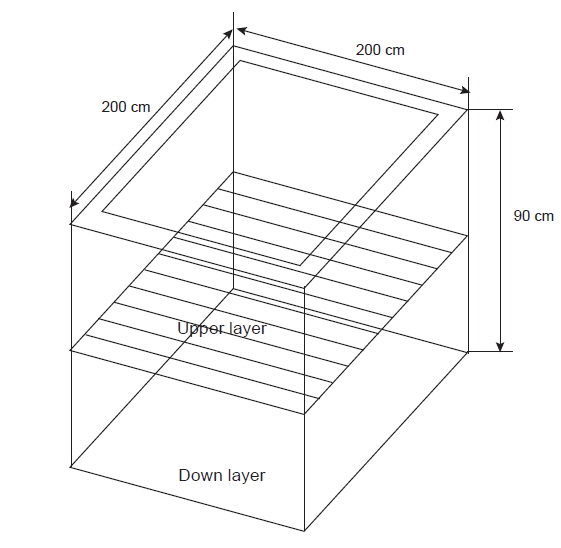
우리나라 양식산 넙치(
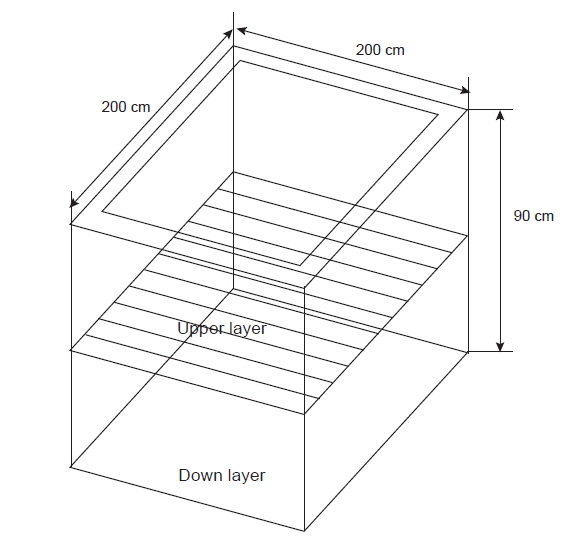
해상장거리 대량수송장치는 활어의 양식산 넙치를 저온 및 절식에 의하여 대사활동을 최소화하여 수송하는 원리이다. 이러한 시스템은 생물을 수송하기 때문에 반드시 수조내에 해수를 넣어 생물을 운송하게 된다. 따라서 수온은 활어 수송에 매우 중요한 인자이다. 낮은 수온은 해수 중 pH 유지와 활어의 대사활동을 감소시킨다. 일반적으로 여름철 냉수성 어종의 최적 수송 수온은 6-8℃이며, 온수성 어종은 10-12℃이다. 봄과 가을철은 각각 3-5℃과 5-6℃, 겨울철은 1-2℃로 알려져 있다(Berka, 1986). 컨테이너 내부에 수조, 냉각기, 산소발생기, 여과장치, 전기장치 등을 갖추고 있어 수송 중 온도와 산소를 일정하게 조절하고, 생물의 대사활동으로 발생하는 대사산물을 여과조를 거쳐 안정적으로 수송할 수 있는 조건을 확립하였으며, 수송온도와 절식이 넙치 근육의 물리, 화학적 변화를 연구하였다(Shim et al., 2012).
따라서 본 연구에서는 해상장거리 대량수송장치에서 수송밀도에 따른 양식산 활넙치의 물리화학적 변화를 조사하여 안정적인 수송을 위한 최적 조건을 확립하고자 하였다.
넙치(
Ando et al. (1991)의 방법에 따라 어육을 일정크기로 절단하여(20×20×10 mm3), 직경 10 mm cylinder plunger를 사용한 rheo meter (Compac-100, Sun, Japan)로 측정하였다.
Iwamoto et al. (1987)의 방법으로 ATP 관련 물질을 추출하여 HPLC (Surveyor Plus HPLC System, USA )를 이용하여 분석하였다.
Barker and Summerson (1941)의 방법에 따라, 근육 2.5 g을 10% TCA용액 22.5 mL을 가하여 균질화하고, 원심분리(398
글리코겐 함량은 시료 0.5 g에 30% KOH 5 mL를 첨가하여 95℃에서 20분간 중탕하고 포화 Na2SO4 용액 0.5 mL와 ethanol 5 mL를 첨가하여 95℃에서 다시 15분간 중탕한 후 원심분리(1,259
조지방은 AOAC법에 따라 Soxhlet 추출법으로 측정하였다(AOAC, 1995).
통계 처리는 SAS 프로그램을 이용한 분산분석표를 작성하였으며, Duncan의 다중범위검정(Duncan's multiple range test)으로
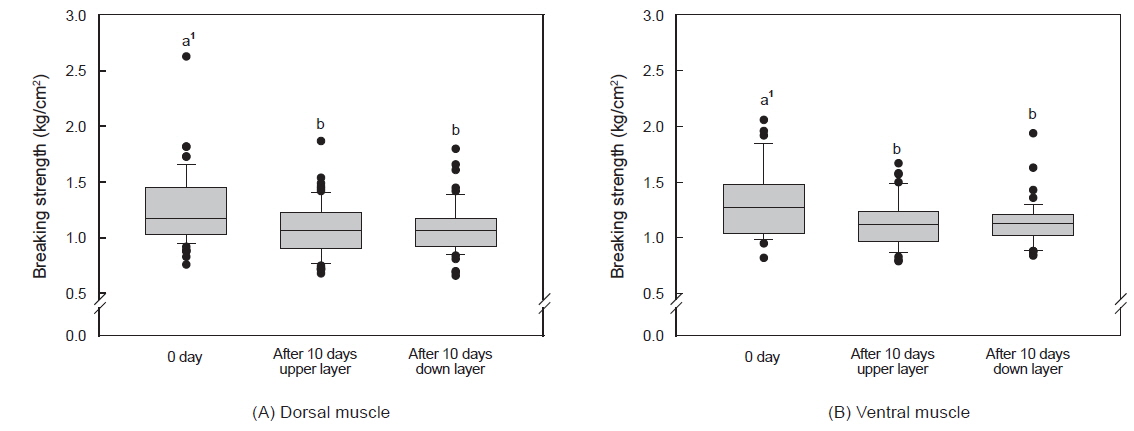
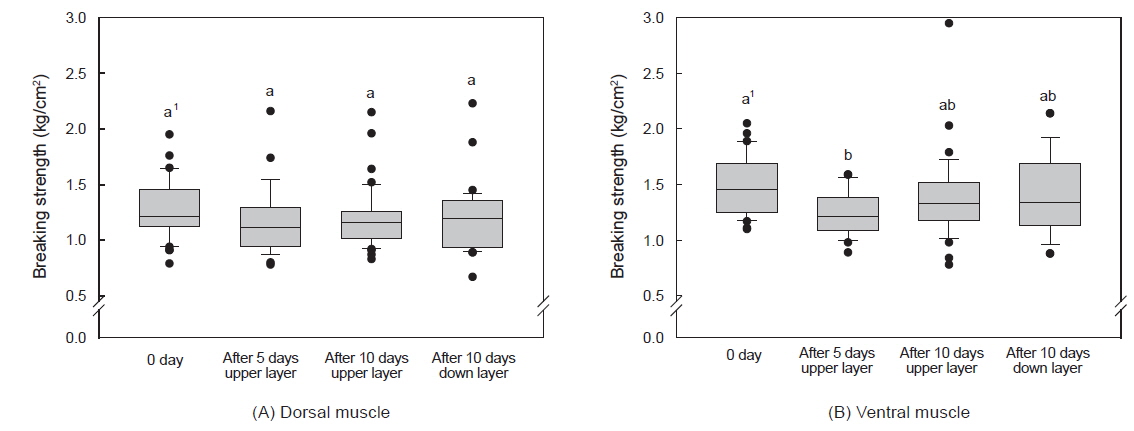
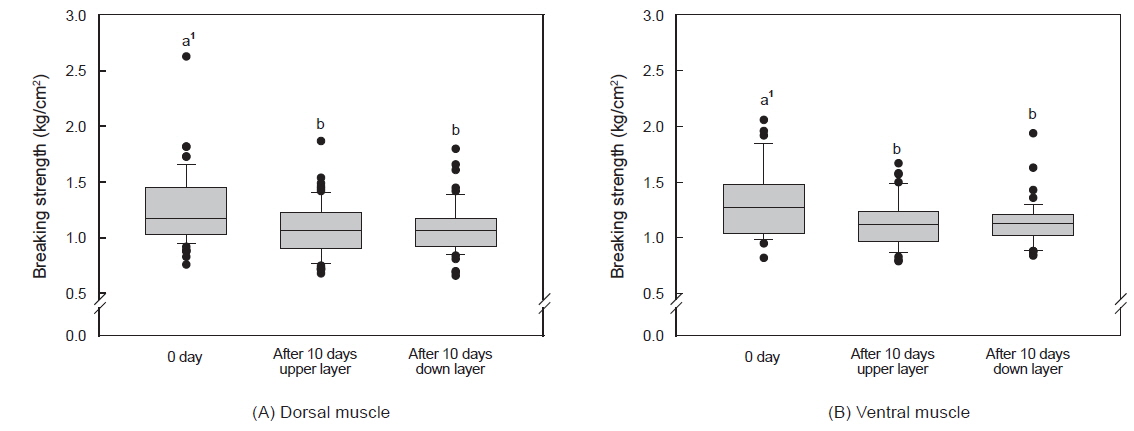
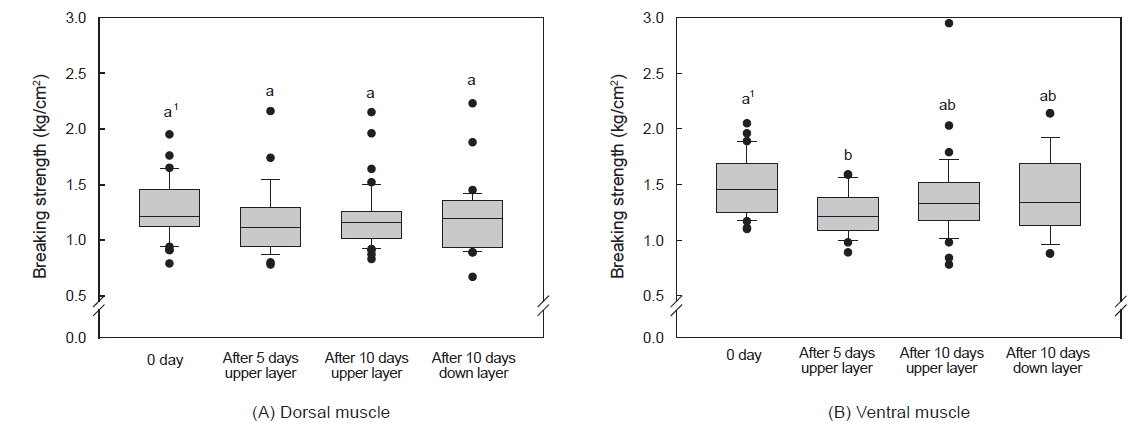
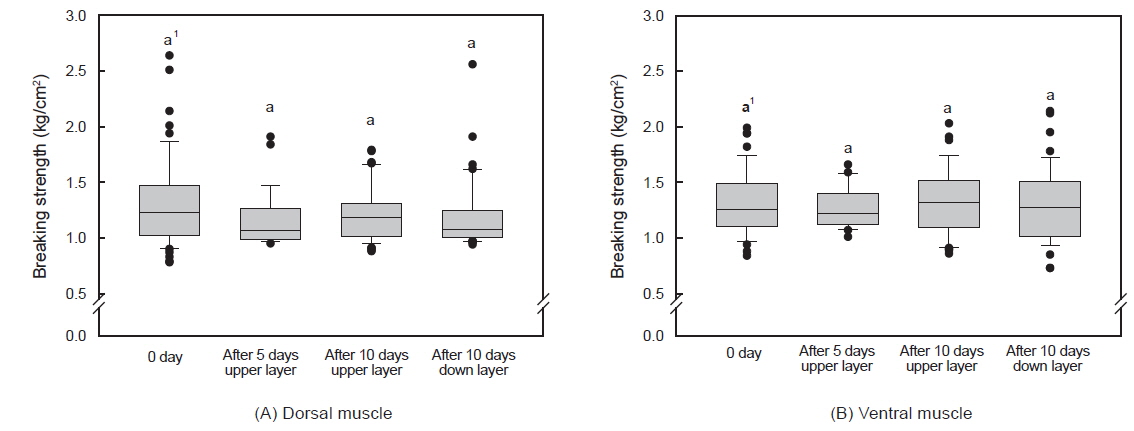
수송밀도에 따른 등 근육이 배 근육에 비하여 근육강도 값이 다소 높았으며, 500 kg을 제외하고는 수송 기간에 따른 유의적인 차이는 없었다(
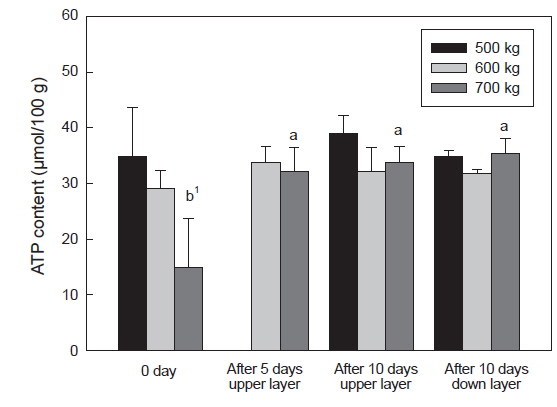
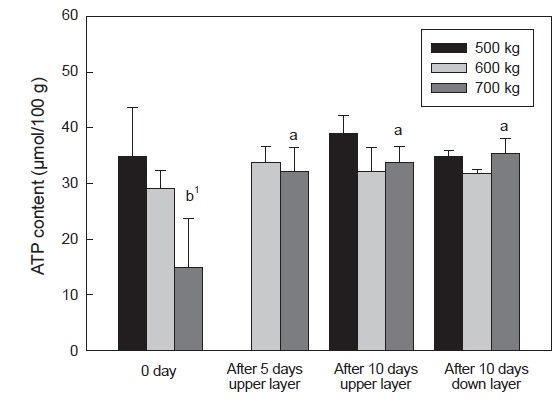
수송 밀도에 따른 넙치 등 근육의 ATP 및 lactate 함량은 Fig. 5와 6에 나타내었다. 수송 밀도 500 kg에서는 수송 직후에 34.8 μmol/g이었으나 10일 경과 후에는 상단과 하단에 각각 39.1 및 34.9 μmol/g로 수송기간에 따른 유의적인 차이는 없었다. 수송 밀도 600 kg에서는 수송 직후에 500 kg 수송밀도 보다는 다소 낮은 함량인 29.04 μmol/g 이었으며, 수송 5일과 10에는 다소 증가하여 각각 33.8, 32.1, 31.7 μmol/g이었다. 반면 수송 밀도 700 kg에서는 높은 수송밀도로 인하여 ATP 함량이 기타 수송 밀도에 비하여 감소하여 14.9 μmol/g이었으며, 이후 수송기간 동안 증가하여 32.1-35.3 μmol/g이었다(Fig. 5).
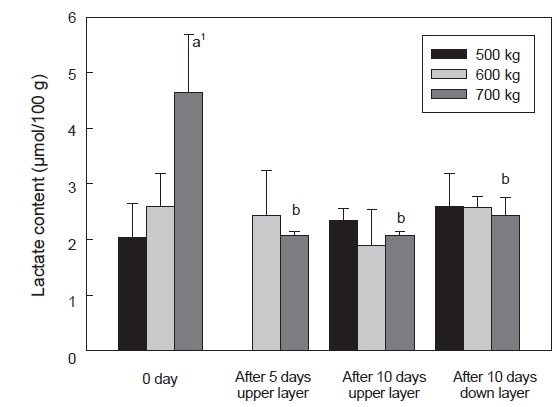
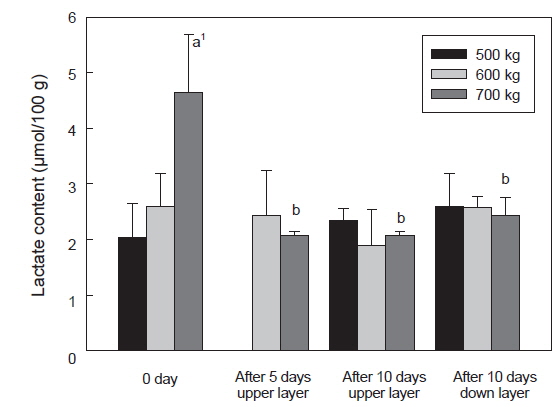
Lactate 함량은 수송 밀도에 따른 ATP 함량 변화와 유사하였으며, 수송 밀도 500, 600 kg에서는 수송 전과 10일 경과 후에 각각 2.0-2.6 및 1.9-2.6 μmol/g이었다. 반면 수송 밀도 700 kg에서는 수송 전에 급격히 증가하여 4.7 μmol/g이었으며, 수송 기간 동안 기타 수송밀도와 유사한 lactate 함량을 나타내었다 (Fig. 6).
수송직후에 근육 내의 ATP 소모와 lactate 축적의 변화가 큰 것은 수송온도 및 수송밀도 에 의한 스트레스로 인한 것으로 사료되며, 특히 수송밀도가 700 kg에서 이들 성분의 변화 폭이 큰 것은 수송밀도의 증가로 인한 어류가 받는 스트레스가 증가하는 것으로 사료된다. 일반적으로 스트레스를 받은 어류의 근육은 죽기 전에 많은 양의 에너지를 소비하게 되며, 이로 인하여 해당과정에서 ATP 소모와 lactate 축적이 이루어지게 되며, 스트레스를 받지 않은 어류는 스트레스를 받은 어류에 비하여 초기 ATP소모는 크지 않았다(Nakayama et al., 1992). 따라서 수송온도에 적응한 넙치는 대사활동은 저하되었지만 고밀도의 수송 조건에서 활어에 스트레스가 주어지게 되어 나타나는 현상으로 사료된다. 또한 넙치 혈액 중 glucose 함량이 700 kg 수송 밀도에서는 수송 전보다 수송 후에 증가하였다(결과 미제시). 수송밀도의 증대는 대사활동에 의한 대사산물이 수질악화를 초래하게 되고, 이는 스트레스로 작용하여 체내에 에너지원인 glycogen을 소비하게 되고, 이로 인하여 혈액 내에 glucose 함량이 증대된 것으로 사료된다(Lupatsch et al., 2010; Yosif, 2002).
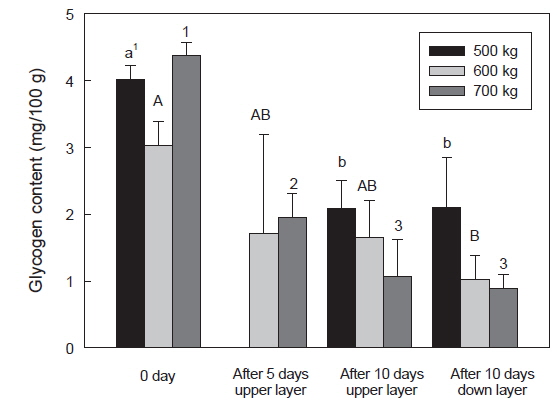
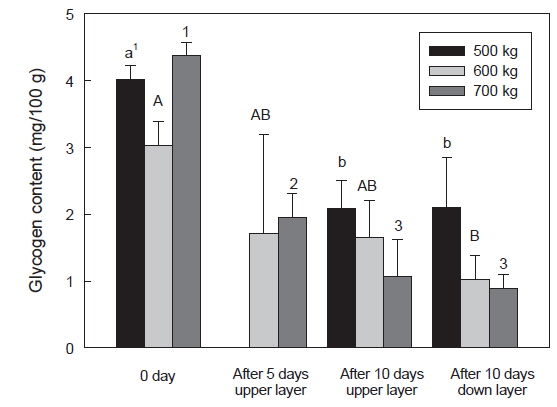
수송밀도에 따른 넙치 근육의 glycogen 함량은 수송 직전보다는 수송 후에 glycogen 함량이 감소하였다. 특히 700 kg 수송 밀도에서는 4.37 g/100 g이었으나 수송 5일에는 1.95 g/100 g 으로 감소하였으며, 수송 10일에는 상단과 하단에 각각 1.07 및 0.88 g/100 g으로, glycogen 함량이 감소하였다(Fig. 7). 어류 근육내의 glycogen 함량은 단기 절식상에서 개체가 크고 수온이 높을수록 큰 폭으로 감소하며, 온도가 낮을수록 감소폭은 양호한 것으로 보고되었다(Sherstneva and Shabalina, 1971). 본 연구에서는 8℃ 수송온도에서 에너지원인 glycogen 함량은 크지 않았지만, 수송밀도의 증가는 근육 내의 glycogen이 감소폭이 큰 것으로 확인되었다.
어류는 대사활동 후 근육내의 glycogen은 lactate와 전환되며, 근육내의 pH가 감소하게 된다. 포유류는 급격하게 형성된 lactate를 간에서 pyruvate로 전환시키지만, 어류는 이러한 과정 없이 체내에 lactate를 축적하게 된다(Hall, 1997). 따라서 체내에 축적된 lactate 함량과 glycogen 함량의 감소는 수송밀도에 의한 어류의 스트레스 정도를 간접적으로 확인할 수 있을 것으로 사료된다.
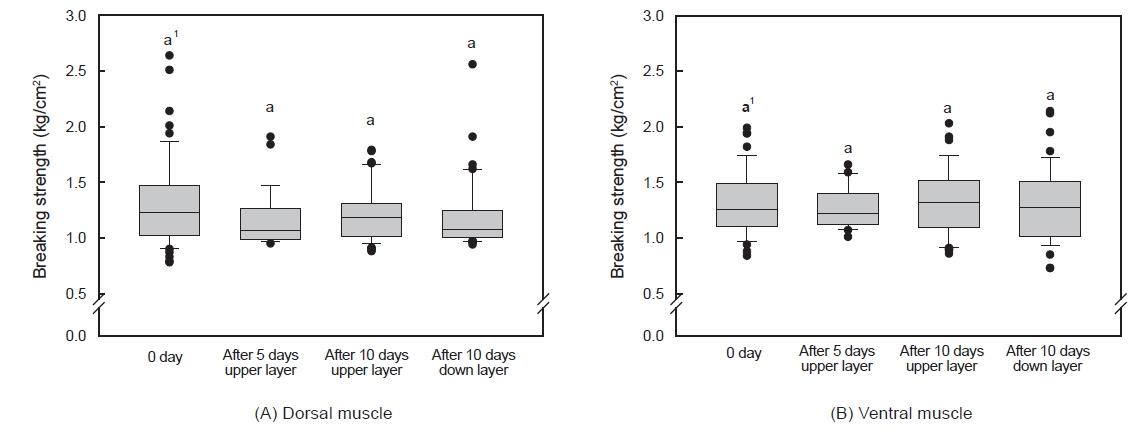
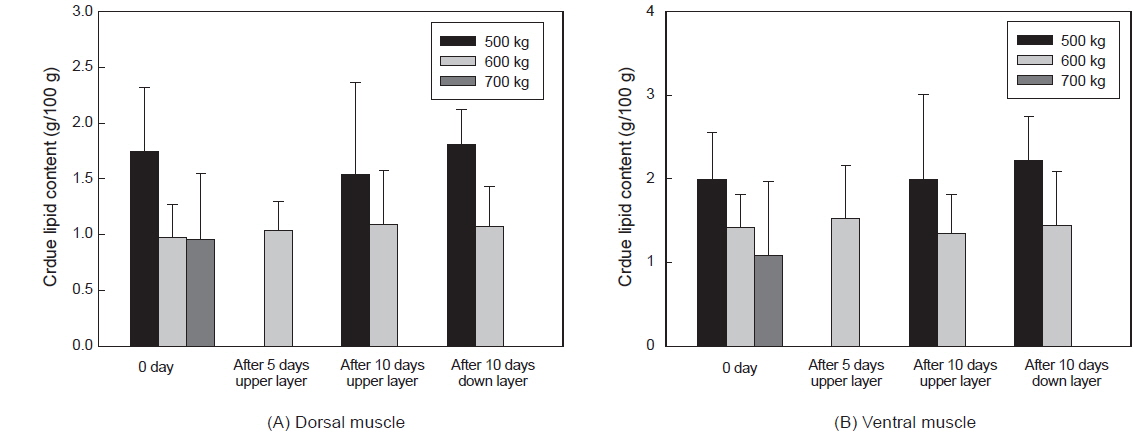
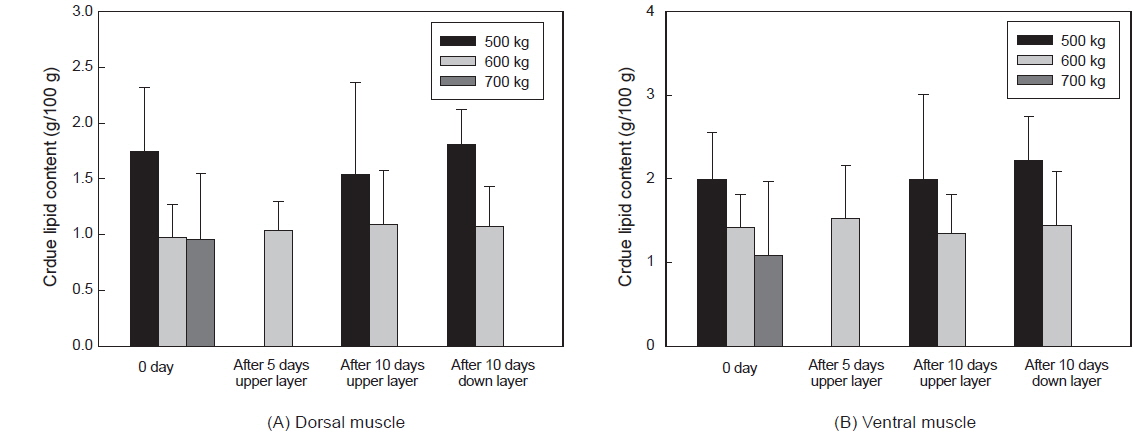
그러나 수송밀도와 수송기간에 따른 조지방 함량은 유의적인 차이가 없었다(
수송밀도에 따른 넙치 근육의 물리·화학적 변화를 조사한 결과, 근육의 물리·화학적 변화는 미미하였다. 그러나 수송기간 10일 동안은 최소 대사에 필요한 에너지원으로 지방질보다는 glycogen을 이용되는 것으로 확인되었으며, 수송밀도의 증가는 glycogen 함량의 큰 변화가 나타났다. 따라서 한 개의 수조내 활어상태 넙치의 안정적인 수송이 가능한 최적 수송밀도는 500 kg, 600 kg이다. 그러나 수송밀도가 증대될수록 스트레스가 증가하고 이로 인한 대사활동과 대사산물은 수조내의 수질상태 악화로 활어의 폐사를 초래할 수 있을 것으로 사료된다.