뱀장어목(Anguilliformes) 바다뱀과(Ophichthidae) 어류에 속하는 바다뱀속(
본 연구에서는 우리나라 제주도 동남부해역 및 한일중간수역에서 부화직후 엽상자어를 포함한 바다뱀 엽상자어17개체가 출현하여 이들의 형태 발달을 상세히 보고하고, 까치물뱀속과의 개체발생학적 특징비교, 해류에 따른 분포와 산란장에 대해 고찰하였다.
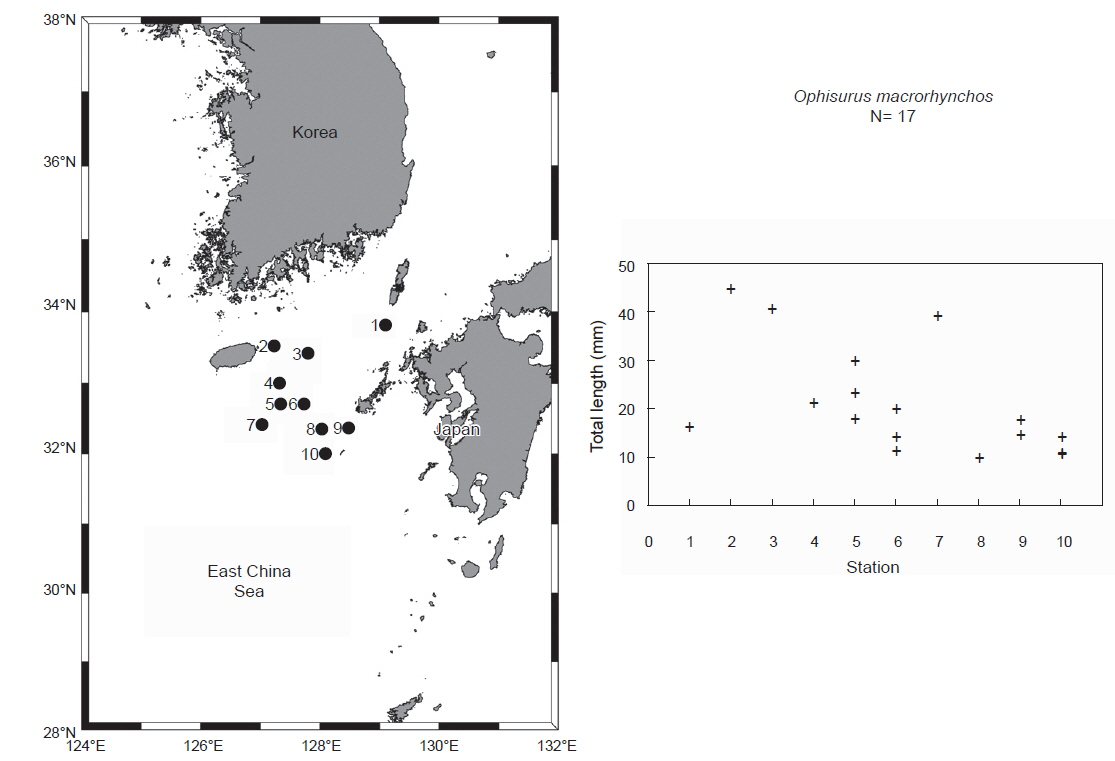
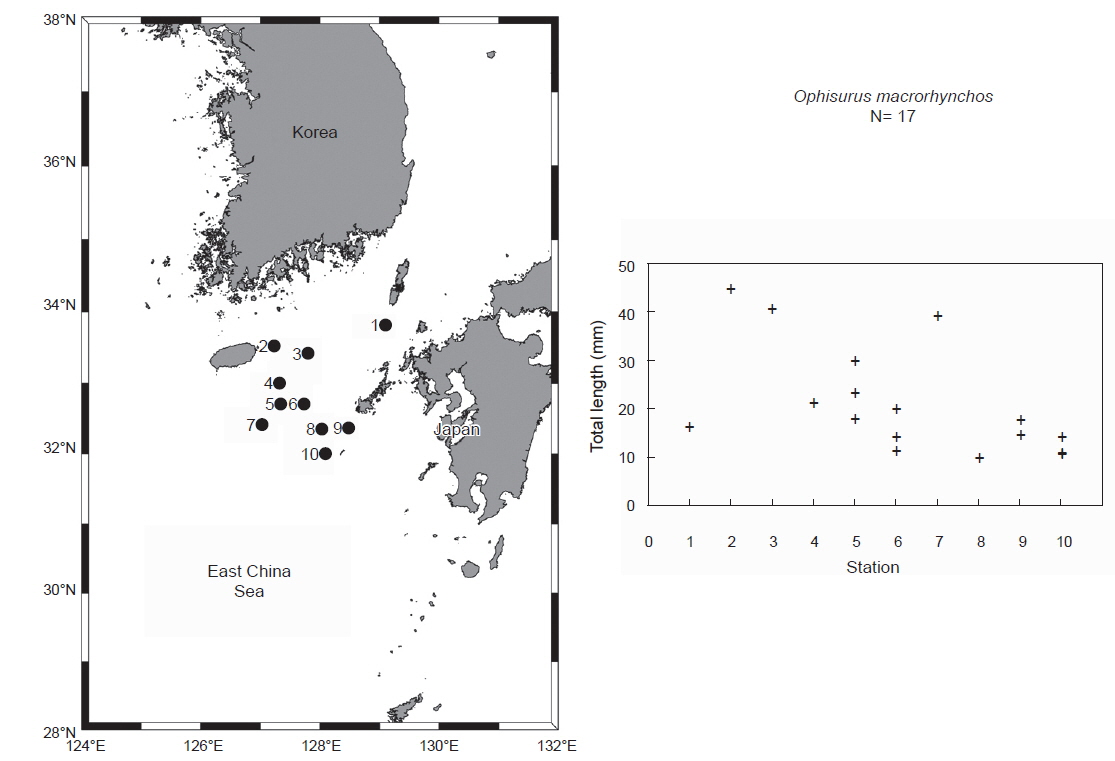
2013년과 2014년 8월 국립수산과학원 시험조사선(탐구 20호)으로 한일중간수역 난자치어 조사 및 제주 동남부 다랑어 난자치어 조사 시 바다뱀과 엽상자어17개체(전장 9.8-44.5 mm)가 Bongo net, IKMT net, 뱀장어네트에서 채집되었다(Fig. 1). 채집 후 표본은 99% 에탄올에 고정하였으며, 이후 국립수산과학원(National Fisheries Research and Development Institute) 난자치어 분석실(Ichthyoplankton Laboratory)에 보관하였다.
몸의 각 부위의 계수 및 계측은 Fahay and Obenchain (1978) 및 Tabeta and Mochioka (1988)의 방법을 따라 입체현미경(Olympus SZX-16) 하에서 관찰하였고, vernier calipers로 0.1 mm 단위까지 측정하였다. 이빨의 배열식은 Castle (1984)를 따랐다.
Genomic DNA는 Chelex resion 100 용액(Bio-rad)을 이용하여 엽상자어의 우측 눈알을 떼어내 추출하였다. 미토콘드리아 DNA 12S rRNA 영역을 종특이적인 12S-F (5'-CAAAGGCCTGGTCCTGACTTTAA-3')와12S-R (5'-CCTTCCGGTACACTTACCATGTTA-3') Primer로 증폭시켰다(Ji and Kim, 2010).
10X PCR buffer 5 μL, 2.5 mM dNTP 4 μL, 12SR primer 5 μL, 12SL primer 5 μL, FX Taq DNA polymerase (GnP, Korea) 0.5 μL를 섞은 혼합물에 genomic DNA 5 μL (100ng)를 첨가한 후, 총 50 μL가 될 때까지 3차 증류수를 넣고 Thermal cycler (Bio-rad MJ mini PTC-1148, USA)를 이용하여 다음과 같은 조건으로 PCR을 수행하였다. Initial denaturation 94℃에서 5분; PCR reaction 35 cycles (denatuation 94℃에서 30초, annealing 56.5℃에서 30초, extension 72℃에서 1분); final extension 72℃에서 7분. 정제는 ExoSAP-IT (United States Biochemical Corporation, USA)을 이용하였다. 염기서열은 ABI 3730XL sequencer (Applied Biosystems Inc., USA)에서 ABI Bigdyeterminator cycle sequencing ready reaction kit v 3.1 (Applied Biosystems Inc., USA)를 이용하여 다음과 같은 조건으로 cycle sequencing하여 얻었다: PCR reaction 35 cycles (denatuation 96℃에서 10초, annealing 56℃에서 10초, extension 60℃에서 3분).
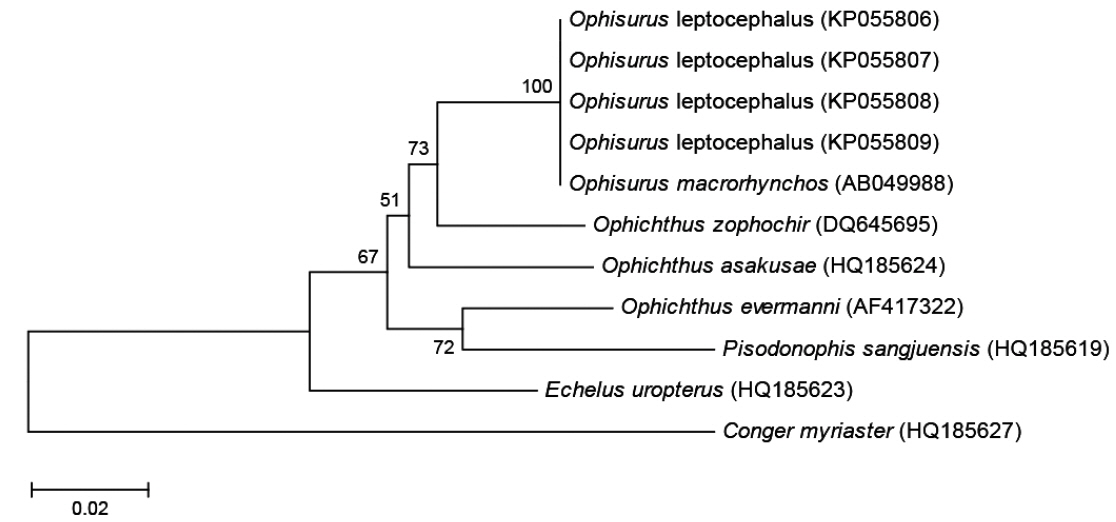
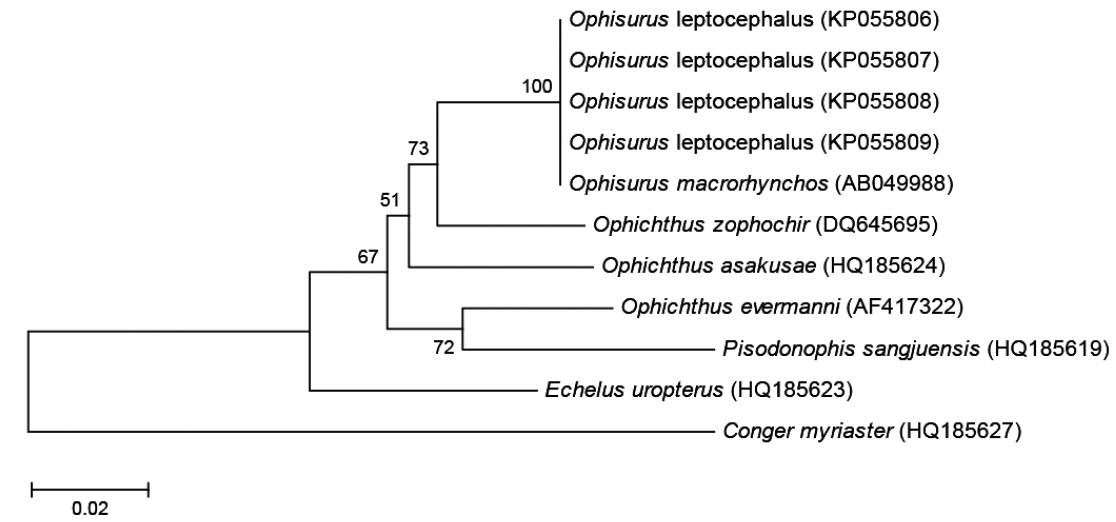
미토콘드리아 DNA 12S rRNA 염기서열은 BioEdit version 7의 ClustalW (Thompson et al., 1994)를 이용하여 정렬하였다. 비교 그룹으로 보유하고 있는 바다뱀과 어류 4종(제주바다뱀,
바다뱀(
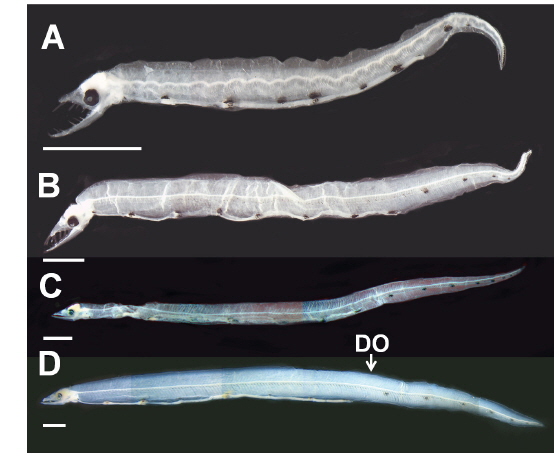
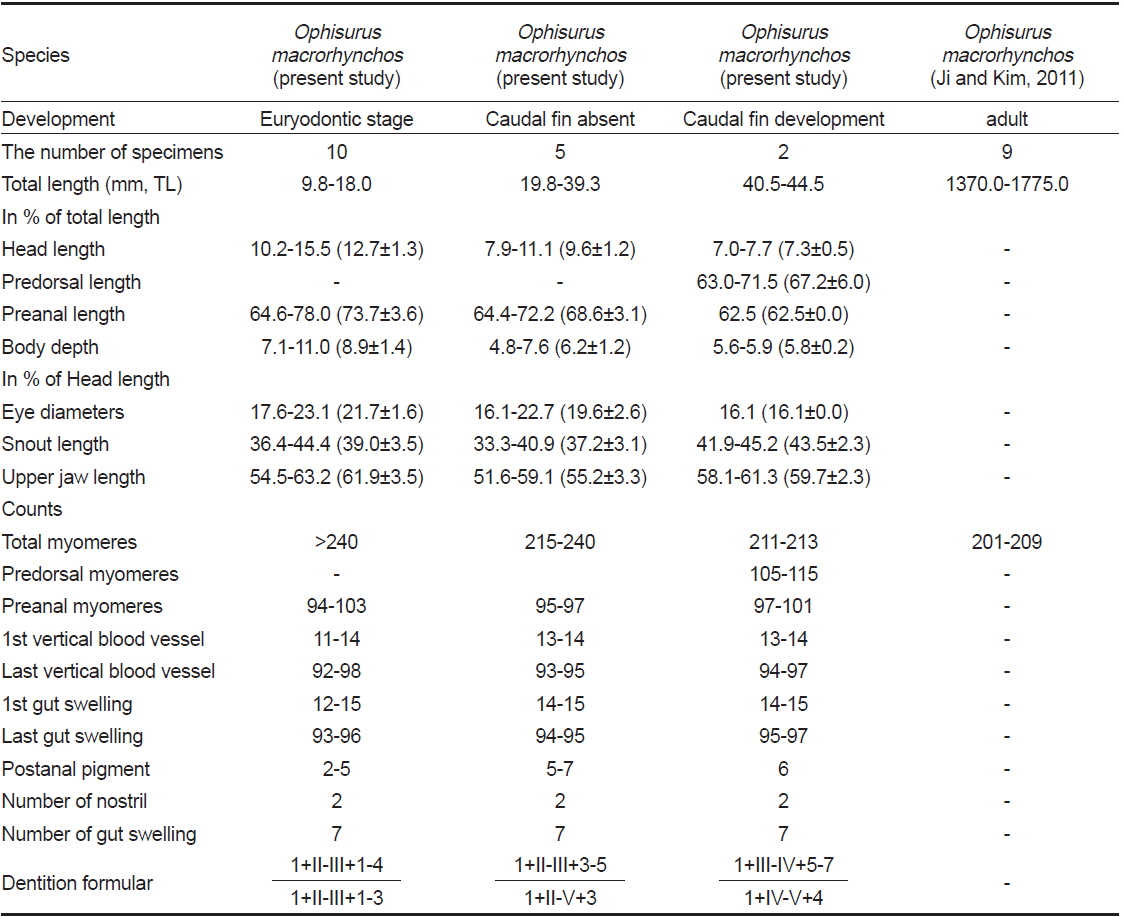
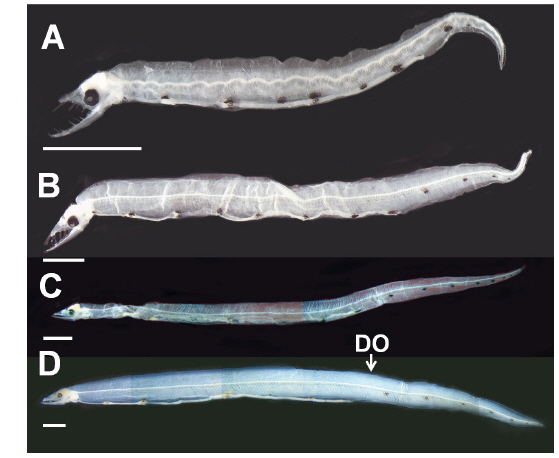
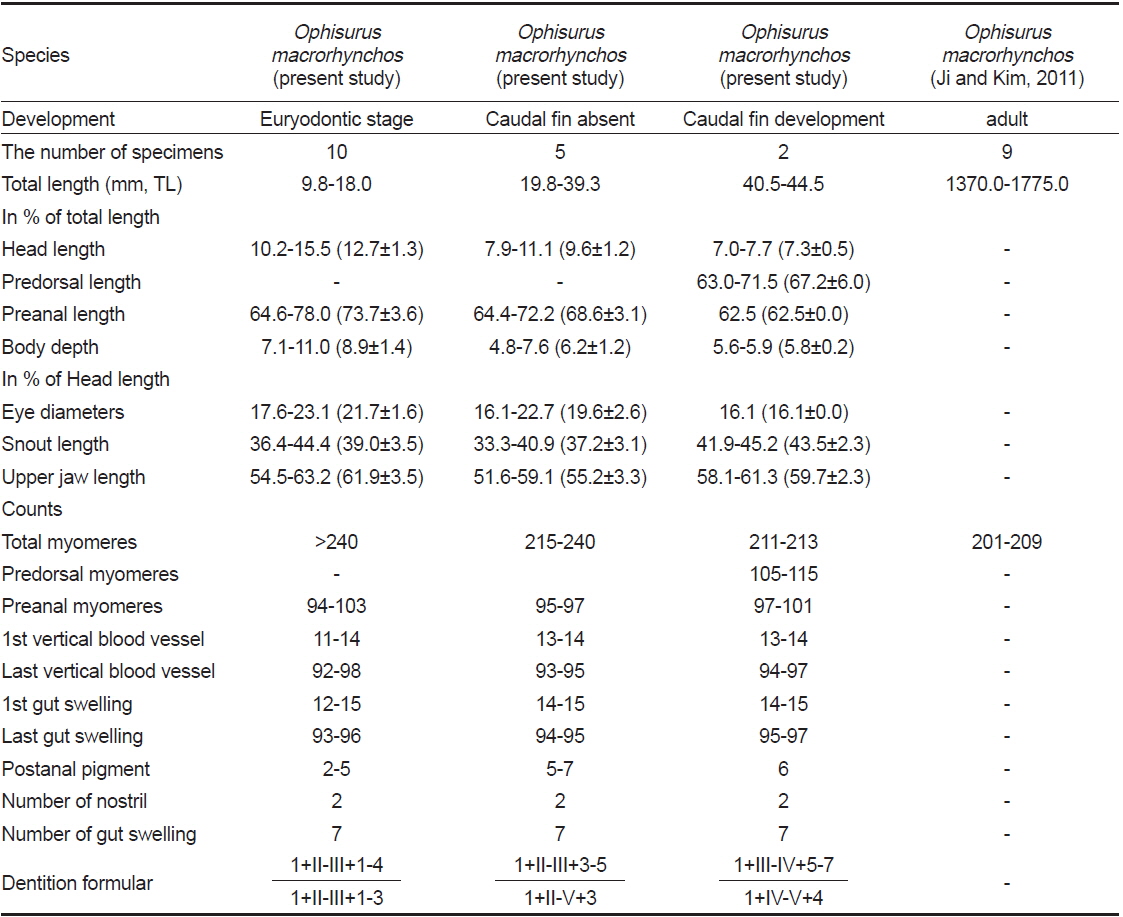
부화직후 추정 엽상자어(전장 9.8-18.0 mm): 전체 근절수는 240개 이상, 항문앞 근절수는 94-103개이다. 머리는 비교적 크고, 주둥이 선단부는 매우 뾰족하다(Fig. 2A, B). 양턱에는 날카로운 이빨이 나 있고, 배열식은 이다. 눈은 작고, 약간 타원형이며 앞쪽에 2개의 비공이 나 있다. 아래턱이 위턱보다 훨씬 전방으로 돌출되어 있고, 위턱은 다소 짧다(Fig. 2A, B). 항문전장은 전장의 64.6-78.0%로 항문은 몸의 중앙 보다 다소 뒤쪽에 위치하며, 꼬리는 몸에 비해 상당히 짧다. 체고는 전장의 7.1-11.0%로 다소 높다(Table 1). 각 지느러미는 막지느러미 형태이다. 흑색소포는 나뭇가지모양으로 7개의 소낭 중앙에 짙게 나타난다. 목덜미에 수개의 나뭇가지모양의 흑색소포가 뚜렷히 관찰되며, 꼬리말단 등쪽 및 배쪽에 짙게 분포한다. 2, 3번째 소낭 사이부터 꼬리 끝까지 체색 중앙의 각 근절에 점모양의 흑색소포가 일렬로 분포한다. 2-5개의 짙은 세로띠 흑색소포가 항문 뒤부터 체측의 배면을 따라 꼬리 끝까지 분포한다(Fig. 2A, 2B).
[Table 1.] Comparisons of measurements and counts for Ophisurus macrorhynchos leptocephali
Comparisons of measurements and counts for Ophisurus macrorhynchos leptocephali
꼬리지느러미 형성기 엽상자어(전장 19.8-39.3 mm): 전장 30 mm 이후 엽상자어부터 전체 근절수는 215개로 성어와 같아진다. 전체 근절수는 215-240개, 항문앞 근절수는 95-97 개이다. 주둥이 선단부의 이빨은 이전보다 다소 짧아진다. 이전보다 위턱이 전방으로 길어지며, 아래턱이 위턱보다 약간 전방으로 돌출되어 있다. 양턱의 이빨 배열식은 으로 이전보다 약간 증가하였다. 두장은 전장의 7.9-11.1%, 체고는 전장의 4.8-7.6%, 항문전장은 전장의 64.4-72.2%로 이전보다 줄어 들었다(Table 1). 꼬리는 이전보다 훨씬 뒤로 신장된다(Fig. 2C). 등지느러미와 뒷지느러미는 막지느러미 형태이다. 흑색소포는 점모양 또는 나뭇가지모양으로 7개의 소낭 등쪽과 중앙에 짙게 나타나며, 꼬리말단의 등쪽, 배쪽에 흔적적으로 분포한다. 이전과 달리 세로띠 흑색소포가 5-7개로 증가하며, 항문 뒤부터 체측의 배면을 따라 꼬리 끝까지 분포한다(Fig. 2C).
꼬리지느러미 발달기 엽상자어(전장 40.5-44.5 mm): 전체 근절수는 211-213개, 등지느러미앞 근절수는 105-115개, 항문앞 근절수는 71-101개이다. 양턱의 이빨 배열식은 으로 이전보다 증가하였다. 양턱의 길이는 거의 비슷하나 아래턱이 위턱보다 약간 전방으로 돌출되어 있다(Fig. 2D). 두장은 전장의 7.0-7.7%, 항문전장은 전장의 62.5%로 이전보다 약간 줄어 들었으며, 체고는 이전과 유사하였다(Table 1). 등지느러 앞길이는 전장의 63.0-71.5%로 몸의 중앙보다 약간 뒤쪽에 위치하였다. 이 시기에는 이전과 달리 모든 지느러미가 분화중인데, 등지느러미는 105-115번째 근절 아래에서 시작하여 꼬리끝까지 이어진다. 뒷지느러미와 꼬리지느러미는 잘 발달하였으며, 꼬리말단에 하미축골이 관찰된다. 6개의 짙은 세로띠 흑색소포가 항문 뒤부터 체측의 배면을 따라 꼬리 끝까지 분포한다(Fig. 2D).
비고: 바다뱀 엽상자어는 변태 전 바다뱀아과의 특징인 두개의 간엽이 서로 연결되어 있고, 담낭은 두번째 간엽(소낭)에 위치하는 점에서 일치하였다. 또한, 전장 30.0 mm 이상 엽상자어의 전체 근절수는 211-215개로, 동아시아에 분포하는 바다뱀과 어류 중 바다뱀의 척추골수(바다뱀과 엽상자어는 211-215개 vs. 성어 바다뱀은 201-210개) 범위에 속하여 성어의 특징과 잘 일치하였다(Hatooka, 2002; Kim et al., 2005; Ji and Kim, 2011).
한편, McCosker (1977)는 바다뱀과의 골격분석을 통한 계통학적 연구에서 바다뱀속은 까치물뱀속과 근연관계를 나타낸다고 보고한 바 있다. 본 연구의 엽상자어기 특징들을 비교하면, 바다뱀 엽상자어는 항문전 소낭수가 7개, 항문 뒤 흑색소포수가 6-7개로 대서양산 및 북동아시아산 까치물뱀속(
또한, 부화직후 엽상자어의 근절수는 성어의 척추골수에 비해 다소 많은 것으로 알려져 있는데(Leiby, 1984), 본 연구에서 바다뱀 부화직후 엽상자어의 경우 전체 근절수가 240개 이상으로 성어의 척수골수 범위(201-210개) 보다 30개 이상의 차이를 보였다(Table 1). 그러나, 양턱의 길이가 같아지는 시기(전장 30.0 mm 이상)가 되면 성어의 척추골수 범위에 들어가는 것으로 사료된다. 한편, 날붕장어 엽상자어 및 까치물뱀속 엽상자어 3종은 양턱 길이가 같아지는 전장 20.0 mm에서 성어의 척추골수와 일치하여, 바다뱀 엽상자어 보다 이른 시기에 성어의 척추골수와 같아 지는 것으로 나타났다(Fahay and Obenchain, 1978; Ji et al., 2011).
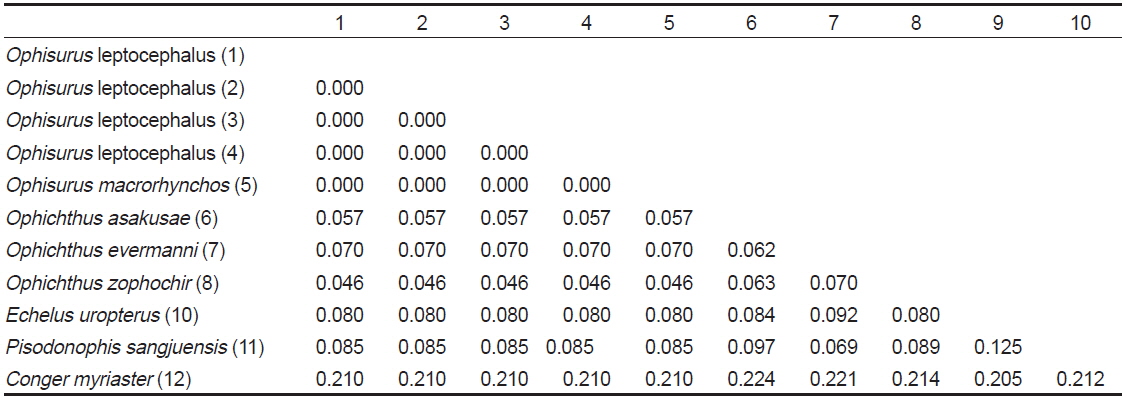
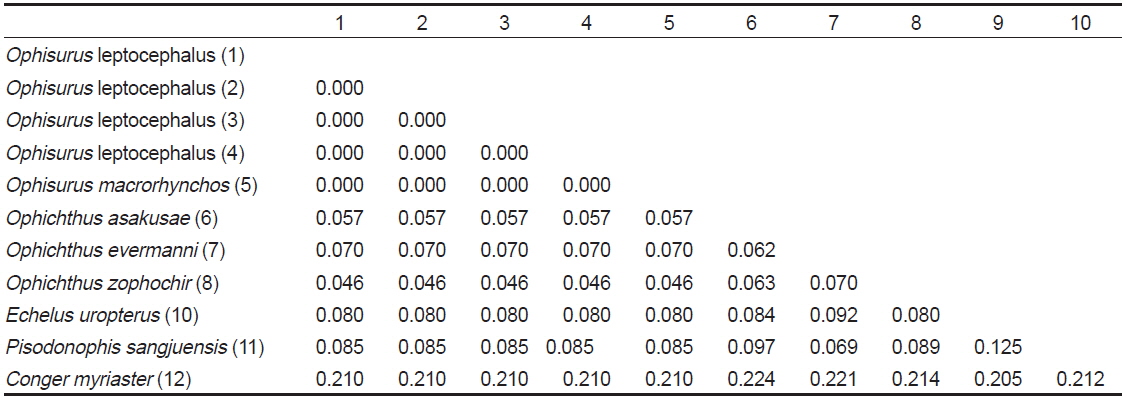
제주도산 바다뱀 엽상자어를 대상으로 미토콘드리아 DNA 12S rRNA 886 base pair 염기서열을 분석한 결과, 우리나라 남해에서 채집된 바다뱀(
[Table 2.] Genetic distance among Ophisurus leptocephali, 6 ophichthid species and 1 outgroup
Genetic distance among Ophisurus leptocephali, 6 ophichthid species and 1 outgroup
본 조사지역에서 바다뱀 엽상자어는 8월에 제주도 동남부, 한일중간수역 및 대마도 남부 해역에서 부화직후 추정 엽상자어(전장 9.8-18.0 mm)와 꼬리지느러미 형성전 엽상자어(전장 19.8-39.3 mm), 꼬리지느러미 발달기 엽상자어(전장 40.5-44.5 mm)가 출현하였다(Fig. 1). 산란장 조사는 어란과 부화자어의 출현으로 규명이 가능하다. 최근 붕장어 부화직후 엽상자어(전장 5.8-7.8 mm)의 출현으로 일본 남부 이주 보닌 마리아나 해구(IBM; Izu-Bonin-Mariana) 해역에서도 산란장이 형성되며 (Kurogi et al., 2012), 은붕장어 2아종(
본 연구결과 바다뱀의 부화직후로 추정되는 엽상자어(전장 9.8-11.3 mm)가 한일 중간수역에서 채집되어 바다뱀의 산란장은 대마난류의 영향이 강한 한국 제주 남동부 해역 또는 일본 큐슈 서부 해역에 형성되는 것으로 사료된다(Fig. 1). 한편, 바다뱀과 2종(