇴적층을 채집하였다. 채집된 퇴적층은 유리재질의 배양조(길이60 cm × 폭33 cm × 높이40 cm)에 5±0.5 cm 높이로 퇴적층이 교란되지 않도록 약 10 L를 주입하였고, 배양조의 물은 현장수를 식물플랑크톤 네트(mesh 5 µm)로 여과하여 각각 60 L(수심 30 cm)를 주입하였다. 배양기간은 2014년 4월 16일부터 5월 16일까지 총 31일간 배양하였으며, 수온은 20±3℃, 광도는 40 µmol m−2s−1 (light:dark=16:8)의 조건으로 배양하였다. 시험기간 동안 퇴적층 배양조에 외부 영양원의 주입은 없었으며, 분석으로 감소되는 물의 추가는 없었고, 분석을 위한 시료 채수량은 최소화하였다.
퇴적층 배양에 따른 이화학적 환경요인, 식물플랑크톤 및 저서성 조류군집, 휴면포자(akinete), 이취미 물질을 확인하기 위하여 주2회 시료를 채집하여 분석하였으며, 배양시작 최초 5일은 1일 간격으로 분석하였다. 이화학적 환경요인은 수질 현장측정기(YSI 6600, USA)를 사용하여 수온, pH, DO, 전기전도도(EC) 및 탁도(Turbidity)를 측정하였고, 총질소(T-N), 총인(T-P), 질산성 질소(NO3-N) 및 인산염인(PO4-P)은 수질오염공정시험기준(MOE, 2010) 및 Standard method (APHA, 2005)에 준하여 분석하였다.
식물플랑크톤은 표층수를 채집하여 Lugol’s solution으로 고정하였고, 저서성 조류는 퇴적표층 2×2 cm2 면적을 100 mL 증류수에 희석하여 포르말린을 고정하였다. 식물플랑크톤과 저서성 조류의 분석은 Sedgwick-Rafter counting chamber를 사용하여 위상차 현미경(Nikon, Japan)하에서 100~1,000 배율로 개체수를 산출하였으며, 규조류는 Simonsen의 분류체계(Krammer and Lange-Bertalot, 1986, 1988, 1991a, 1991b)에 따라 정리하였고, 그 외 분류군은 Chung(1993), John et al. (2002), Hirose et al. (1977)을 참고하였다.
휴면포자의 분석은 퇴적표층을 2 cm 깊이로 채집, 습중량 1 g(w/w)을 취하여 증류수에 분산시켰다. 분산은 초음파기(WUC-A03, Korean)로 30초간 2회 처리하여 현탁액을 일정하게 분쇄하였다. 분산액은 90 µm, 60 µm Nylon재질의 체로 순차적으로 분리하였으며, 최종적으로 10 µm Nylon 체로 여과한 퇴적물 시료를 Panning methods(Matsuoka and Fukuyo, 2000)을 사용하여 휴면포자를 분리하였다. 분리된 휴면포자는 10 mL 부피로 농축하여 Sedgwick-Rafter counting chamber를 사용하여 위상차 현미경에서 100~400배율로 현존량을 산출하였다.
이취미 분석은 먹는물 수질감시항목 운영지침(MOE, 2011)에 따라 GC/MS(Bruker, 450GC/320MS)를 이용하여 HS-SPME 법으로 geosmin과 2-MIB(2-methyliso-borneol)를 분석하였다.
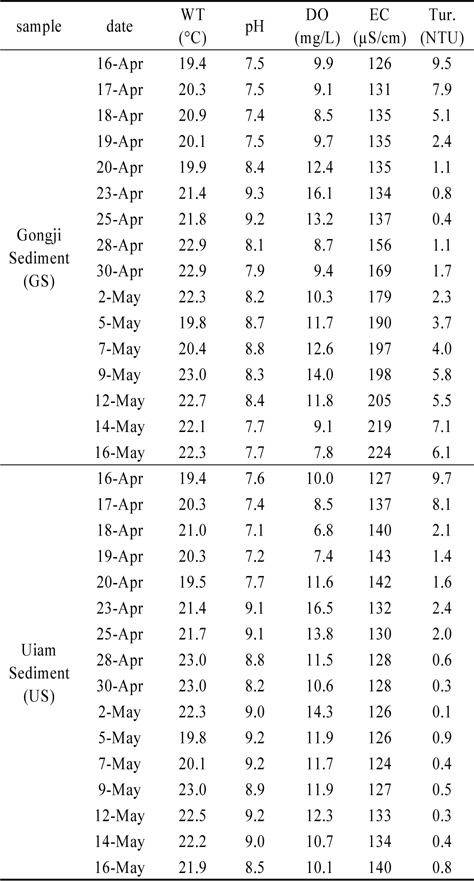
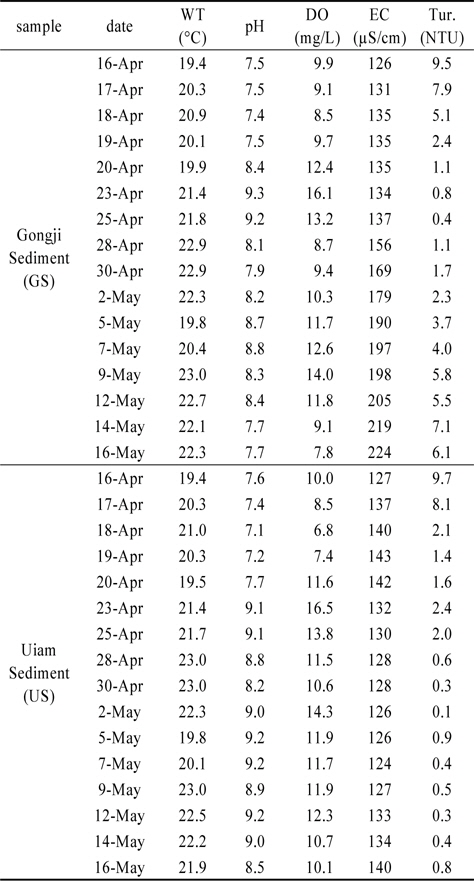
퇴적층 배양조의 일반 수질항목은 여과한 현장수를 주입 후 부유 퇴적층을 안정화시키기 위해 24시간 정치하였다(Table 1). 실험기간 동안 배양조의 수온은 20±3℃이었다.
[Table 1.] Variation of water quality at Gongji (GS) and Uiam (US) sediment by incubation period
Variation of water quality at Gongji (GS) and Uiam (US) sediment by incubation period
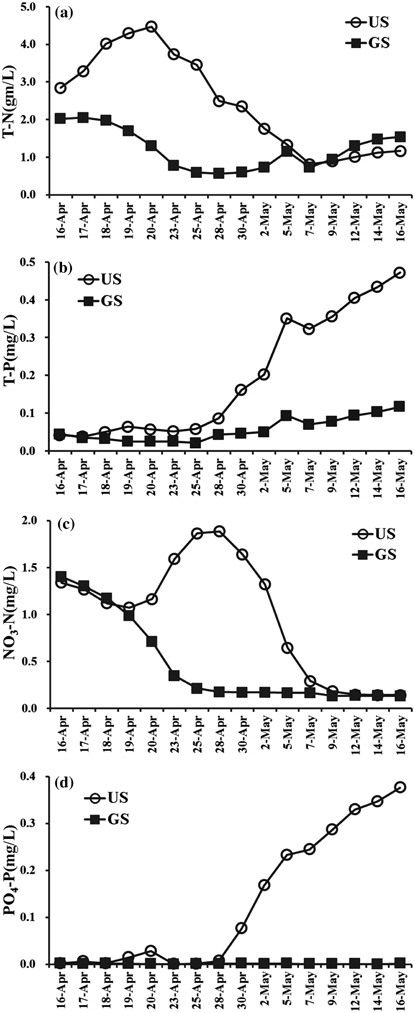
pH는 공지천퇴적층(GS)이 7.4~9.3 범위, 의암호퇴적층(US)이 7.1~9.2의 범위였으며, DO는 GS가 7.8~16.1 mg/L 범위, US가 6.8~16.5 mg/L 범위로 나타났다. GS의 경우 조류의 성장이 활발했던 4월 23일, 5월 7일 전후에 pH와 DO의 농도가 높았다. US는 4월 23일 가장 높았으며, 배양 종료시점까지 pH는 8.5이상, DO는 10 mg/L이상의 수준을 유지하였다. 이러한 pH와 DO농도 변화는 조류(algae)의 광합성에 따라 수중 용해된 중탄산염(HCO3−)을 흡수함으로써 pH가 상승하고, DO농도가 증가하는 기작에 의한 것으로 판단된다(Graham et al., 2009; Kim et al., 2009).
전기전도도는 GS가 126~224 µS/cm 범위, US가 124~143 µS/cm 범위로 나타났으며, GS는 배양이 진행됨에 따라 지속적으로 증가하였다. US는 배양기간 동안 큰 차이가 없이 평균 132 µS/cm 수준을 유지하였다. GS 전기전도도 농도가 4월 28일 급증하였으며, pH와 DO 감소 및 탁도가 급증한 이유는 퇴적층 표층에 형성된 저서성 남조류 매트가 배양조 표층으로 상승하여 퇴적층 유기물과 동시에 부상하였기 때문이다.
탁도의 경우 GS는 0.4~9.5 NTU, US는 0.1~9.7 NTU의 범위로 실험초기 부유되었던 유기물로 인해 4월 16일 두 수조에서 모두 가장 높게 나타났으며, 배양시작 후 3일까지 영향을 미쳤던 것으로 판단된다. US는 3일 이후 3 NTU이하로 유지되었으나, GS의 경우 배양 후기인 5월 9일 이후로 점차 증가하였다(Table 1).
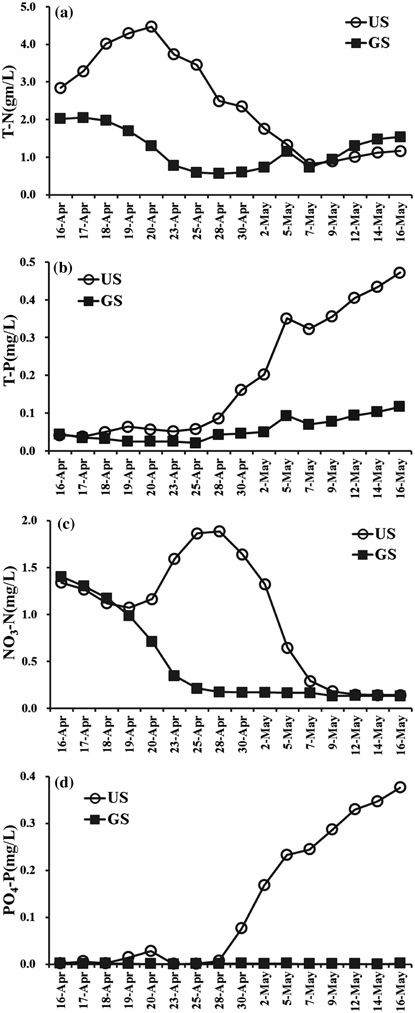
퇴적층 배양에 따른 영양염류 농도변화는 GS와 US가 뚜렷한 차이를 나타냈다(Fig. 1). 실험기간 중 총질소의 경우 GS는 0.566~2.024 mg/L, US는 0.813~4.468 mg/L 범위로 나타났다. US는 배양 초기 3일간 급격히 증가하여 4월 20일에 가장 높은 농도로 나타났으며, 5월 7일까지 지속적으로 감소하였다. GS는 배양 초기 3일간 농도의 변화가 없었으며, 4월 20일 이후 점차 감소하였고, 5월 5일 일시적으로 증가한 뒤 1 mg/L의 수준을 유지하였다. Fig. 1(c)와 같이 질산성 질소는 GS가 0.211~1.403 mg/L, US는 0.141~1.886 mg/L의 범위로서 GS는 배양 초기부터 4월 24일까지 급격히 감소하여 0.170 mg/L의 수준을 유지되었다. US는 배양 초기부터 4월 20일까지 감소하였다가 4월 28일에 농도가 가장 높았으며, 5월 7일까지 큰 폭으로 감소 후 GS와 같이 0.170 mg/L의 수준을 유지하였다.
GS 배양조의 경우 4월 17일부터 저서성 조류에 의해 매트형성이 시작되어 저층에서 기포가 발생되는 것이 관찰되어 매트가 부상한 4월 23일까지 조류 성장에 필요한 질소원을 섭취하여 질산성질소가 감소되는 것으로 판단된다. US의 경우에는 매트형성이 지연되어(4월 23일경) 질산성질소는 4월 28일부터 조류증식에 섭취되어 감소하여 5월 12일부터 0.13 mg/L 수준 이하로 유지되었다.
총인의 경우 GS는 0.021~0.117 mg/L, US는 0.038~0.472 mg/L의 범위로 US 배양조에서 높게 유지되었으며, 두 배양조 모두 4월 28일부터 증가하기 시작하여 실험종료 시점까지 지속적으로 증가하여 최대 0.472 mg/L을 나타냈다(Fig. 1(b)). 인산염인은 GS배양조에서 0.002 mg/L 이하로 지속되었으며, US에서는 4월 28일 이후 0.376 mg/L까지 지속적으로 증가하는 경향을 나타냈다(Fig. 1(d)). 이와 같이 인산염인 및 총인이 증가하는 원인은 Table 1에 나타난 바와같이 US배양조의 pH가 7.7(4월 20일)에서 9.1(4월 23일)로 높아짐에 따라 금속류와 착물로 형성된 인 용출이 일어났기 때문으로 판단된다. 퇴적물을 pH 6~9 변화시켰을 때 인 용출은 pH 9에서 급격히 증가하는 연구결과와(Jun and Park, 1989) 유사한 경향으로 판단된다.
GS와 US의 영양염의 농도는 Forsberg and Ryding (1980)의 기준으로 부영양(T-N: 0.6 mg/L 이상, T-P: 0.25 mg/L 이상)에서 과영양(T-N: 1.5 mg/L 이상, T-P: 0.1 mg/L 이상)의 영양단계(Trophic state)로 나타났으며, 퇴적층 채집 지점인 공지천교의 연평균 영양염류의 농도(T-N: 3.324 mg/L, T-P: 0.032 mg/L)와 유사한 수준인 것으로 나타났다(NIER, 2014).
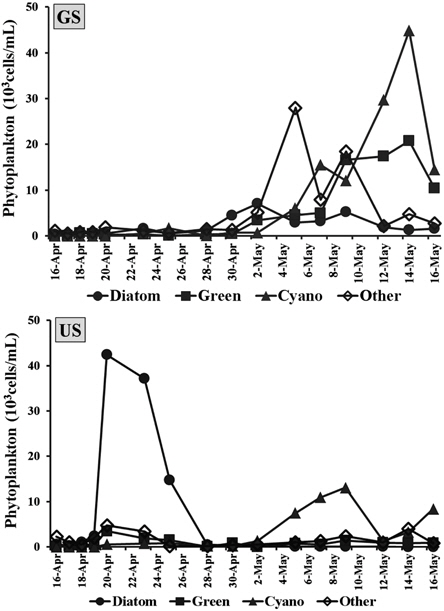
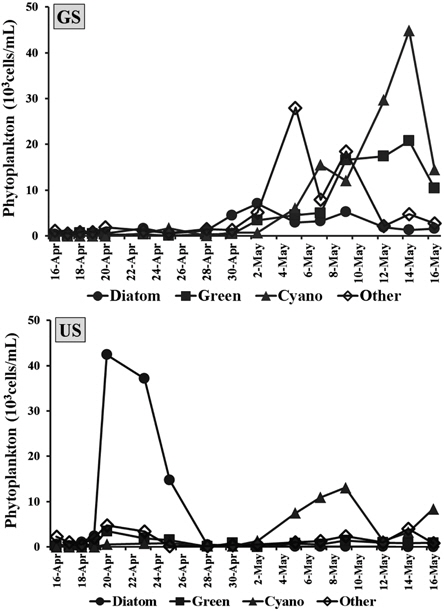
퇴적층 배양시 식물플랑크톤 군집은 GS배양조에서 총 45분류군의 식물플랑크톤이 출현하였으며, 녹조류 15분류군으로 가장 다양하게 출현하였고, 규조류 14분류군, 남조류 9분류군, 유글레나조류 및 갈색편모조류 등 그 외 조류가 7분류군 출현하였다. US배양조에서는 총 40분류군이 출현하여 녹조류 18분류군, 규조류 10분류군, 남조류 6분류군, 그 외 조류가 6분류군 출현하였다. Hašler et al. (2004)의 퇴적층 배양 연구에서도 49분류군이 확인되었고, 녹조류가 가장 많이 출현한 것으로 보고하고 있어 본 연구결과와 유사하였다.
GS배양조의 총식물플랑크톤 현존량은 0.9~71.8×103 cells/mL로서 배양 초기에는 낮았으나 4월 28일부터 현존량이 지속적으로 증가하면서 5월 14일에 최대 현존량을 나타냈다(Fig. 2). 배양 초기에는 갈색편모조류가 주로 출현하여 5월 4일에는
US배양조의 총식물플랑크톤 현존량은 0.8~51.2×103 cells/mL 범위였으며, 배양 5일째인 4월 20일 규조류
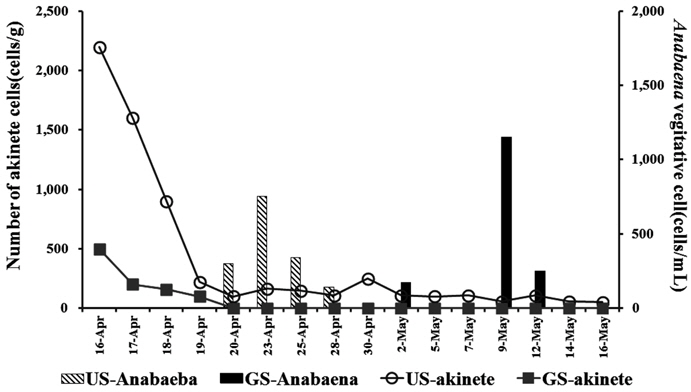
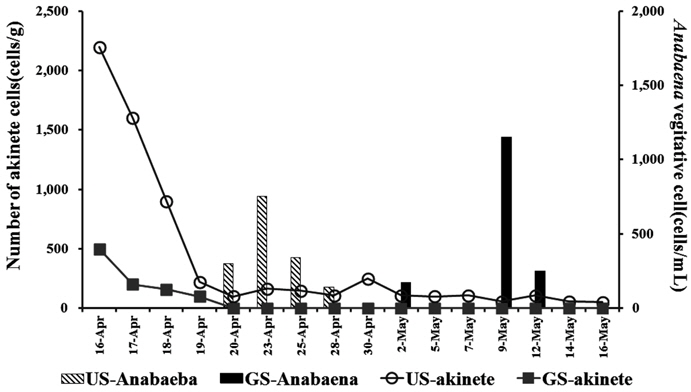
서식 환경이 열악하게 되면 남조류는 휴면기 세포인 휴면포자(akinete)를 만들며, 환경이 호전되면 다시 생존하게 된다. 퇴적층 표층에 존재하는 이러한 휴면포자 현존량은 잠재적인 남조류의 발생량이 되므로 의의를 가진다. Fig. 3과 같이 퇴적층 배양기간 동안 주기적으로 퇴적표층에 분포하는 휴면포자 현존량을 조사한 결과, 두 배양조 모두 배양초기에는 높았으며, 배양이 진행될수록 점차적으로 감소하였다. GS배양조의 휴면포자는 초기 500 cells/g(w/w)로 검출되어, 배양 2일째에 200 cells/g(w/w)로 감소하였고, 4월 20일 이후에는 관찰되지 않았다. US 배양조는 초기 2,200 cells/g(w/w)로서 GS보다 현존량이 높았으며, 배양 시작 후 급격히 감소하여, 4월 20일 이후에 배양 종료 시까지 120 cells/g(w/w)의 수준을 유지하였다(Fig. 3).
GS배양조의 휴면포자 검출 수준은 ‘유해조류 분포 및 생리·생태연구(Han River Watershed Management Committee, 2012)’에서 25~660 cells/g(w/w)의 범위로 나타난다고 보고한 것과 유사하였다. 또한 휴면포자의 감소경향은 석촌호수에서 3월~5월 사이 3,500 cells/g(w/w)로 분포하였으나 수온이 20℃ 이상으로 증가하는 5월 이후 233 cells/g(w/w)로 감소하는(Kim et al., 2005) 것과 유사하였다.
휴면포자의 발아(gemination)는 대상 분류군이나 조건에 따라 다르지만 20℃, 40 µmol m−2s−1의 조건에서 3일 동안 17~28%의 비율로 발아하는 것으로 알려져 있다(van Dok and Hart, 1997). 본 연구에서도 배양시작 3일간 휴면포자의 현존량이 10% 수준으로 감소하였으며, US의 경우
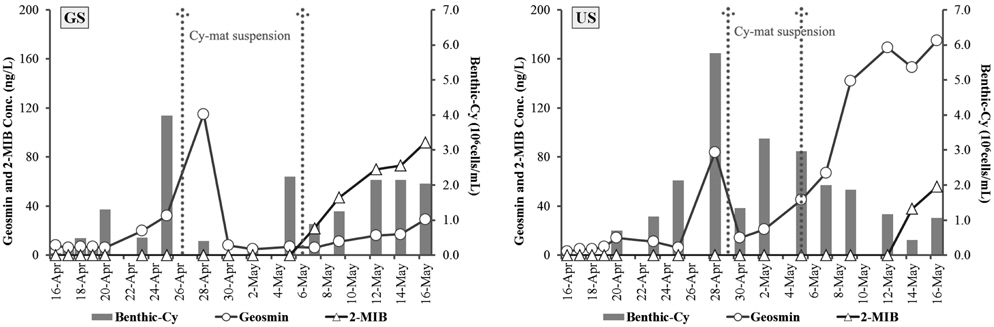
3.5. 이취미 물질(geosmin, 2-MIB)과 남조류 발생의 관계
수계에서 발생되는 이취미물질은 남조류의 성장과 밀접한 관련이 있으며, 플랑크톤성 남조류보다 저서성 남조류의 증식에 의해서도 영향을 받는다고 보고한다(Jüttner and Watson, 2007).
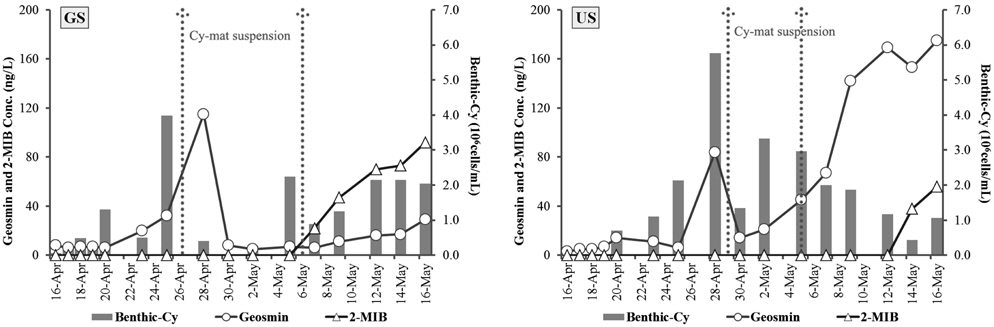
GS배양조의 경우 geosmin 5~115 ng/L의 범위로 4월 28일에 가장 높았으며, 비슷한 시기인 4월 25일에 저서성 남조류(benthic-cy)의 현존량은 3.96×106 cells/cm2로 가장 높았다. 퇴적표층에 형성된 남조류 매트가 표층으로 1차 부상하였고(4월 26일), 표층으로 부상한 매트는 4월 28일에 퇴적표층으로 다시 침강하였다. 저서성 남조류가 2.25×106 cells/cm2로 다시 증가한 후(5월 5일), 남조류 매트가 다시 표층으로 2차 부상하였다(5월 6일). 두 번째 남조매트의 부상 이후에는 geosmin이 아닌 2-MIB 농도가 22~92 ng/L의 범위로 증가하였으며(Fig. 4), 이 시기에는 부유성 남조류의 현존량도 44.85×103 cells/mL로 높았다(Fig. 2).
US배양조에서 geosmin은 3~175 ng/L의 범위로 5월 16일에 최대 농도로 나타났으며, 저서성남조류는 1차 매트 부상직전인 4월 28일에 5.77×106 cells/cm2로 가장 현존량이 높았다. 남조류 매트가 표층으로 1차 부상하였고(4월 29일), 5월 5일 남조류 매트의 2차 부상 이후, 2-MIB 농도가 증가하는 경향을 나타냈다. US배양조에서 geosmin 농도는 저서성 남조류가 가장 높은 시점과 증가 후 사멸하는 시기에 높았다(Fig. 4). 각 배양조에서 출현한 남조류 우점종은 GS배양조에서
이상과 같이 수온 20℃, 40 µmol m−2s−1 광도조건에서 3~4일 후 저서성 남조류에 의한 매트가 형성되어, 10~12일 후에는 매트가 부상되는 것을 확인하였다. 저서성 남조류 매트의 생성 및 부상은 조류 현존량의 변화에 영향을 미치며, 이취미 물질인 geosmin과 2-MIB 농도 변화에도 중요한 인자가 되는 것을 확인하였다. 또한 호소에서 부유성 남조류가 출현하지 않더라도 저서성 남조류에 의해 이취미가 유발될 수 있다는(Chen et al., 2010; Sugiura et al., 1998) 것 이 본 연구에서도 확인되었다. 또한 공지천 지점의 경우 2013년의 이취미 발생원인 연구(Han River Watershed Management Committee, 2013)에서도 6~12월에 이취미 원인 조류가 출현하지 않았음에도 geosmin이 32.8±24.5 ng/L의 범위로 검출되었으며, 본 연구에서와 같이 저서성 남조류에 의해 이취미 물질이 발생한 것으로 사료된다.
일반적으로 이취미물질은 조류의 성장기와 안정기에는 체외로 거의 배출되지 않지만 사멸기에는 체내 축적된 물질이 체외로 배출되는 것으로 알려져 있다(Bae et al., 2008; Jüttner and Watson, 2007) 본 연구 결과에서도 저서성 남조류 매트의 부상에 따라 조류가 사멸하여 이취미물질이 증가하는 것으로 판단된다. 그러나 표층에 남조류가 출현하지 않는 원인불명의 이취미 발생이 모두 저서성 남조류에 의한 것으로 판단할 수는 없다. 이취미 발생 원인생물로 남조류 외에 곰팡이(
의암호 내 춘천하수처리장방류구 부근의 퇴적층(US)과 유입지천인 공지천 하류지점 퇴적층(GS)을 배양하여, 이화학적인 환경요인의 변화와 조류군집 형성 및 이취미 발생을 분석하여 다음과 같은 결론을 얻었다.
1) 퇴적층 표층에 저서성 남조류 매트가 증가되어 pH와 DO농도가 증가하게 되고, 최대 pH 9.3까지 증가하여 착물로 형성된 인산염인의 용출로 0.044 mg/L에서 0.472 mg/L로 약 10배까지 증가하는 것을 확인하였다. 또한 퇴적층 매트의 부상으로 인하여 전기전도도 증가가 확인되었다.
2) 식물플랑크톤 군집은 배양기간 동안 GS에서 45분류군, US에서 40분류군이 출현하였으며, 두 배양조 모두 녹조류가 가장 다양하게 출현하였고, 유해남조류로 분류된
3) 퇴적층 표층의 배양초기 휴면포자 현존량은 GS배양조는 500 cells/g, US는 2,200 cells/g로서 배양에 따라 휴면포자는 감소하여 배양 4일째에 약 10%로 감소하였다. 표층에서
4) 퇴적층 표층의 매트에서는
5) 이취미 물질은 남조매트의 부상시기를 전후로 geosmin 농도가 가장 높았으며(GS: 115 ng/L, US: 175 ng/L), GS의 경우 두 번째 남조류 매트 부상이후에는 2-MIB의 농도가 22~92 ng/L의 범위로 증가하여, 저서성 남조류 매트의 형성과 부상이 이취미물질 발에 중요한 인자가 되는 것을 확인하였다.
1) 퇴적층 표층에 저서성 남조류 매트가 증가되어 pH와 DO농도가 증가하게 되고, 최대 pH 9.3까지 증가하여 착물로 형성된 인산염인의 용출로 0.044 mg/L에서 0.472 mg/L로 약 10배까지 증가하는 것을 확인하였다. 또한 퇴적층 매트의 부상으로 인하여 전기전도도 증가가 확인되었다. 2) 식물플랑크톤 군집은 배양기간 동안 GS에서 45분류군, US에서 40분류군이 출현하였으며, 두 배양조 모두 녹조류가 가장 다양하게 출현하였고, 유해남조류로 분류된 Microcystis, Anabaena, Oscillatoria 가 표층에서 관찰되었으며, 배양 경과시간의 증가에 따라 저서성 남조류 출현이 증가하였다. 3) 퇴적층 표층의 배양초기 휴면포자 현존량은 GS배양조는 500 cells/g, US는 2,200 cells/g로서 배양에 따라 휴면포자는 감소하여 배양 4일째에 약 10%로 감소하였다. 표층에서 Anabena 가 출현시기는 US는 배양 4일째, GS는 15일째에 출현하여 퇴적층에 분포한 휴면포자가 발아하여 수체에서 검출되는 것으로 판단된다. 4) 퇴적층 표층의 매트에서는 Oscillatoria tenuis, O. limosa 및 Lyngbya hieronymusii 등의 저서성 남조류가 출현하였다. GS에서 최대 3.96×106 cells/cm2, US에서 최대 5.77×106 cells/cm2 로 매트를 형성하였으며, 배양시작 10~12일 후 매트가 부상하였다. 5) 이취미 물질은 남조매트의 부상시기를 전후로 geosmin 농도가 가장 높았으며(GS: 115 ng/L, US: 175 ng/L), GS의 경우 두 번째 남조류 매트 부상이후에는 2-MIB의 농도가 22~92 ng/L의 범위로 증가하여, 저서성 남조류 매트의 형성과 부상이 이취미물질 발에 중요한 인자가 되는 것을 확인하였다.