Norovirus, a genus in the family Caliciviridae, is a group of non-enveloped viruses that have a single-stranded, positive sense RNA genome (Atmar 2010). Norovirus infection can usually be caused by contaminated water or food, and it spreads via human contact with infected materials through the fecal-oral route (Choi et al. 2014). This infection has been recognized as a leading cause of epidemics with the symptoms of vomiting, diarrhea, mild fever, abdominal cramping, and nausea in the community (La Rosa et al. 2013). According to the Centers for Disease Control and Prevention (2011), over 70% of water-related gastroenteritis patients and over 50% of patients due to food-consumption resulted from norovirus infection in USA. Likewise, norovirus causative issues have been a great concern over the world during the recent years.
Norovirus has the characteristics such as low infectious dose, prolonged shedding period, strong stability, great diversity, and frequent genome mutations (Lee et al. 2014b). Norovirus could be effectively reduced by using disinfectants such as alcohols and povidone idodines (Belliot et al. 2008). However, chemical sanitizers can cause various side effects in humans, such as fever and itching (Choi et al. 2014). Recently, attempts have been made to find sustainable solutions from medicinal plants and marine organisms (Balunas and Kinghorn 2005, Jain et al. 2008, Choi et al. 2014). Among marine organisms, chitosan and chitosan oligosaccharides were reported to be able to effectively reduce the infectivity of human enteric viral surrogates (feline calicivirus [FCV] F-9) (Su et al. 2009, Davis et al. 2012). However, there are limitations in the progress of this study, due to inefficient cell culture amplification process for norovirus (Guix et al. 2007, Lay et al. 2010). Recent studies revealed that the structure and genetic relatedness of murine norovirus (MNV) to human norovirus makes this virus a promising and relevant surrogate for studying the environmental survivability of human norovirus (Cannon et al. 2006, Zhang et al. 2012).
In this study, we investigated the possibility of using the marine alga Eisenia bicyclis extract and its ingredients as an alternative antiviral agent against MNV. The brown algae have also been reported to exhibit several antimicrobial activities against pathogenic bacteria and FCV (Eom et al. 2013a, Choi et al. 2014, Lee et al. 2014a). Since inadequate scientific research findings are available on the antiviral activity from marine organisms against MNV, the present study with MNV as a norovirus surrogate may have great contribution to the development of effective antiviral agents to control human norovirus.
In the late September of 2013, E. bicyclis was purchased from Ulleung Trading Co. (Ulleung, Korea). A voucher specimen was refrigerated at -80°C. Dried E. bicyclis was triturated into powder with electronic grinder (HMF-1000A; Hanil Electronics, Seoul, Korea). The dried E. bicyclis powder (1.0 kg) was extracted with methanol (MeOH; 10 L × 3) at 68°C for 3 h (3 times), and the solvent was evaporated using rotary evaporator (Eyela Co., Tokyo, Japan) under vacuum at 45°C. The combined crude MeOH extract (164.3 g) was suspended in 10% MeOH (1.0 L), and then it was fractionated in turn with n-hexane (Hexane; 1.0 L × 3), dichloromethane (DCM; 1.0 L × 3), ethyl acetate (EtOAc; 1.0 L × 3), and n-butanol (BuOH; 1.0 L × 3) solution, to yield Hexane-soluble extract (42.3 g), DCM-soluble extract (2.5 g), EtOAc-soluble extract (23.8 g), BuOH-soluble extract (26.5 g), and H2O-soluble extract (69.1 g).
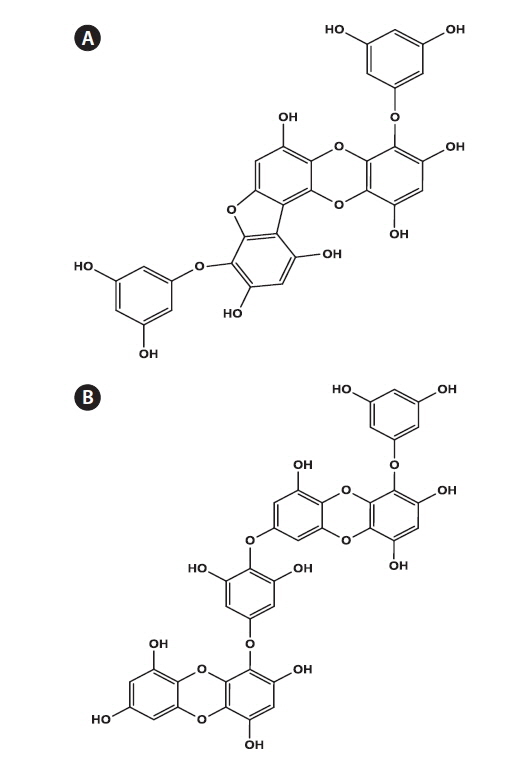
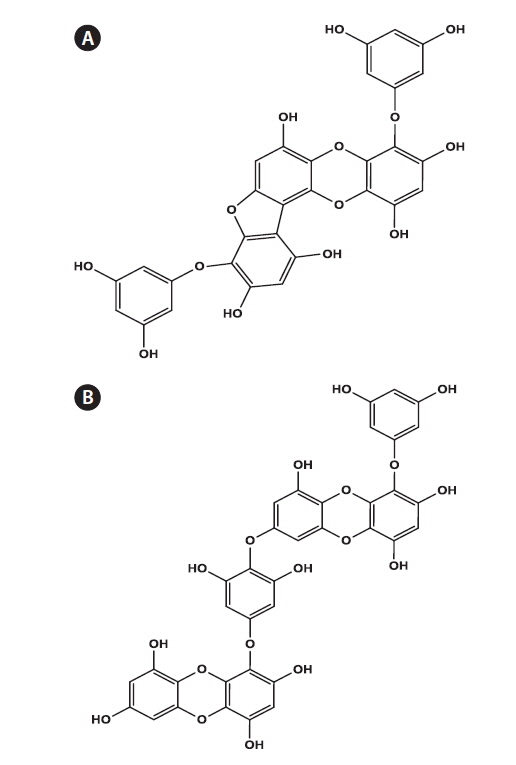
Phlorofucofuroeckol-A (PFF) Fig. 1A and dieckol (DE) Fig. 1B were isolated from the EtOAc-soluble extract of E. bicyclis by using Sephadex LH-20 and RP-18 open column chromatography, as a part of previous research, along with several other phloroglucinol derivatives (Eom et al. 2013b) (Fig. 1).
RAW 264.7 cells, obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA), were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Gibco BRL, Grand Island, NY, USA) with 1% penicillin (100 U mL-1; Gibco BRL)-streptomycin (100 μg mL-1; Gibco BRL) and heat inactivated 10% fetal bovine serum (Gibco BRL) at 37°C in 5% CO2 atmosphere. MNV-1 was obtained from Dr. Gwang Pyo Ko at Seoul National University (Seoul, Korea) with the permission of Dr. Herbert W. Virgin at the Washington University School of Medicine (St. Louis, MO, USA); and it was propagated in RAW 264.7 cells. RAW 264.7 cells were harvested three times by using freezethawing method. The virus stock was stored at -80°C for further use.
The cytotoxicity of E. bicycilis extracts was determined by quantifying RAW 264.7 cells viability using MTT (3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide) assay. The medium containing RAW 264.7 cells were cultured into a 96-well plate at a density of 3 × 105 cells mL-1. The 50% cytotoxicity concentration (CC50) value was examined following 24 h of exposure of RAW 264.7 cell monolayers to different concentrations of extracts. For control, equal volume of medium was added and incubated at 37°C in 5% CO2 atmosphere. Formazan solubilizing solution was added after incubation. Absorbance of each well was measured at 540 nm using a microplate reader (Molecular Devices, Sunnyvale, CA, USA). The optical density of formazan formed by untreated cells was taken as 100% of viability.
The antiviral activity of E. bicyclis extracts against MNV was evaluated by tissue culture infectivity dose 50% (TCID50), according to the method used by Kim et al. (2010). The antiviral effective concentration was expressed as 50% effective concentration (EC50) value, which is defined as the concentration of the sample required to exhibit only 50% of cytopathogenic effects caused by the virus (Goldman and Prabhakar 1996, Goodfellow et al. 2005, Choi et al. 2014). Concentration of 5 log TCID50 mL-1 MNV suspensions was treated with identical volumes of serially diluted extracts at room temperature for 24 h. The mixtures were added to a monolayer of RAW 264.7 cells in a 96 well plate. After incubating at 37°C for 24 h, the cells were fixed with formaldehyde and stained with crystal violet solution to be monitored for cytopathic effects. Untreated control cells were suspended with maintenance medium (DMEM; Gibco BRL), instead of extracts or phlorotannins. The relative effectiveness for inhibiting viral infection inducing cell death is defined as the selectivity index (SI; CC50 EC50-1).
Data were represented as the means for three replicate samples and expressed as the mean ± standard deviation. A student’s t-test and one-way ANOVA were used by SPSS ver. 12.0 (SPSS Inc., Chicago, IL, USA). p < 0.05 was considered statistically significant. The CC50 and EC50 were calculated by regression analysis for the dose-response curves.
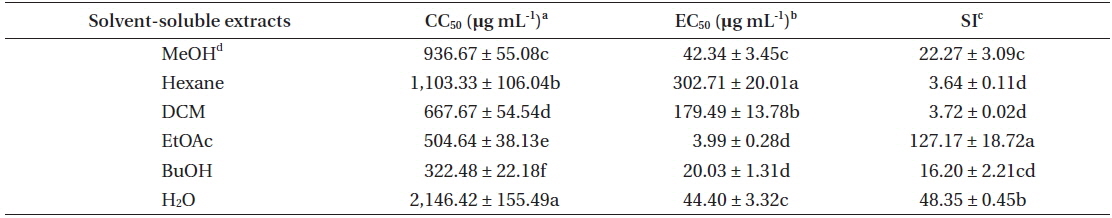
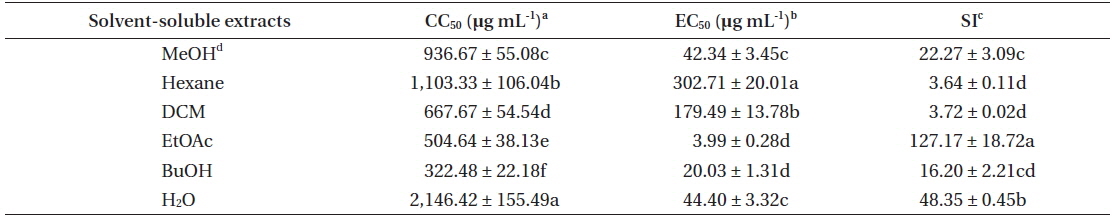
In recent years, norovirus has gained a lot of attention concerning the management of food safety, since 50% of all outbreaks related to food-poising were caused by norovirus across the nations (Centers for Disease Control and Prevention 2011). Although various biological activities of marine organism have been reported, there are still many unknown potentials that could be utilized. We previously reported that E. bicyclis methanolic extract exhibit antiviral activity against FCV (Choi et al. 2014). However, as described above, FCV may not be the most suitable surrogate for evaluating the efficacy of disinfectants against human norovirus (Cannon et al. 2006, Zhang et al. 2012). Therefore, the antiviral activity of E. bicyclis extracts against MNV as a norovirus surrogate was evaluated. Cytotoxicity of E. bicyclis extracts was evaluated by determining CC50 value using RAW 264.7 cells. It has been proposed the following cutoff points to determine a cytotoxicity of plant extracts toward normal cells: 1) significant or strong cytotoxicity: <100 μg mL-1; 2) moderate cytotoxicity: 100 μg mL-1 to <300 μg mL-1; 3) low cytotoxicity: 300 μg mL-1 to <1,000 μg mL-1; 4) no cytotoxicity: <1,000 μg mL-1 (Kuete and Efferth 2015). Thus, the MeOH extract and its solvent-soluble extracts showed no severe cytotoxicity towards RAW 264.7 cells with the CC50 values of 322.48 to 2,146.42 μg mL-1 (Table 1). The cytotoxicity of E. bicyclis methanolic extract against RAW 264.7 cells was higher than that of our previous report using CrFK cells (CC50 = 410) (Choi et al. 2014). This result could be due to the difference in toxic susceptibility of host cells to the methanolic extract. Antiviral activity of E. bicyclis extracts, inhibiting viral infection of host cells, was evaluated by measuring EC50 value as described in MATERIALS AND METHODS. E. bicyclis methanolic extract showed an antiviral activity against MNV in RAW 264.7 cells with EC50 value of 42.34 μg mL-1 (Table 1). It has been reported that E. bicyclis methanolic extract exhibit EC50 value of 80 μg mL-1 against FCV (Choi et al. 2014). Then, the antiviral activity of the methanolic extract was evaluated by SI analysis, as described above. It is desirable to have a high SI giving maximum antiviral activity, with minimal cell toxicity as well as higher possibility of application (Oh et al. 2013). The SI value of E. bicyclis methanolic extract against MNV is 22.27, while SI value of the same extract against FCV is 5.13 (Choi et al. 2014). Thus, E. bicyclis methanolic extract exhibits higher inhibiting effectiveness against MNV infection in host cells, compared to that of FCV. In addition, the SI value is similar to that of green tea (Camelia sinensis) methanolic extract with SI value of 18.57 against FCV (Oh et al. 2013).
To elucidate an active substance(s) responsible for the antiviral activity against MNV, the methanolic extract was further partitioned as Hexane, DCM, EtOAc, BuOH, and H2O soluble extracts. The antiviral activity of solvent-soluble extracts was also evaluated by the SI analysis. Accordingly, EtOAc extract showed the highest SI value, followed by H2O, BuOH, DCM, and Hexane soluble extract (Table 1). These results clearly showed that EtOAc extract has strong antiviral activity with low cytotoxicity. Thus, the extract will be a good choice for the search of novel antiviral compounds against MNV.
Several phlorotannins, phloroglucinol derivatives, from the EtOAc-soluble extract of E. bicyclis were previously isolated. Of these, DE and PFF exhibited the strongest antibacterial activity against pathogenic microbes such as Staphylococcus aureus and Propionibacterium acnes (Eom et al. 2013a, Lee et al. 2014a). The structural characteristics of DE and PFF have been reported earlier (Eom et al. 2013b) (Fig. 1). Therefore, in the present study, the antiviral activity of these compounds against MNV in RAW 264.7 cells was demonstrated, as described above. The CC50 values of DE and PFF were calculated to be 495.71 μM and 579.02 μM, respectively (Table 2). DE and PFF inhibited the infection of MNV in RAW 264.7 cells with an EC50 value of 0.90 μM. Consequently, the calculated SI values of DE and PFF were 550.60 and 668.87, respectively, which is suggesting that PFF can more effectively inhibit MNV infection in host cells compared to that of DE (Table 2). Compared to epigallocatechin gallate (a catechin from green tea) with SI value of 26.67 against FCV (Oh et al. 2013), the present study showed SI values of DE and PFF against MNV were about 20 and 25 times higher, respectively, than that of epigallocatechin gallate. This finding clearly highlights the existence of potential antiviral compounds against MNV in edible brown alga E. bicyclis.
Recent studies have revealed the antiviral effect of plant-derived compounds against MNV (Li et al. 2013). However, to our knowledge, there has been no previous report on marine-derived polyphenols exhibiting antiviral activity against MNV. Choi et al. (2014) reported that E. bicyclis methanolic extract possess an anti-norovirus activity which inhibits FCV infection in CrFK cells with EC50 value at the concentration of 80 μg, but MNV belongs to the human norovirus unlike FCV. In addition, MNV is demonstrated as enteric pathogens of mice and humans, respectively, whereas FCV infects via nasal, oral, and conjunctival routes (Radford et al. 2007, Howell and D’Souza 2013). MNV has been increasingly used as a surrogate for human norovirus in virucidal efficacy evaluations (Howell and D’Souza 2013). Our results provide evidence that the phlorotannins (DE and PFF) could provide a way to develop effective marine-derived antiviral agents against human norovirus. However, further antiviral experiments should be performed to delineate how phlorotannins deactivate MNV and the mechanism of action. Generally, although it is not fully understood, the antiviral action mechanism of natural compounds against human norovirus or its surrogates is usually supposed to be the prevention of the viral attachment to host cells (Howell and D’Souza 2013).
The EtOAc-soluble extract from E. bicyclis exhibited the strongest antiviral activity against MNV among five solvent-soluble extracts. The antiviral activity of EtOAc-soluble extract against MNV may also be correlated with their marine-derived polyphenolic components. Therefore, previously isolated phlorotannins, such as DE and PFF, were evaluated for antiviral activity against MNV. DE and PFF demonstrated strong anti-MNV activity with low cytotoxicity against RAW 264.7 cells. The results of the present investigation are expected to contribute to the development of an alternative phytotherapeutic agent against MNV, as natural disinfectant for the improvement in food safety.