어류 및 패류양식어장의 운영이 저서생태계에 미치는 영향을 파악하기 위하여 다수의 연구들이 수행되었다(Tsutsumi et al., 1991; Park et al., 2000a; Lim et al., 1992; Yokoyama, 2002; Jung et al., 2002, 2007; Carroll et al., 2003; Crawford et al., 2003; Edgar et al., 2005; Yoon et al., 2007). 일반적으로 가두리형 어류양식어장에서는 사료 투입과 주변으로부터 유입된 유기물이 저층에 퇴적됨으로써 해수의 탈산소화를 유발하여 서식생물에게 부정적인 영향을 미친다(Tsutsumi, 1995; Yokoyama et al., 2004; Edgar et al., 2005). 반면 수하식 패류양식 어장에서는 양식생물과 부착생물에서 기원한 분(faeces)과 위분(pseudofaeces)이 저층 퇴적물의 화학적 조성은 물론 대형저서동물의 군집변화를 유발한다는 것이 지배적이다(Uyeno et al., 1970; Crawford et al., 2003). 특히 패류양식업이 활발한 진해만에서는 이미 양식생물과 부착생물의 배설물로부터 유래되는 유기물이 퇴적물 오염의 중요한 요인으로 고려되고 있다 (Cho, 1991). 한편 대형저서동물 군집은 서로 다른 환경교란에 적응내성을 가지는 다양한 종들을 포함하고 있으며, 퇴적물 내부 또는 표면 위를 흐르고 있는 해수와의 영양염 순환(물질순환)에서 매우 중요한 역할을 담당하고 있다(Diaz et al., 2004). 따라서 대형저서동물의 생물다양성, 생물량(개체수 및 생체량)과 군집구조 파악은 해역 전반의 안정성과 건강도 평가에 매우 중요하며, 동시에 환경교란의 정도를 감시하는데 있어 유용한 정보를 제공한다(Parker, 1975; Rosenberg and Nilson, 2005; Wildsmith et al., 2009). 이러한 이유로 해양생태계에서 대형저서동물을 대상으로 하는 다양한 생물 평가지수(biotic index)들이 개발되었다. 이렇듯 생물 평가지수들은 서로 다른 형태의 교란으로부터 생태계의 질적인 상태(quality status)를 평가하고, 감시하는데 효율적인 수단이 되고 있다(Borja et al., 2004). 지금까지 환경교란에 따른 대형저서동물 군집의 변화정도를 파악하기 위하여 단변량인 출현 종수, 개체수 및 생태학적 제 지수 등의 분석과 함께 그래픽 분석을 이용한 방법이 주로 이용되었다(Pearson and Rosenberg, 1978; Gray, 1981; Warwick and Clarke, 1994). 특히 본 연구에서 적용된 AMBI (Borja et al., 2003)와 M-AMBI (Muxika et al., 2007)는 전 세계적으로 무산소와 빈산소해역, 부영양화해역, 영양염 부하해역, 준설 및 유류오염해역과 생활 및 산업오·폐수 유출해역 등과 같이 인위적인 교란을 받는 환경에 대한 감지와 평가능력이 높은 것으로 입증되고 있다(Borja et al., 2009). 우리나라의 경우에는 BPI 지수(Choi et al., 2003)와 ISEP 지수(Yoo et al., 2010)가 정부 발주의 연구사업에서 대표적으로 적용되고 있으나, 일반화 또는 상용화라는 관점에서는 다소 고려해야 할 부분이 있다. 즉 BPI 지수는 분석에 이용되는 개별 생물의 식성에 대한 정보가 아직까지 많이 부족한 실정이며, ISEP 지수는 연구대상 해역의 생물다양성이 매우 낮음에도 불구하고 오히려 양호한 환경상태를 나타내는 경우가 종종 발생하고 있다는 점이다. 한편 대형저서동물은 행동적으로 이동성이 미약하여 급성 또는 만성적인 서식처 교란에 대해서 능동적으로 회피할 수 있는 능력이 제한되어 있고, 상대적으로 긴 수명을 가지고 있어 환경변화를 잘 반영하는 생물지시자로 이용되어 왔다(Dauer et al., 2000). 지금까지 본 연구해역을 포괄적으로 포함하는 진해만에서 수행된 대형저서동물 연구로는 해양오염이 저서동물에 미치는 영향(Hong and Lee, 1983), 빈산소수괴의 발생과 저서동물의 관계(Hong, 1987; Lim et al., 2006; Kim et al., 2012), 양식장 밀집해역에서의 저서동물의 분포(Lim et al., 1992)와 군집구조(Paik and Yun, 2000) 및 저서동물 군집을 통한 환경평가(Park and Yi, 2002; Lim et al., 2007) 등 매우 다양하다. 본 연구는 패류양식어장이 집중적으로 분포하고 있는 진해만 서쪽해역의 원문만과 가조도 인근해역 등을 대상으로 대형저서동물 군집의 춘계와 하계의 생물다양성, 개체수 및 군집구조를 파악하고, 현 시점에서의 건강도를 평가하는데 목적이 있다. 결국 본 연구는 패류양식어장의 운영에 따른 저서생태계의 과학적인 평가와 동시에 효율적인 어장운영을 위한 방안을 마련하는데 기초자료로써 활용이 가능하다는 점에서 의의가 있다.
진해만은 부산광역시 강서구의 가덕수로에서 경상남도 창원시의 행암만, 마산만, 진동만, 통영시의 원문만과 거제시의 고현만 등 총 6개의 소규모 만으로 구성되어 있고, 만은 복잡한 해안선을 가지는 반폐쇄적 형태를 보이고 있다. 진해만의 전체면적은 약 637 km2이며, 수심은 만의 안쪽에서 중심부 해역을 향하면서 5-20 m내외이다. 진해만은 참굴(
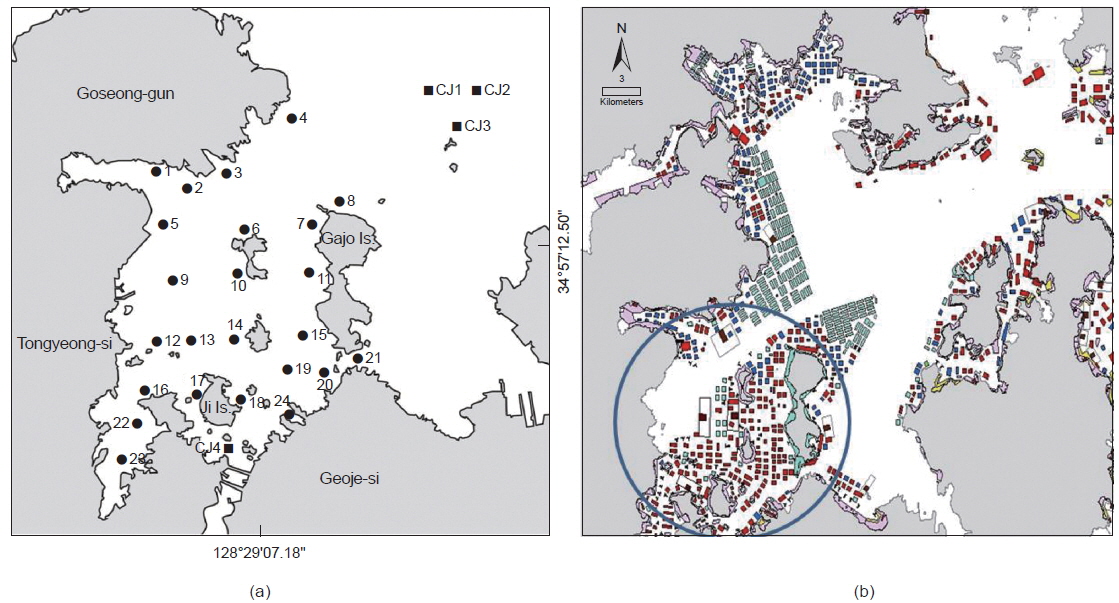
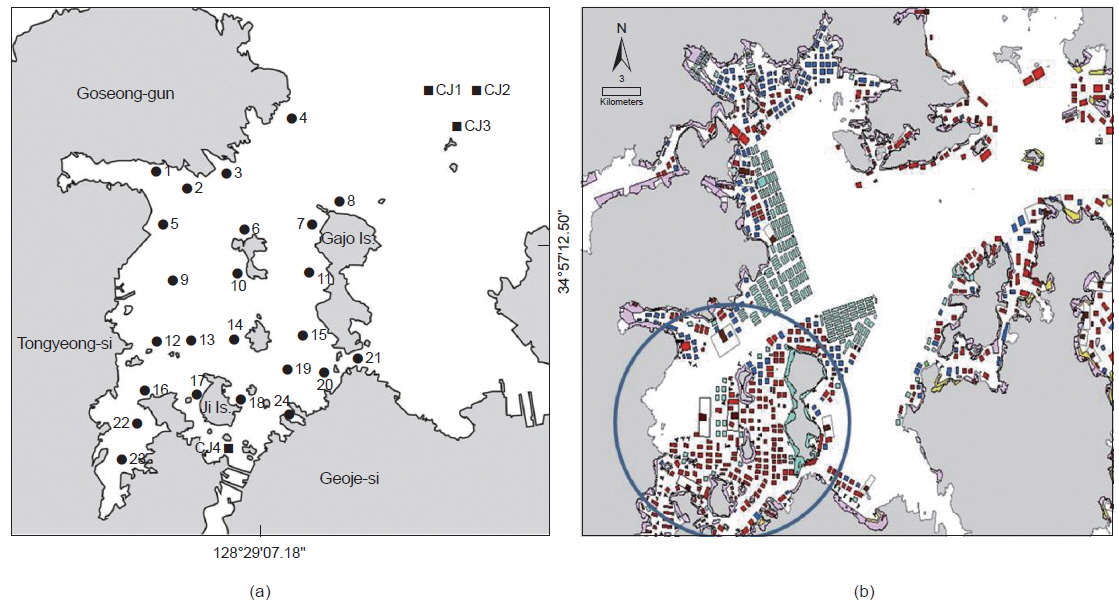
연구해역에 서식하는 대형저서동물의 출현 종수, 개체수, 군집구조 및 건강도 등을 파악하기 위하여 2013년 5월(이하 춘계)과 8월(이하 하계)에 각각 28개 정점을 대상으로 현장조사를 실시하였다(Fig. 1). 이 중 정점 CJ는 패류양식어장이 시설되어 있지 않은 정점에 해당한다. 한편 대형저서동물의 채집은 채집면적이 0.1 m2인 채니기(van Veen grab sampler)를 이용하여 정점 당 2회(0.2 m2)의 퇴적물을 채취하였다. 채취된 퇴적물은 현장에서 1 mm망목의 체를 이용하여 대형저서동물을 분리하였고, 10%의 중성 포르말린으로 고정하여 실험실로 운반하였다. 그 후 분류군별로 선별 및 동정을 실시하였고, 개체수를 계수하였다. 한편 연구해역의 환경요인으로 저층수의 수온, 염분 및 용존산소량은 대형저서동물의 채집과 동시에 다기능수질측정기(YSI 600QS)를 이용하여 측정하였다. 또한 퇴적물의 강열감량(Ignition Loss)은 환경보전도 지수의 산출을 위하여 산휘발성황화물(AVS, Acid Volatile Sulfide) 분석과 동시에 실시하였다. 또한 환경요인은 출현 종수 및 개체수와의 관련성을 파악하고자 SPSS program을 이용하여 상관분석(Spearman rank correlation coefficient)을 실시하였다.
대형저서동물의 군집구조를 파악하기 위하여, 종 다양도 (Shannon and Weaver, 1949)와 균등도(Pielou, 1977) 지수를 구하였다. 한편 출현 종과 개체수의 자료를 이용한 집괴분석은 Bray and Curtis (1957)의 유사도지수(similarity index)를 이용하였고, 유사도 지수 행렬로부터 각 조사 정점과 출현 종을 연결하는 방법으로는 group-average를 적용하였다. 군집분석은 PRIMER (Plymouth Routines Multivariate Ecological Research) computer package를 이용하여 수지도(dendrogram)와 다차원배열법(nMDS ordination)으로 나타내었다(Clarke and Warwick, 2001). 우점종은 출현 개체수를 기초로 선정하였다.
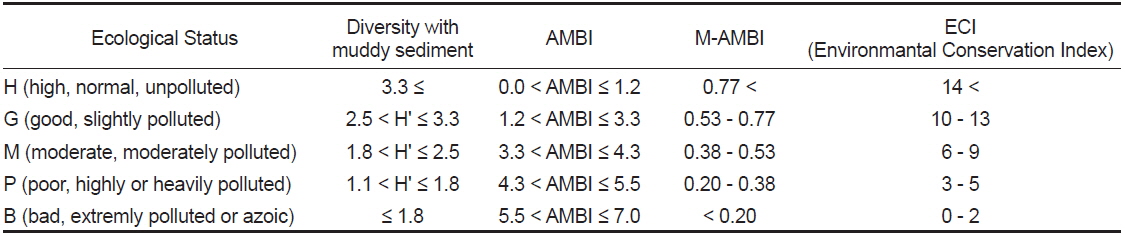
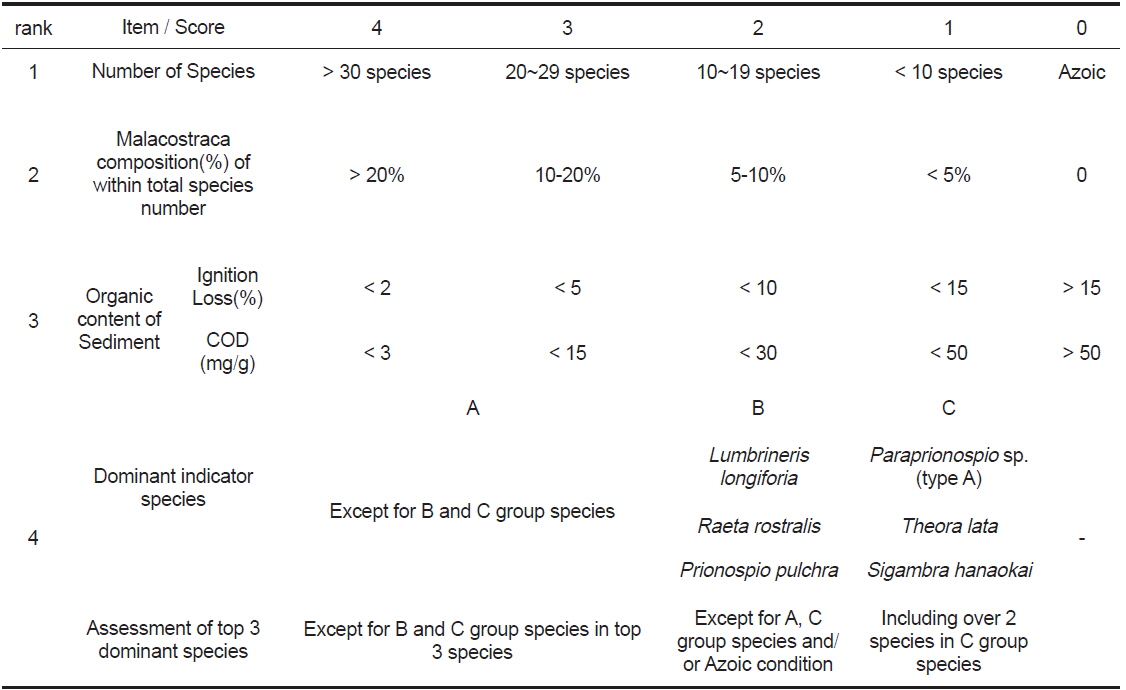
생태계의 건강도는 항상성, 복잡성 및 다양성, 안정성 및 회복성과 생태계 구성인자의 균형성 의 특성을 가지는 건강성을 의미하며, 이러한 특성을 잘 반영하는 건강도 지수가 요구되고 있다(Costanza and Mageau, 1999). 이에 Jorgense et al. (2005)은 건강도 평가 지표의 선정요건으로 생태계의 구조 및 기능과 관련성이 있고, 각각의 항목을 조합하여 통합적이면서 공인된 기준을 갖춘 가이드라인에 의해 선별되어야 함을 제시한 바 있다. 따라서 현재 전 세계적으로 매우 다양한 건강도 지수가 소개되고 있다(Pinto et al., 2009). 한편 본 연구해역에서는 건강도의 평가를 위하여 AMBI (Atzi’s Marine Biotic Index) 및 M-AMBI (Multivariate-AMBI) 지수와 함께 환경보전도 지수(ECI, Ecological Conservation Index, http://www.tokenshikankyo.jp)를 동시에 산출하였다(Table 1 and 2). 이 중 AMBI와 M-AMBI 지수는 AZTI 해양연구소에서 공식적으로 제공하는 software program (http://www.azti.es)을 이용하였다. 또한 일본 동경만 연안을 중심으로 제한적으로 이용되고 있는 환경보전도 지수는 2개 지수와의 비교를 시도하기 위하여 도입하였다. 환경보전도 지수는 평가를 위하여 요구되는 요소가 심도있는 과학적 지식이나 과정을 요구하기 보다는 비교적 일반인도 객관적인 평가가 이루어질 수 있도록 항목의 간소화가 이루어져 있다는 점에서 장점이 있다. 그러나 현재까지 영문(英文)으로 소개되지 않고 있어서 지수명칭과 등급기준의 표기는 임의로 작성하였다.
[Table 1.] Thresholds utilized for each index in Wonmun Bay, on the southern coast of Korea
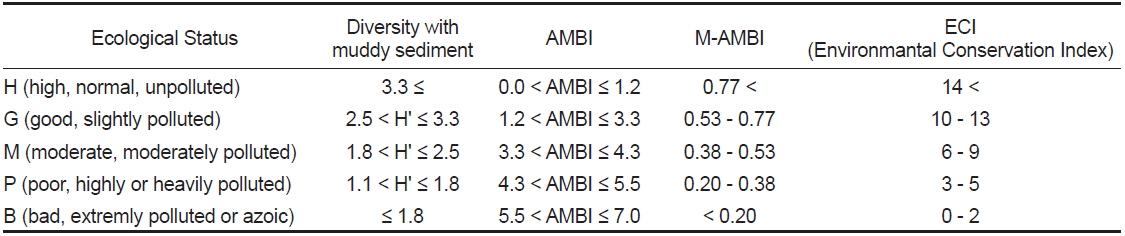
Thresholds utilized for each index in Wonmun Bay, on the southern coast of Korea
[Table 2.] Environmental Conservation Index of using the macrobenthos and sediment organic content
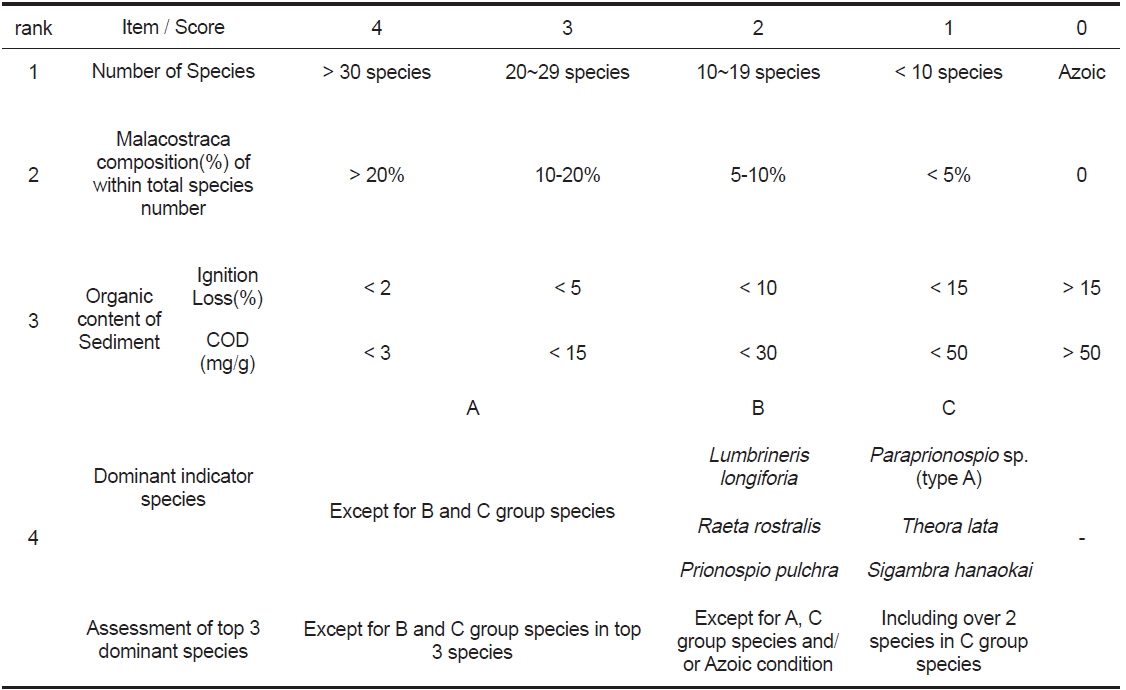
Environmental Conservation Index of using the macrobenthos and sediment organic content
1) AMBI 지수
= [0×%G1+1.5×%GII+3.0×%GIII+4.5×%GIV+6.0× %GV/N] ∴ N = G1+G2+G3+G4+G5
GI = 유기물 증가에 매우 민감한 종[정상환경에서만 출현하며, 특정 육식자와 서관을 가진 식자를 포함함]
GII = 유기물 증가에 둔감한 종[정상환경에서 낮은 밀도를 가지는 종으로, 여과식자, 일반적인 육식자 및 부식자를 포함함]
GIII = 유기물 증가에 내성이 강한 종[표층퇴적물식자로 얼굴 갯지렁이류(Spionidae)등이 포함됨]
GIV = 2차 기회종[표층하퇴적물식자로 실타래갯지렁이류(Cirratulidae) 등이 포함됨]
GV = 1차 기회종[현저한 불균형 환경의 종으로 환원환경의 퇴적물에서 번성하는 퇴적물식자를 포함함]
결과 값은 0-6범위로 0.0-1.2 (가장 양호), 1.2-3.3 (양호), 3.3-4.3 (중간), 4.3-5.5 (불량), 5.5-7.0 (가장 불량)으로 구분할 수 있다. 또한 본 지수에서 = 7은 무생물상태를 나타낸다.
2) M-AMBI 지수
본 지수는 Shannon의 다양도, 풍부도와 AMBI 등의 3개 지수를 통합하여 다변량 분석법의 하나인 인자분석(Factor Analysis)을 수행하여 산출한다. M-AMBI 지수는 1에 가까울수록 최적의 상태인 high status (가장 양호)로, 0에 가까울수록 최악의 상태인 bad status (가장 불량)로 고려되고 있으며, 결과 값은 다양한 형태의 상호검증의 과정을 거쳐 산출되었다. 본 지수는 high (가장 양호), > 0.77; good (양호), 0.53-0.77; moderate (중간), 0.38-0.53; poor (불량), 0.20-0.38, bad (가장 불량), < 0.20로 정의되고 있다(Borja et al., 2009).
3) 환경보전도 지수
환경보전도 지수는 동경만을 공유하는 7개의 지방자치단체가 저질개선대책 수립을 위하여 고안한 평가법이다. 본 지수는 연구해역에서 출현한 대형저서동물의 출현 종수, 전체 출현 종 수 중에서 갑각류의 점유율, 우점지표생물(오염지시종)과 퇴적물의 유기물 함량(강열감량 또는 화학적산소요구량) 등을 종합적으로 고려하여 산출한 값이며, 1995년부터 현재까지 동경도 내만해역의 저질개선 대책을 위한 모니터링의 관점에서 지속적으로 이용되고 있다(www.tokenshi-kankyo.jp). 환경보전도지수는 총 5개 등급으로 구분되며, 이 중 최고등급인 환경보전도 Ⅳ등급(가장 양호)은 다양한 생물이 서식하며, 용존산소량이 풍부한 환경으로 가장 양호하게 보존된 상태를 나타낸다. 다음으로 환경보전도 Ⅲ등급(양호)은 환경은 어느 정도 보전되어 있으나, 하계에 저층수의 용존산소량이 감소하는 등 서식환경이 일시적으로 악화된 상태를 반영한다. 환경보전도 Ⅱ등급(중간)은 퇴적물의 유기오염이 진행되어 빈산소수역의 가능성이 있으며, 저서생물은 오염에 내성을 가진 종이 우점한다. 환경보전도 Ⅰ등급(불량)은 일시적으로 무산소수역이 되며, 퇴적물은 대부분 검은색을 띠고, 저서생물은 오염에 대한 내성을 가지는 종을 중심으로 구성되는 환경을 나타낸다. 마지막으로 최하등급인 환경보전도 0등급(가장 불량)은 Ⅰ등급과 유사하게 퇴적물은 흑색을 띠면서 용존산소가 거의 없어 저서생물이 서식하지 않으며, 가장 불량한 환경상태를 반영한다. 한편 본 지수는 1)전체 출현 종수가 4종 이하인 경우에는 갑각류의 비율이 높다 하더라도 평점이 1로 부여된다거나 또는 2)전체 출현 종 수가 2종 이하인 경우에는 우점종의 출현에 상관없이 평점을 1로 부과하는 등 제한요인이 있다. 또한 퇴적물의 유기물 함량은 강열감량의 적용을 기본으로 하고 있으나, 이를 측정하지 못했을 경우에는 퇴적물의 화학적산소요구량으로 대체·평가할 것을 권고하고 있다.
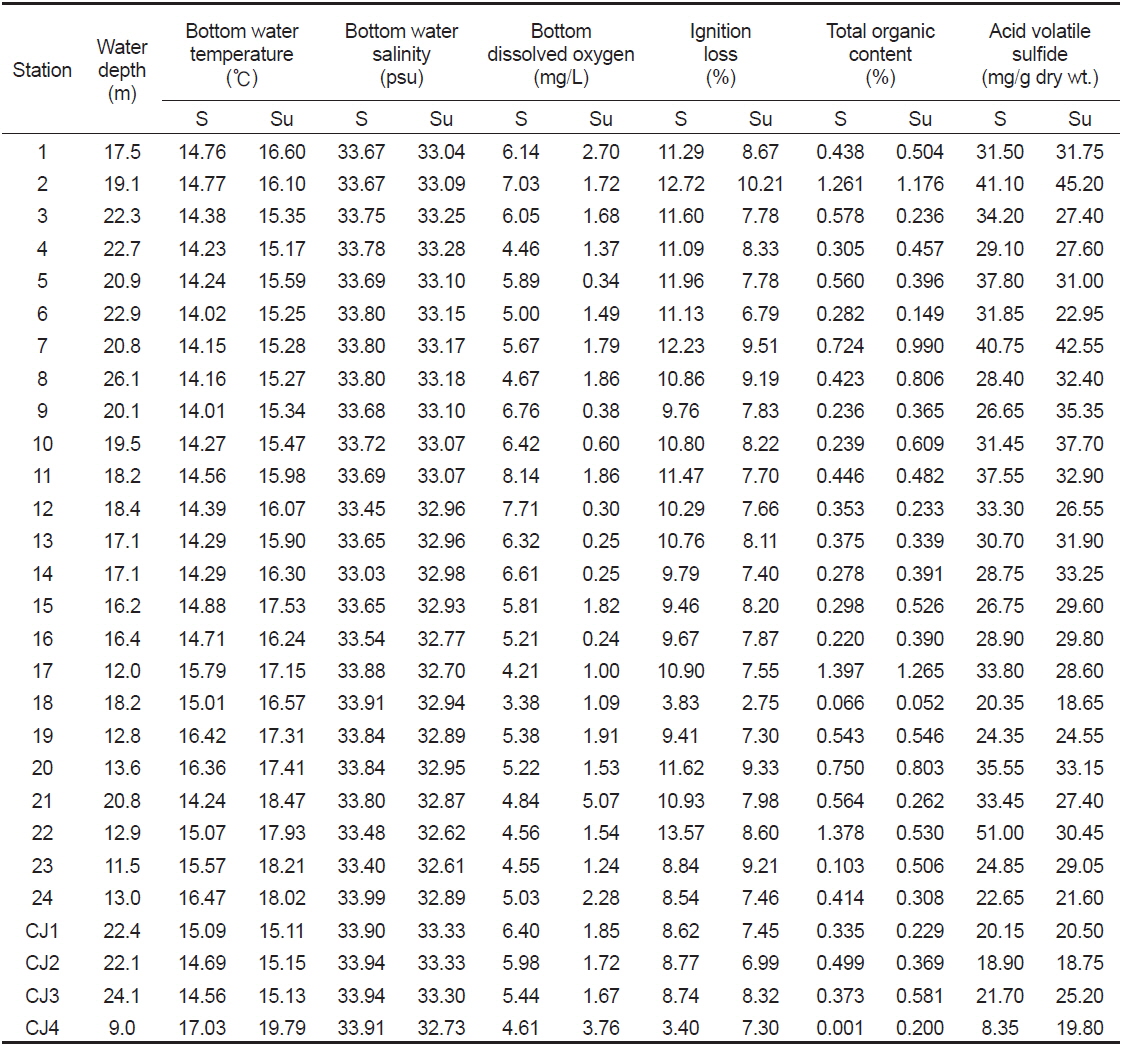
연구해역의 수심은 9.0-26.1 m의 범위에 정점 CJ4에서 가장 얕았고, 정점 8에서 가장 깊었다. 저층수의 수온은 춘계와 하계에 각각 14.01-17.03℃ (평균 14.87±0.83℃)와 15.11-19.79℃ (평균 16.42±1.25℃)의 범위를 나타내었다. 염분은 춘계에 최소 33.03 psu에서 최대 33.99 psu을 나타내었고, 평균 값은 33.72 psu (±0.20) 이었다. 또한 하계에도 32.61-33.33 psu (평균 33.01±0.21)의 범위를 보여 수온과 염분은 조사정점과 시기에 따라 차이가 거의 없었다. 용존산소량은 춘계에 최소 3.38 mg/L에서 최대 8.14 mg/L를 나타내었고, 평균 값은 5.62±1.08 mg/L이었다. 반면 하계에는 0.24-5.07 mg/L (평균 1.55±1.07 mg/L)의 범위를 보여 전반적으로 매우 감소하였다. 특히 정점 21과 CJ4를 제외한 모든 정점에서 빈산소농도인 2.0 mg/L 이하를 나타내었다. 한편 연구해역에서 퇴적물의 유기물 함량(강열감량)은 춘계와 하계에 각각 3.40-13.57% (평균 10.07±2.24%)와 2.75-10.21% (평균 7.91±1.30%)의 범위를 나타내었다. 반면 산휘발성황화물농도는 춘계에 최소 0.001 mg/g dry wt.에서 최대 1.397 mg/g dry wt.를 나타내었고, 평균 값은 0.480±0.353 mg/g dry wt.이었다. 또한 하계에는 최소 0.052 mg/g dry wt.에서 최대 1.265 mg/g dry wt.(평균 0.489±0.292 mg/g dry wt.)의 범위를 보였다(Table 3).
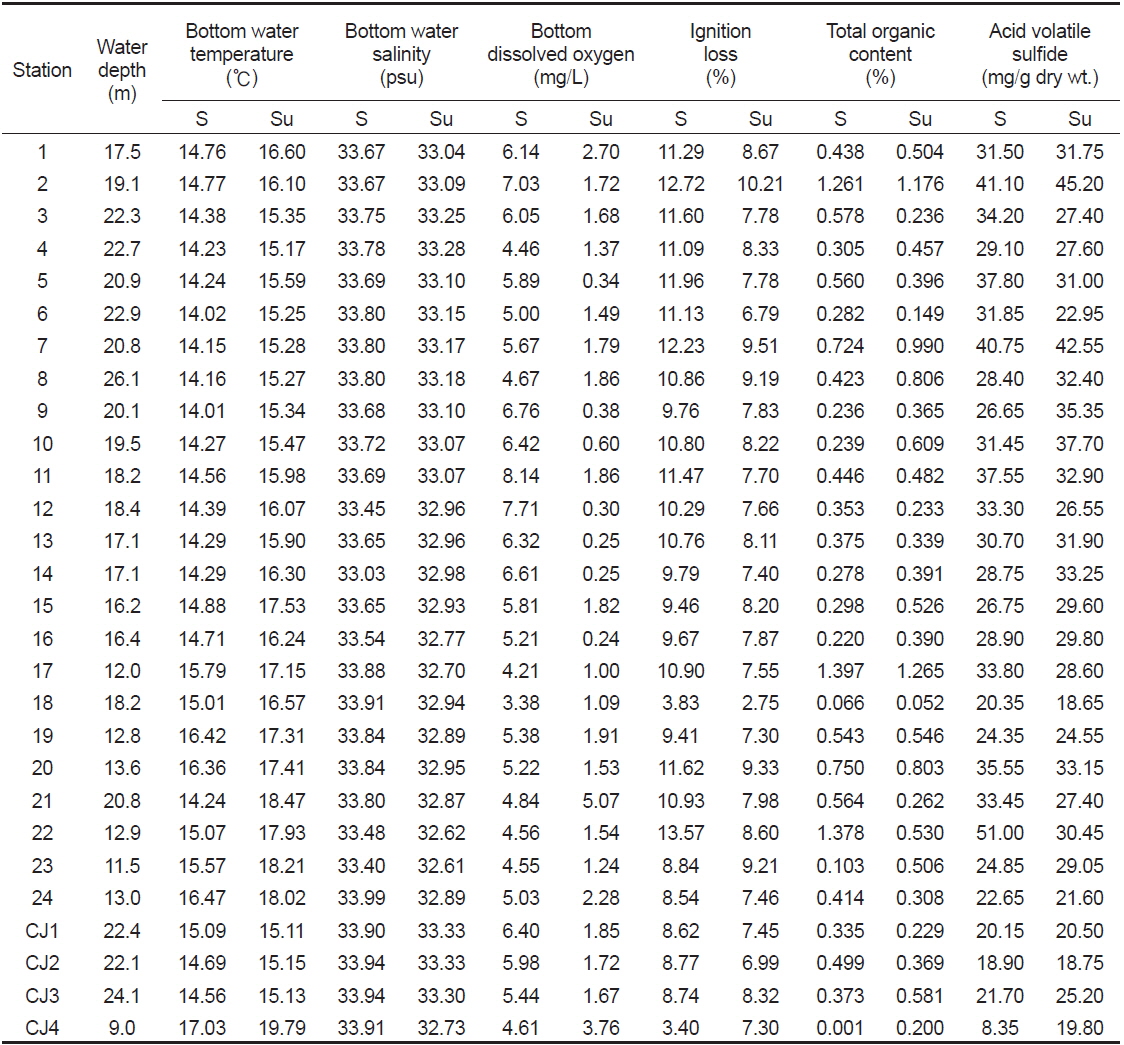
Summary of physical and sedimental parameters at each station in Wonmun Bay, on the southern coast of Korea (S, spring; Su, summer)
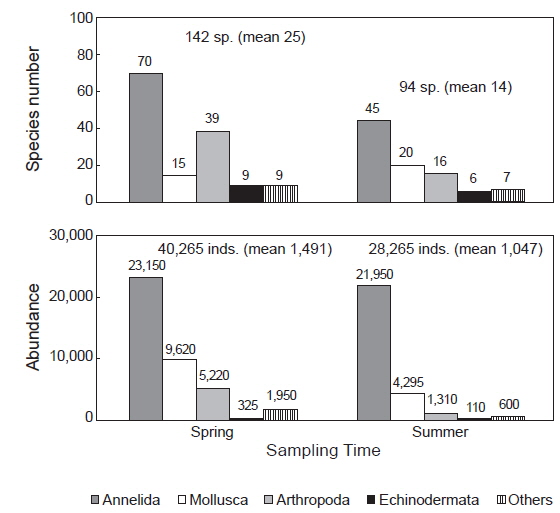
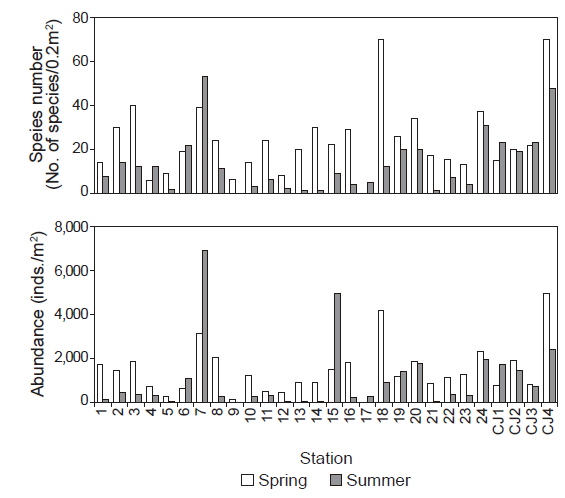
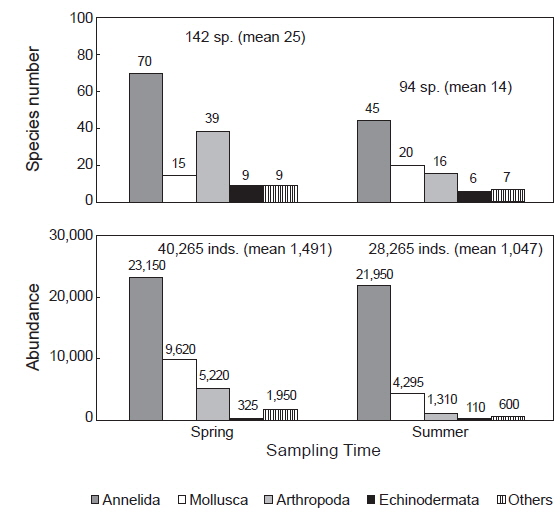
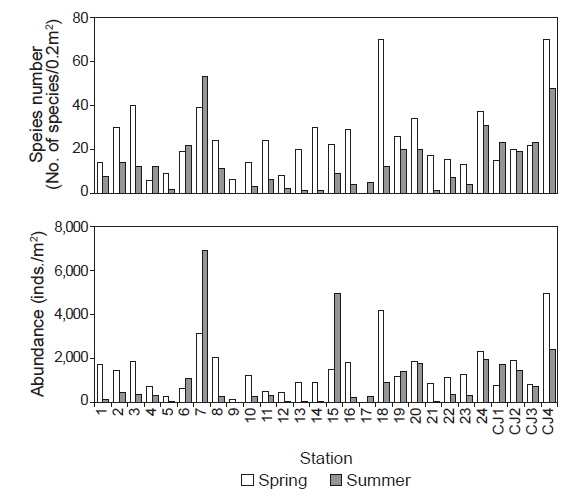
연구해역에서 출현한 대형저서동물은 170종/11.2 m2과 68, 530개체(1,269개체/m2)이었다. 출현 종수는 환형동물의 다모류와 절지동물의 갑각류가 각각 77종과 43종이 채집되어 45.29%와 25.29%의 높은 점유율을 보였고, 다음으로 연체동물은 28종, 16.47%를 차지하였다. 이 밖에 기타동물군과 극피동물은 공통적으로 11종이 채집되어 6.47%의 낮은 점유율을 보였다. 조사시기별 출현 종수는 춘계와 하계에 각각 142종/5.4 m2과 94종/5.4 m2이 출현하여 시간경과에 따라 출현 종수는 크게 감소하였다(Fig. 2). 한편 춘계의 정점 17과 하계의 정점 9에서는 대형저서동물이 출현하지 않은 가운데 조사정점별 출현 종수를 보면, 춘계에는 6-70종의 범위에 정점 4와 9에서 공통적으로 적었고, 정점 18과 CJ4에서 가장 많았다. 이 외에 정점 2, 3, 7, 14, 20 및 24에서 평균 30종 이상이 채집되어 상대적으로 다양하였다. 하계에는 최소 1종(정점 13, 14와 21)에서 최대 53종(정점 7)의 범위를 나타내었다. 이 밖에 정점 24와 CJ4에서 각각 31종과 48종이 출현하여 생물다양성이 상대적으로 높았다(Fig. 3). 개체수에 있어서도 다모류는 45,100개체가 채집되어 65.81%의 높은 점유율을 보였고, 다음으로 갑각류와 연체동물이 각각 13,915개체, 20.30%와 6,350개체, 9.53%를 차지하였다. 이 외에 기타동물군과 극피동물은 각각 2,550개체와 435개체가 채집되어 3.72%와 0.63%로 점유율이 매우 낮았다. 조사시기에 따라서는 춘계와 하계에 각각 40,265개체/27 m2와 28,265개체/27 m2가 출현하여 출현 종수와 동일하게 하계에 밀도가 크게 감소하였다(Fig. 2). 한편 각 조사정점별 출현 개체수를 보면, 춘계에는 125-4,950개체의 범위에 정점 9에서 가장 적었고, 정점 CJ4에서 가장 많았다. 이 외에 정점 7과 18에서 각각 3,125개체와 4,180개체가 출현하여 상대적으로 많았다. 또한 하계에는 최소 5개체(정점 13과 14)에서 최대 6,875개체(정점 7)의 범위를 보였고, 이 밖에 정점 15와 CJ4에서 각각 4,940 개체와 2,400개체의 밀도를 나타내었다(Fig. 3).
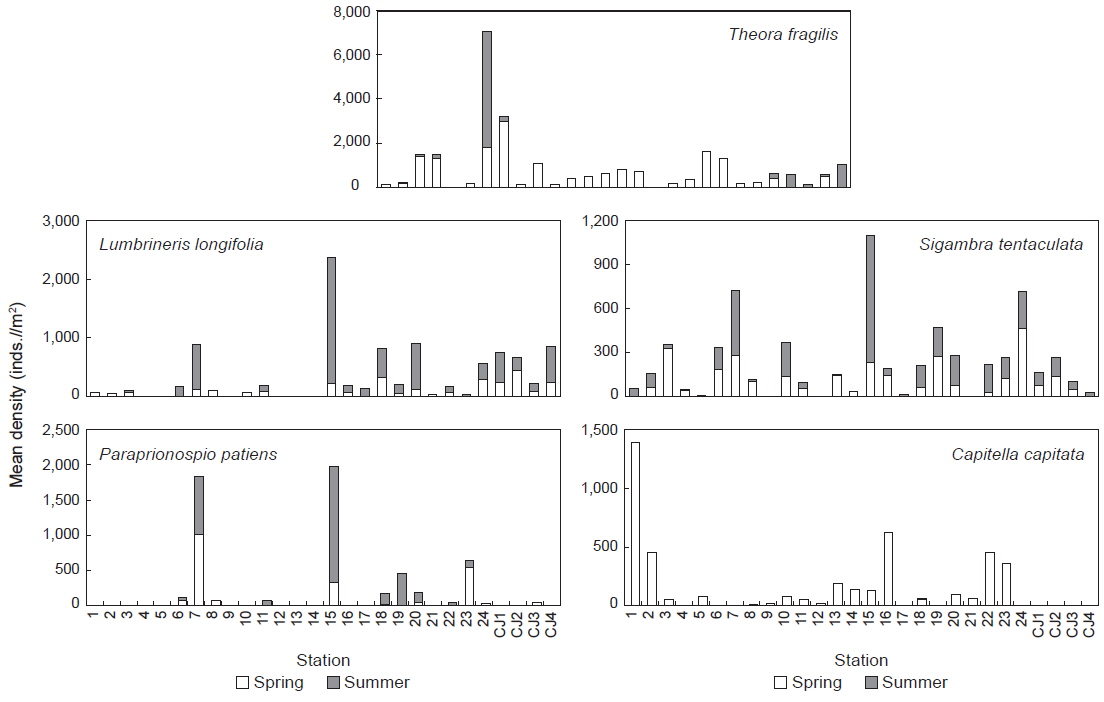
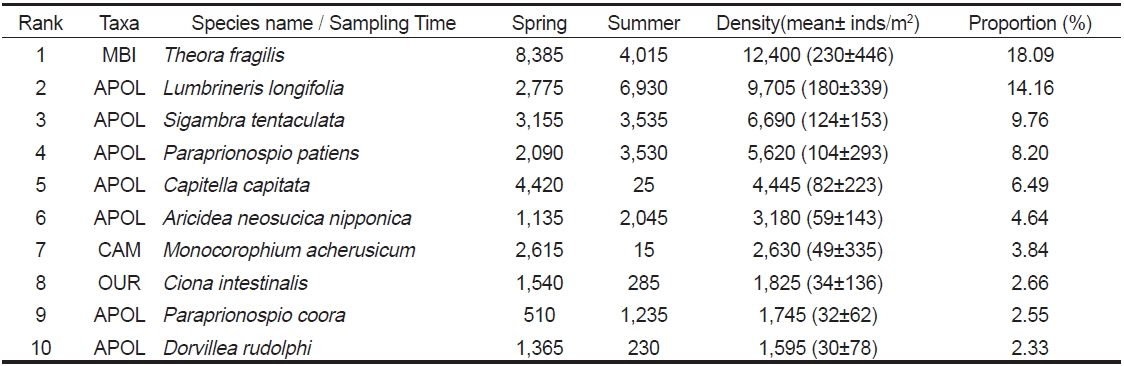
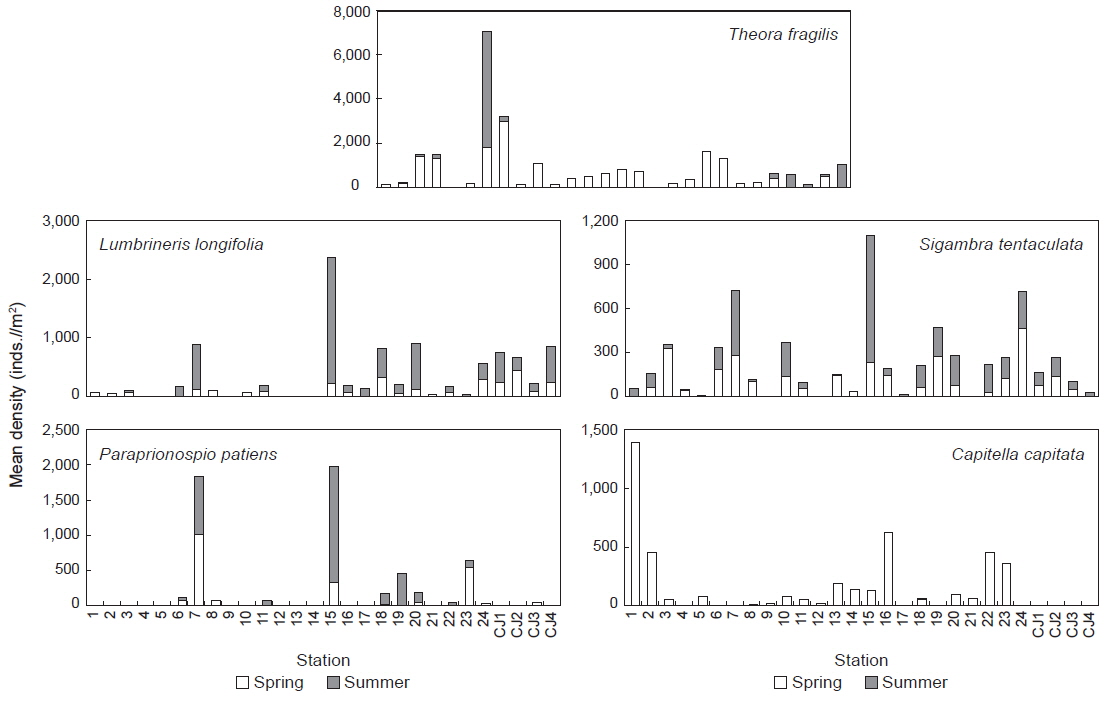
연구해역에서 출현한 대형저서동물의 개체수를 기준으로 상위 우점종을 선정한 결과, 이매패류의 아기반투명조개(
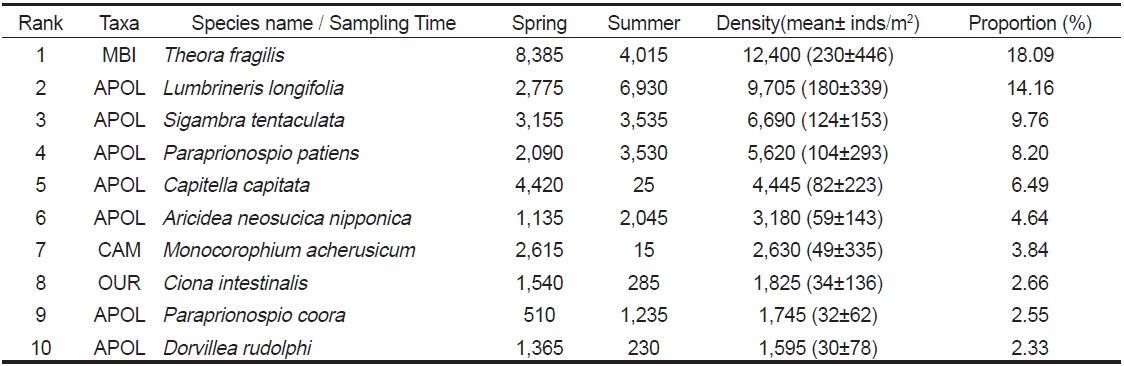
The top 10 dominant species raking based on abundance data in Wonmun Bay, on the southern coast of Korea
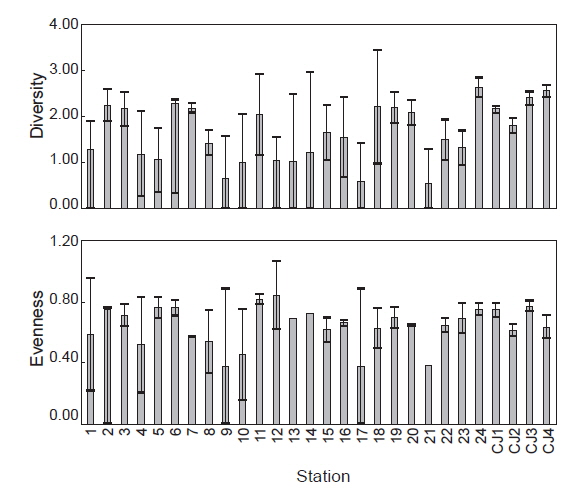
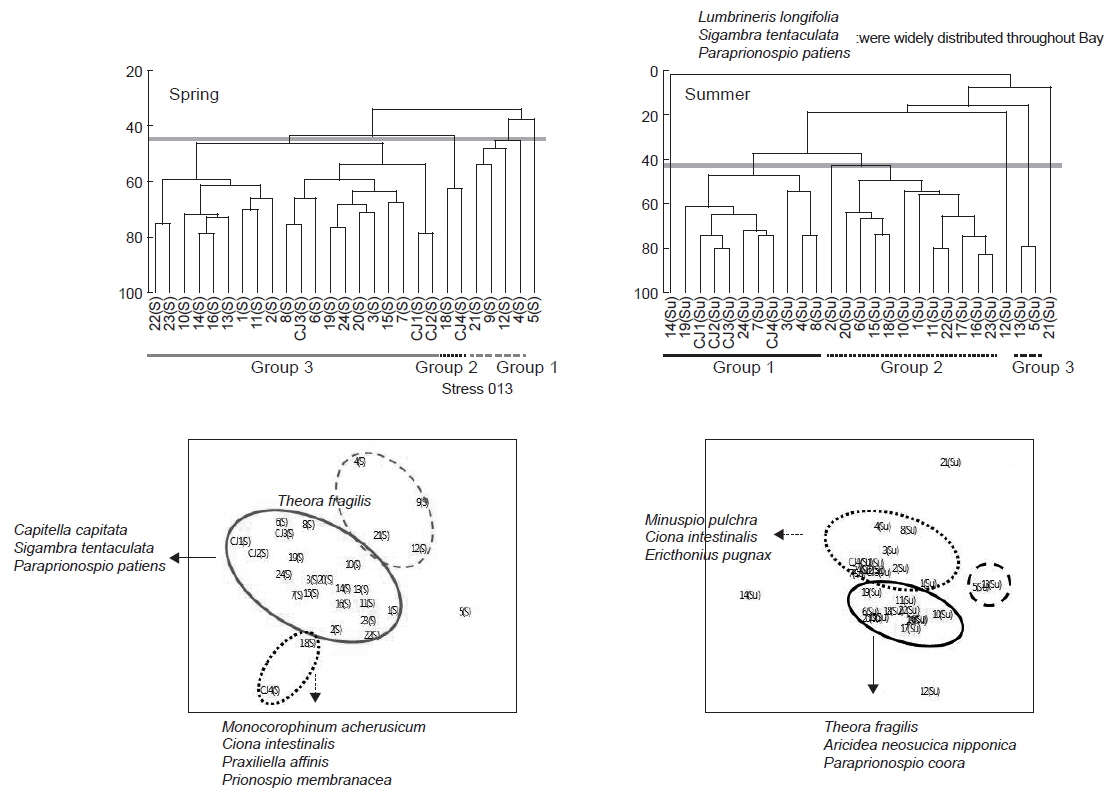
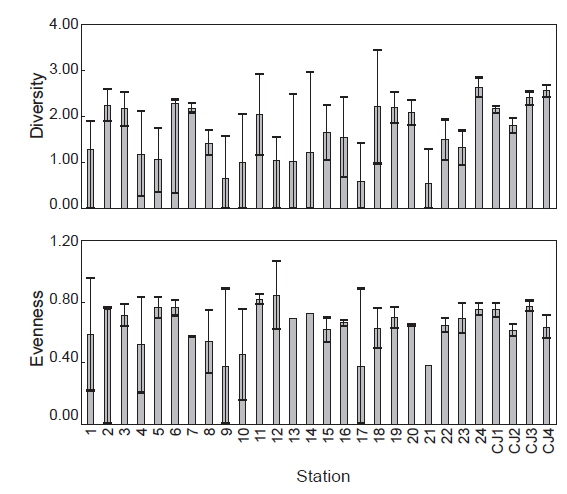
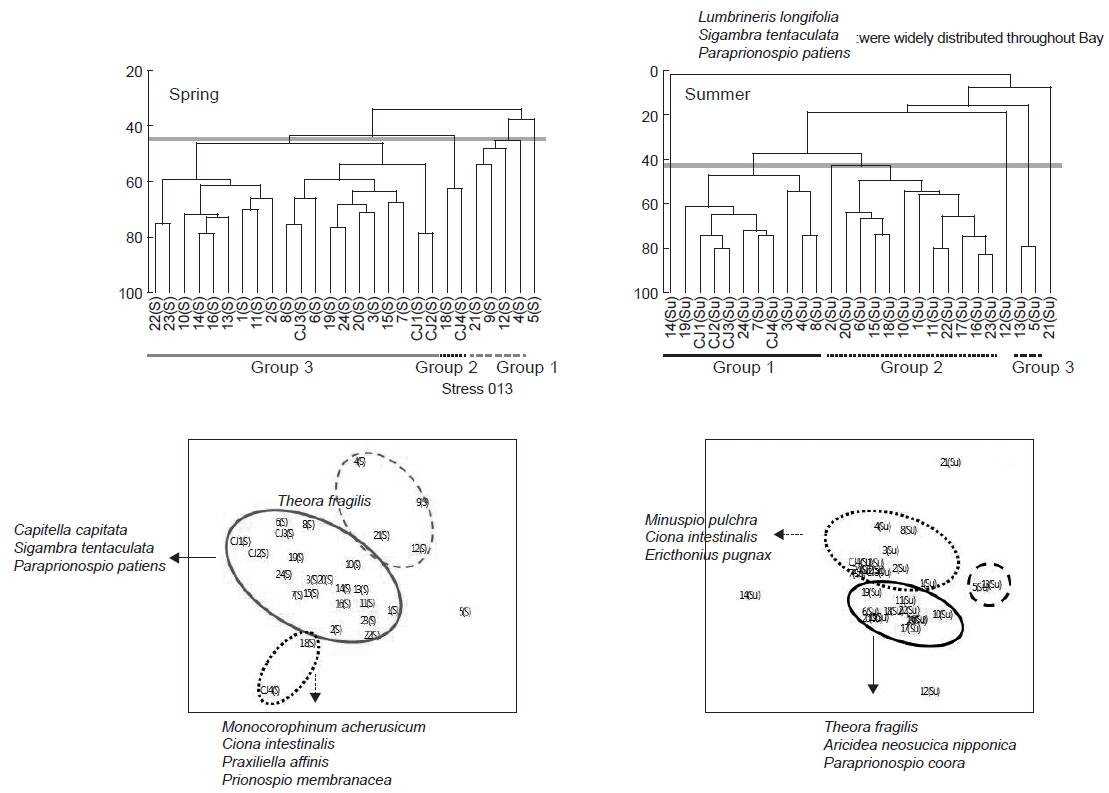
연구해역에서 출현한 대형저서동물의 개체수 자료를 기초로 생태학적 제 지수를 구하였다. 종 다양도는 0.26-3.09의 범위에 평균 값은 1.65이었다. 하계의 정점 10에서 가장 낮았고, 춘계의 정점 18에서 가장 높았다. 한편 조사시기에 따른 평균 다양도는 춘계와 하계에 각각 1.91과 1.39의 값을 나타내 하계에 크게 감소하였다. 조사정점에 따른 평균 다양도는 0.54-2.64의 범위에 정점 21에서 가장 낮았고, 정점 24에서 가장 높았다(Fig. 5). 균등도는 0.29-1.00의 범위에 평균 값은 0.64이었다. 춘계의 정점 4에서 가장 낮았고, 하계의 정점 12에서 가장 높았다. 그러나 하계의 정점 12는 2종이 동일한 밀도로 출현하여 나타난 결과로 이를 제외하면, 춘계의 정점 11과 하계의 정점 1에서 가장 높은 값을 나타내었다. 조사정점에 따른 평균 균등도는 0.37-0.84의 범위에 정점 12에서 가장 높았고, 정점 9와 17에서 공통적으로 낮았다. 이 외에 정점 10과 21에서도 각각 0.45와 0.38의 수치를 나타내 상대적으로 낮았다. 이렇듯 일부 정점에서의 낮은 균등도 수치는 정점 9, 10과 21에서는 아기반투명조개가, 정점 17에서는 긴자락송곳갯지렁이가 높은 밀도로 출현하였기 때문이었다. 한편 본 연구해역에 서식하는 대형저서동물의 출현 종과 개체수의 자료를 근거로 군집구조 분석을 실시하였다. 분석에서 춘계에는 220개체 이상(출현 밀도의 0.50% 이상) 출현한 28종을 대상으로, 하계에는 160개체 이상(출현 밀도의 0.50% 이상) 출현한 22종을 대상으로 하였고, 우점종에 의한 효과를 줄이기 위하여 원자료를 log(X+1)로 변환하였다. 분석 결과, 춘계에는 크게 3개의 그룹과 1개의 정점(정점 5)으로 구성되었다. 이 가운데 그룹 1은 정점 4, 9, 12 및 21이, 그룹 2는 정점 18과 CJ4가 그리고, 그룹 3은 이들을 제외한 모든 정점들이 포함되었다. 한편 그룹 1과 3에서는 공통적으로 아기반투명조개가 우점한 가운데 이를 제외한 각 그룹별 대표종을 보면, 그룹 1에서는 특별히 대표종이 없었던 반면, 그룹 2에서는 옆새우류의
AMBI 지수
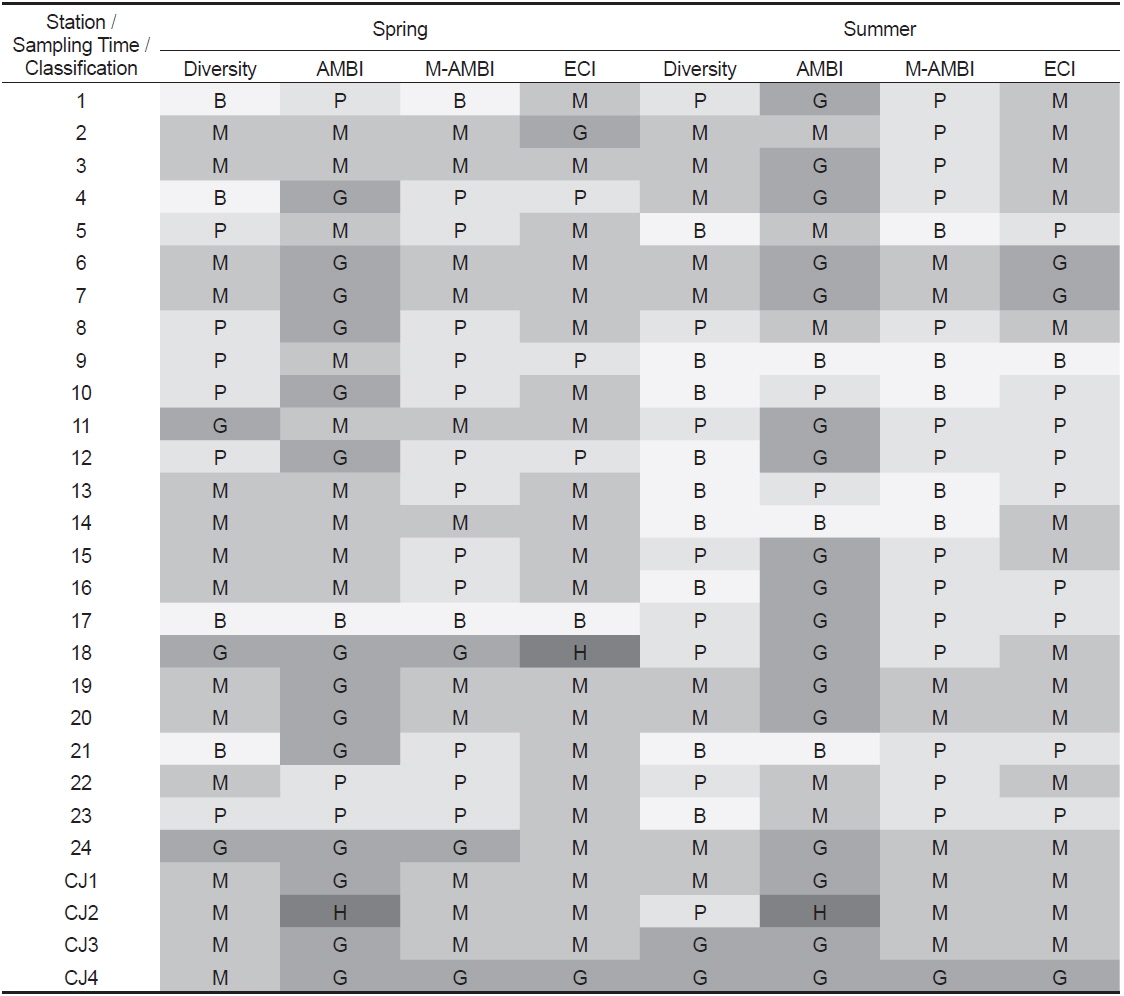
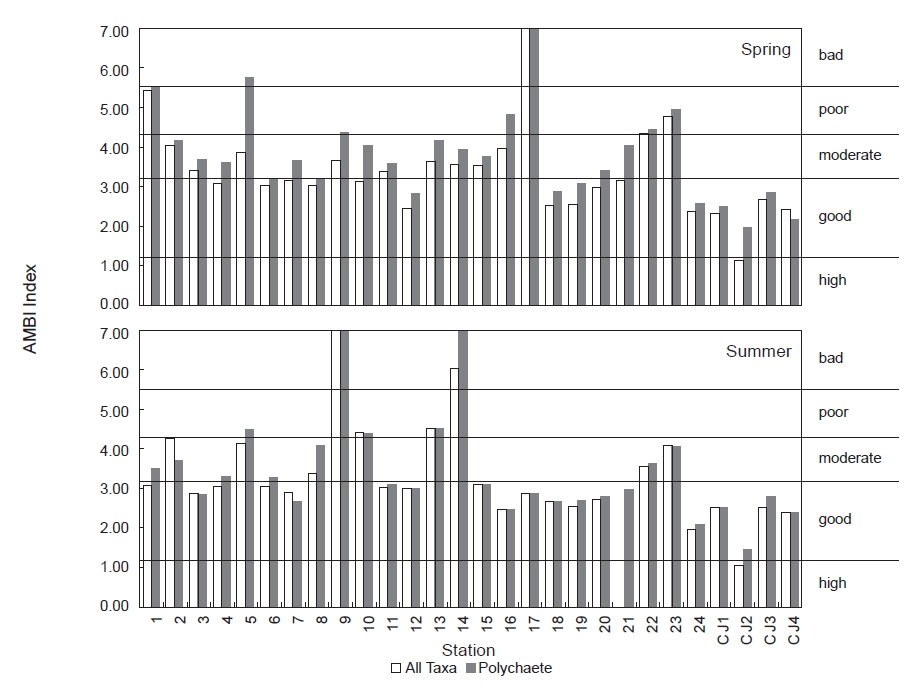
AMBI 지수의 산출에서 유기물에 매우 민감한 종군(I군)이 100%를 차지하여 0.00의 값을 나타낸 하계의 정점 21은 ‘normal(가장 양호, high status; 이하 high)' 상태를, 생물이 전혀 출현하지 않아 무생물조건인 7.00의 수치를 나타낸 춘계의 정점 17과 하계의 정점 9는 동일하게 ‘extremely polluted (가장 불량, bad status; 이하 bad)' 상태를 나타내었다. 이를 제외한 연구해역의 AMBI 지수는 1.03-6.00의 범위에 하계의 정점 CJ2에서 가장 낮았고, 동일 시기의 정점 14에서 가장 높았다. 한편 AMBI 지수는 춘계에는 유일하게 정점 CJ2에서 ‘high (가장 양호)’상태를, 다음으로 정점 4를 포함한 14개 정점에서 ‘slightly polluted (양호, good status; 이하 good)’상태를 나타내었다. 이외에 정점 2를 비롯한 9개 정점에서는 ‘moderately polluted (중간, moderate status; 이하 moderate)'의 상태를, 마지막으로 정점 1, 22와 23 등의 3개 정점에서는 ‘highly polluted (불량, poor status; 이하 poor)’상태를 반영하고 있었다. 하계에는 정점 21에서 유일하게 ‘high (가장 양호)’상태를, 다음으로 정점 1을 포함한 17개 정점에서 ‘good (양호)’상태를 나타내었다. 이 밖에 정점 2를 비롯하여 5개 정점에서는 ‘moderate (중간)’의 상태를 보였고, 정점 10과 13에서는 ‘poor (불량)’상태를 반영하였다. 마지막으로 정점 9, 14와 21은 ‘bad (가장 불량)’상태를 나타내었다(Table 5). 한편 연구해역에서 출현한 다모류만을 대상으로 실시한 AMBI 지수 분석 결과에서도 소수의 정점(정점 1과 5)에서 차이가 있었으나, 전반적인 수치와 경향성은 모든 분류군을 대상으로 실시한 경우와 동일하였다(Fig. 7; Table 5).
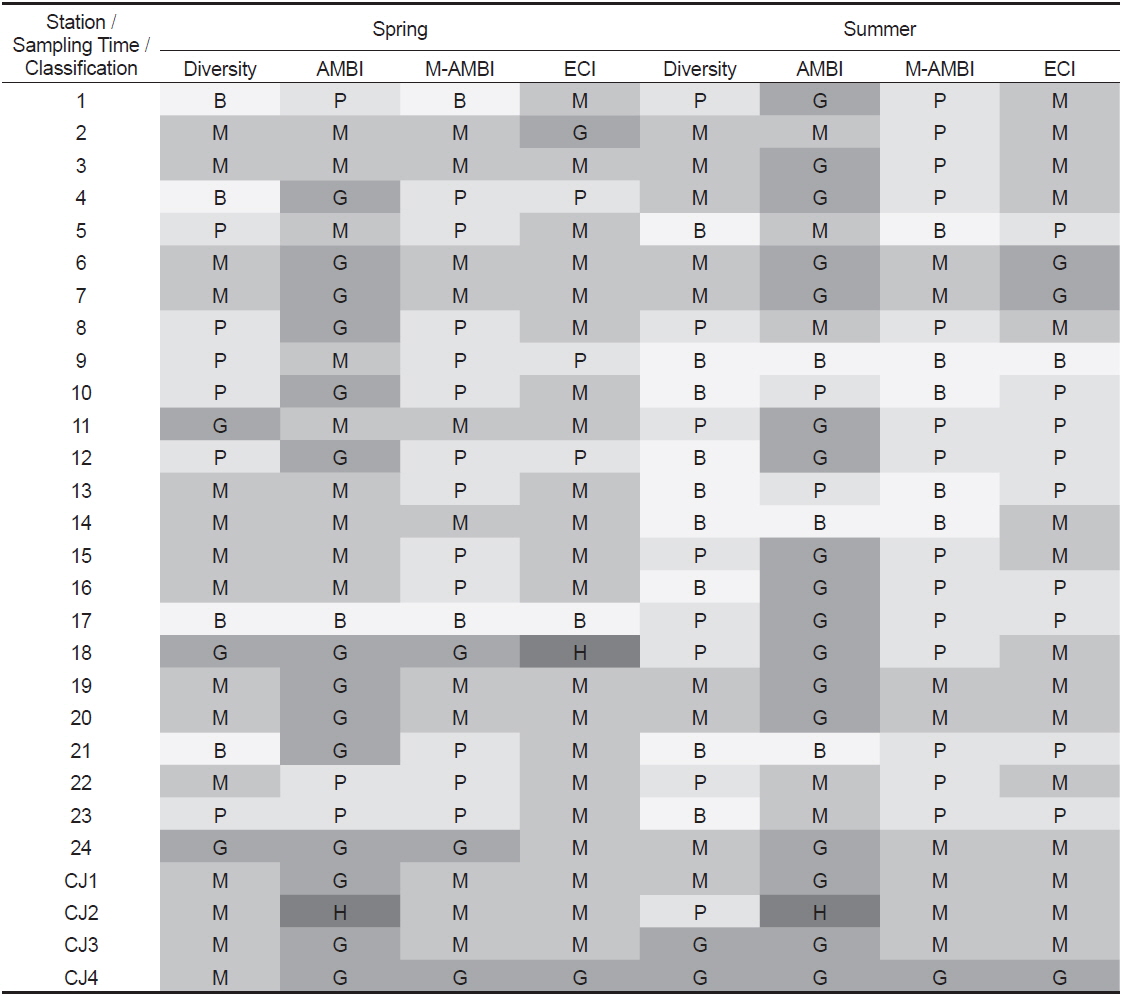
Degree of diversity and three biotic indices (AMBI, M-AMBI and ECI Index) at each station in Wonmun Bay, on the southern coast of Korea (H, high; G, good; M; moderate; P, poor; B, bad status)
M-AMBI 지수
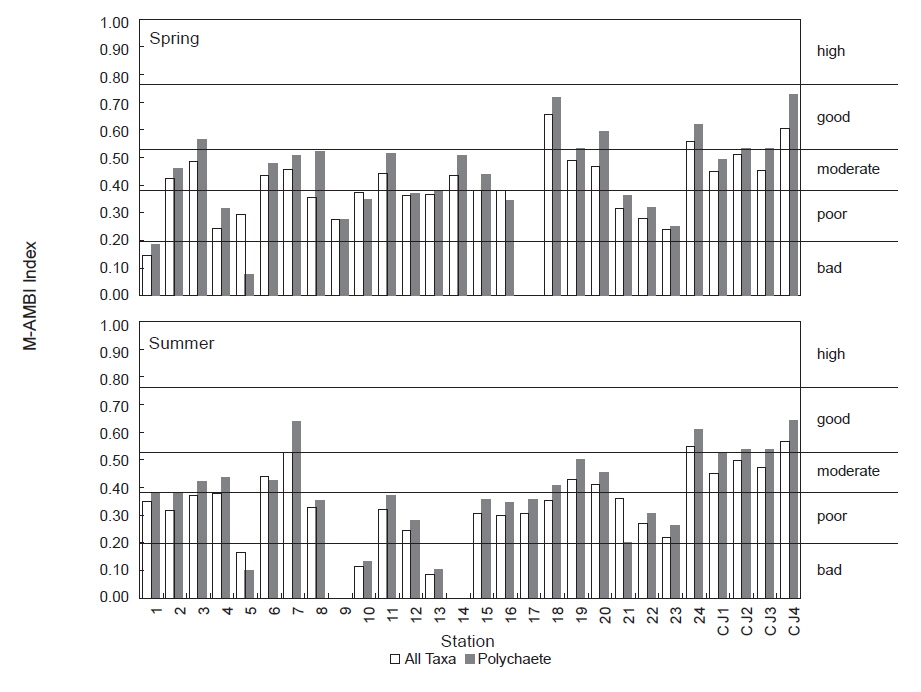
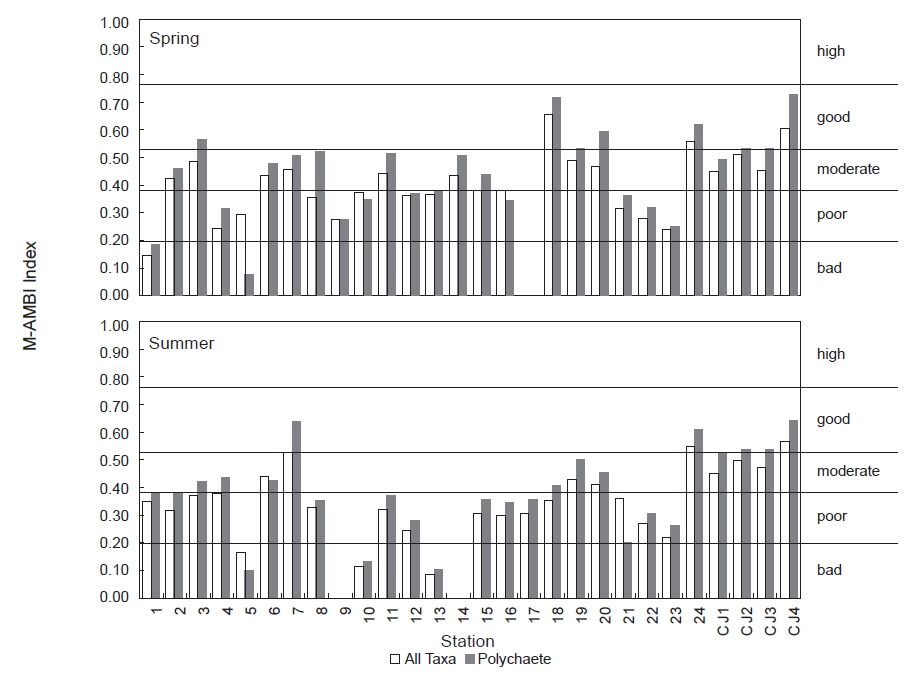
연구해역의 M-AMBI 지수는 0.003-0.660의 범위에 하계의 정점 14에서 가장 낮았고, 춘계의 정점 18에서 가장 높았다. M-AMBI 지수의 산출값을 기초로 해역의 건강도를 평가하면, 춘계에는 가장 좋은 환경상태를 반영하는 ‘high status (가장 양호, 이하 high)’는 없었고, 비교적 양호한 상태를 반영하는 ‘good status (양호, 이하 good)’상태가 3개 정점(정점 18, 24 및 CJ4)이었다. 다음으로 중간단계인 ‘moderate status (중간, 이하 moderate)’상태가 정점 2를 포함한 11개 정점에서, 마지막으로 건강도가 좋지 않은 상태를 반영하는 ‘poor status (불량, 이하 poor)’ 와 ‘bad status (가장 불량, 이하 bad)’상태는 각각 정점 4를 포함한 12개 정점과 정점 1을 포함한 2개 정점이었다. 반면 하계에는 다소 차이를 보여 ‘good (양호)’상태는 정점 CJ4가 유일하였고, ‘moderate (중간)’상태는 정점 6을 포함한 8개 정점이 포함되었다. 마지막으로 열악한 상태를 반영하는 지표인 ‘poor(불량)’와 ‘bad (가장 불량)’는 각각 14개와 5개 정점이었다. 따라서 M-AMBI 지수의 결과로 고려하면, 연구해역은 시간경과에 따라 환경상태가 열악해진 것으로 추정할 수 있었다(Table 5). 한편 다모류만을 대상으로 한 분석에서 춘계에는 정점 5, 8, 15 및 20이, 하계에는 정점 2, 3, 4, 18, CJ2 및 CJ3 등에서 차이가 있었고, 기타 정점에서는 모든 분류군을 대상으로 분석했을 때와 동일하였다. 다만, 이 중에서 춘계의 정점 5는 유일하게 환경상태가 악화된 것으로 나타나 차이가 있었고, 이를 제외한 정점에서는 반대로 향상된 등급을 보였다(Fig. 8; Table 5).
환경보전도 지수
연구해역의 환경보전도 지수는 1-14의 범위에 춘계의 정점 17에서 가장 낮은 값을 나타내었고, 동일 시기의 정점 18에서 가장 높았다. 조사시기별로 보면, 춘계에 0등급(가장 불량, bad status)에는 생물이 출현하지 않은 정점 17이 유일하게 포함되었고, Ⅰ등급(불량, poor status)은 정점 4를 포함한 3개 정점에서, Ⅱ등급(중간, moderate status)은 정점 1을 비롯한 21개 정점에서, Ⅲ등급(양호, good status)과 최고 등급인 Ⅳ등급(가장 양호, high status)은 각각 2개(정점 2와 CJ4)와 1개 정점(정점 18)으로 구성되었다. 반면 하계에는 0등급이 정점 9에서 유일하였고, 다음으로 Ⅰ등급은 정점 5를 포함한 9개 정점이, Ⅱ등급은 정점 1을 포함한 15개 정점이, 그리고 Ⅲ등급은 3개 정점 (정점 6, 7 및 CH4)이 포함되었다. 반면 최고 등급인 Ⅳ등급은 없었다. 따라서 환경보전도 지수는 시간경과에 따라 열악한 환경상태를 반영하는 Ⅰ등급과 Ⅱ등급의 정점은 증가한 반면, 양호한 환경상태를 나타내는 Ⅲ등급과 Ⅳ등급의 정점은 감소하였다(Table 5).
해양생태계에서 연성기질에 서식하는 대형저서동물 군집에게 서식처의 기질로서 작용하는 퇴적물의 조성은 행동, 먹이섭취 및 분포양상에 지대한 영향을 미치는 중요한 환경요인이다(Rhoads and Young, 1970; Gray, 1974). 그럼에도 불구하고, 본 연구에서는 퇴적물의 평균입도, 모래 및 펄함량과 분급도 등은 분석에서 제외하였다. 왜냐하면 본 연구해역은 과거로부터 퇴적상이 대형저서동물의 분포에 영향을 미치는 요인으로 고려되지 않고 있기 때문이다(Lim et al., 1992). 또한 지금까지도 진해만은 만 입구를 제외한 대부분의 해역이 실트성 점토질(silty clay와 clay)로 구성되어 있고, 특히 연구해역이 속한 서쪽해역은 매우 세립한 퇴적물이 지속적으로 유지되고 있기 때문이다(Lim et al., 1992). 비교적 최근에 수행된 Lim et al. (2007)의 결과에서도 모래가 많은 해역이 일부 존재하나, 대부분의 표층퇴적물은 세립질의 silt와 clay의 함량이 각각 1.4-45.7%와 34.3-95.9%의 범위를 나타내 과거와 차이가 없어 이를 입증하고 있다. 다음으로 본 연구해역에서 저층수의 수온과 염분은 춘계와 하계에 각각 평균 14.87℃, 16.42℃와 32.62 psu, 31.49 psu를 나타내 전자는 계절변화를 나타낸 반면, 후자는 하계의 강수량 증가에 따라 다소 감소하는 결과를 보였다. 그러나 용존산소량은 춘계와 하계에 각각 평균 8.43 mg/L과 5.80 mg/L의 범위를 보여 크게 차이가 있었다. 특히 춘계와 비교해 하계에는 정점 1, 21, 24와 CJ4를 제외한 모든 정점에서 빈산소농도에 해당하는 2.00 mg/L 이하의 수치를 나타내고 있다. 한편 패류양식어장이 집중적으로 분포한 해역에서 퇴적물내의 높은 유기물 함량은 미생물 활동을 촉진시켜 결과적으로 산소소비를 유도하고, 특히 조류가 비교적 약한 해역에서는 생물기원의 분이나 위분이 퇴적물 위에 침강하여 쌓임으로써 저층의 산소소비를 보다 가속시키는 것으로 알려져 있다(Yang and Hong, 1988). 그 결과 저층수의 용존산소량이 빈산소농도에 이르면 대형저서동물은 폐사하기 시작하고, 그 이하의 상태에서는 개체군의 적응내성이 뛰어난 소수의 기회종만이 살아남게 되어 매우 빈약한 생물상을 보이게 된다(Pearson and Rosenberg, 1978). 본 연구해역에서도 빈산소수괴가 전역에서 발생한 하계에 출현 종수와 개체수의 감소가 공통적으로 관찰되었고, 기회종으로 분류되고 있는 일부 종들(긴자락송곳갯지렁이, 투구갯지렁이,
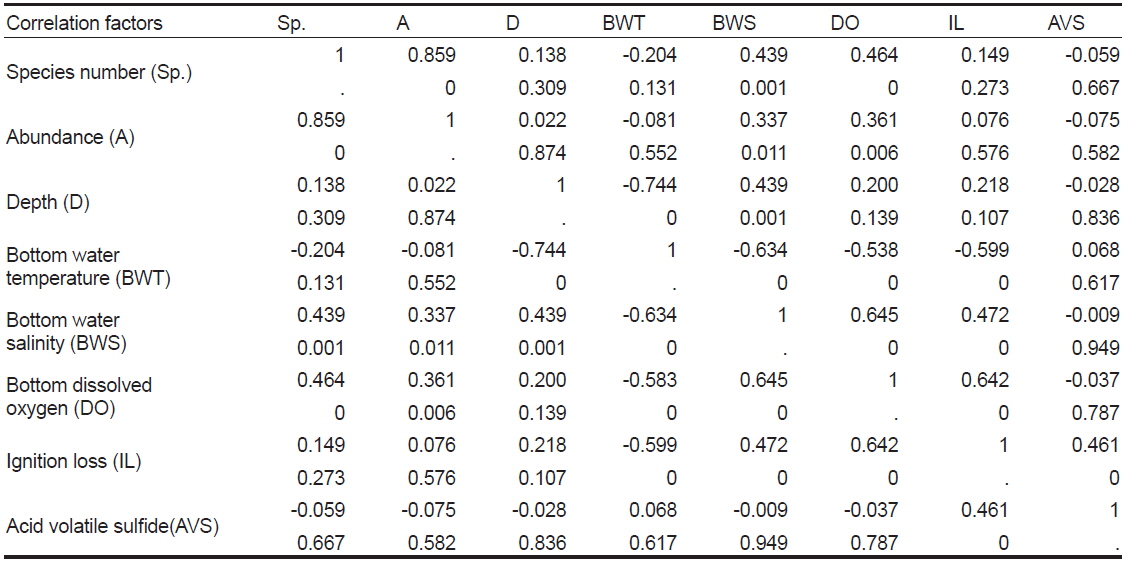
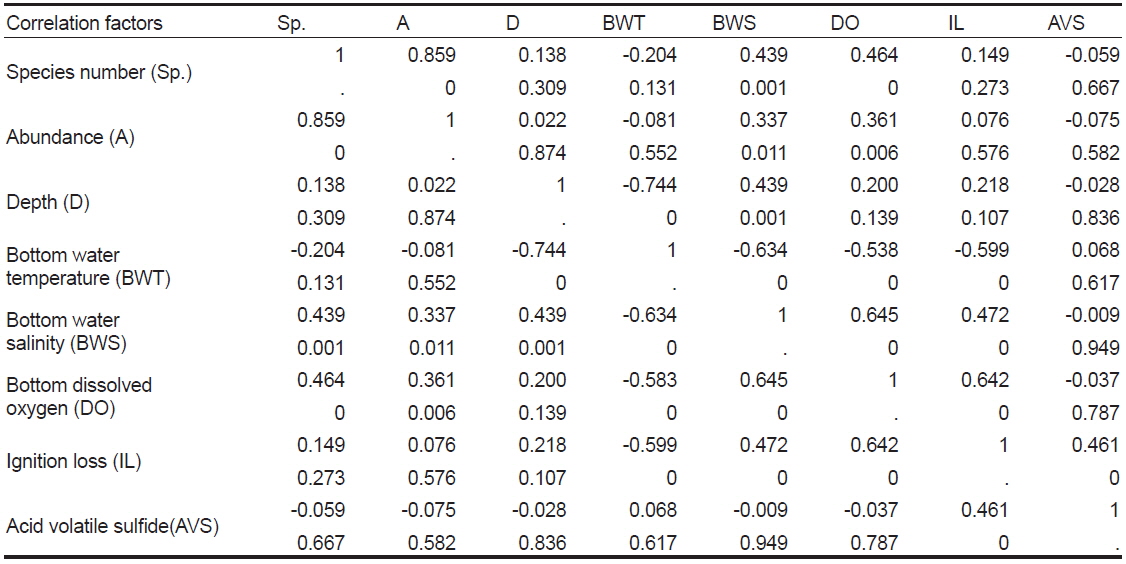
Spearman rank correlation coefficients between abiotic and biotic factors in Wonmun Bay, on the southern coast of Korea
본 연구해역에서 출현한 대형저서동물은 총 170종/11.2 m2에 단위면적당(m2) 개체수는 1,269개체이었다. 본 연구에서와 같이 동일한 해역 또는 서로 다른 해역을 대상으로 하는 출현 종수 및 개체수의 비교는 대상해역의 물리환경 특성(퇴적물 및 수질 환경), 채집환경 특성(면적, 시기, 장소 및 반복의 정도 등) 및 종 동정 수준의 정도에 따라 차이가 발생함으로 무의미한 결과를 도출할 가능성이 있다. 이에 본 연구에서는 채집면적이 상이한 경우에 출현 종수의 정량적 비교를 위하여 적용되는 Whittaker (1975)의 방법을 도입하였다. 본 연구에서 출현 종수와 Whittaker의 수치는 각각 170종과 162로 동일한 해역에서 수행된 Hong and Lee (1983)의 65종과 108 및 Lim et al. (1992)의 107종과 90보다는 많았거나 또는 높았다. 그러나 Kim et al. (2012)의 109종과 263, Lim and Hong (1997)의 287종과 136, Paik and Yun (2000)의 237종과 241 및 Lim et al. (2007)의 225종과 200보다는 적거나 또는 낮았다. 이렇듯 동일해역에서 수행된 과거 연구에서 제시되는 높은 생물다양성은 채집면적이 상대적으로 크고, 공간적으로도 본 연구에서와 같이 국소적인 해역에 제한된 것이 아니라, 상대적으로 광범위하고 다양한 형태의 서식처를 가지는 진해만의 전역을 대상으로 수행했다는 점에서 그 원인이 있다고 판단된다. 한편 단위 면적당(m2) 개체 수에 있어서는 1,269개체가 출현하여 Kim et al. (2012)의 883 개체, Lim and Hong (1997)의 1,046개체 및 Lim et al. (2007)의 984개체보다는 많았고, Lim et al. (1992)의 0-2,022개체와 Paik and Yun (2000)의 1,939개체보다는 적었다. 지금까지 본 연구해역을 포괄적으로 포함하고 있는 진해만은 만 입구해역을 포함하는 동쪽해역에서 대형저서동물의 출현 종수와 개체 수가 높은 반면, 서쪽해역은 생물상이 매우 빈약한 것으로 알려져 있다(Lim et al., 2007). 그러나 본 연구결과에서 제시된 바와 같이 연구해역인 서쪽해역의 출현 종수와 개체수가 Lim et al. (2007)의 동쪽해역과 비교해 상대적으로 낮다고 볼 수 없어 차이가 있었다. 한편 Hong (1987)은 고현해역과 서쪽해역이 다른 해역과 비교해 각각 0-100개체와 100-1,000개체 내외가 출현하여 상대적으로 밀도가 낮음을 밝힌 바 있다. 그러나 Lim et al. (2007)은 고현해역과 가조도 인근해역의 수로 상에 위치한 정점에서 다모류의 긴자락송곳갯지렁이,
본 연구에서는 아기반투명조개를 포함하여 상대적으로 수명이 짧고, 개체 크기가 소형이면서 서식 밀도가 높은 긴자락송곳갯지렁이, 투구갯지렁이,
지금까지 대형저서동물을 이용한 생물학적 지수들이 서로 다른 형태의 교란으로부터 생태계의 건강도를 평가하고자 개발되었다(Borja et al., 2004). 이들 중에서 AMBI 지수는 개별생물의 환경에 대한 민감도를 이용하여 산출하는 지수로 환경교란의 요인이 달라도 잘 적용되기 때문에 비교적 보편적으로 이용되고 있다(Borja et al., 2003). 특히 본 지수는 다양한 장소(인도양, 발트해, 지중해, 북해 및 중국의 발해만)와 교란인자(무산소와 빈산소, 부영양화, 영양염 부하, 유류오염, 준설 및 양식해역)를 대상으로 서식처의 질적 저하를 감지하는데 있어 유효한 생물학적 지수임이 이미 증명되었다(Muxika et al., 2005; Borja et al., 2009; Cai et al., 2012). 그럼에도 불구하고, AMBI 지수는 대형저서동물 군집의 복잡성과 다양성으로 인해 몇몇 조건하에서 오염과 관련한 환경인자와 대비해서 항상 일관된 값을 보이지 않는다. 특히 하구역에서 가장 안쪽에 위치한 해역이나 또는 출현 종수가 매우 낮은 경우에는 단점으로 작용한다는 것이 지적된 바 있다(Borja and Muxica, 2005). 따라서 최근에는 AMBI 지수 적용에 의해 발생하는 단점을 극복하고, 등급 오분류에 대한 문제를 줄이기 위하여 M-AMBI가 제안되었다. MAMBI 지수는 등급의 산정을 위하여 Shannon의 다양도, 풍부도 및 AMBI 지수 등의 3개 지수를 통합하여 고려하는 방법으로 다변량 분석기법인 인자분석을 통하여 산출된다. 본 연구해역에서 AMBI 지수는 조사시기에 따라 정상 상태인 ‘high (가장 양호)’는 춘계와 하계에 동일하게 1개 정점이 포함되었고, 약간 오염상태인 ‘good (양호)’은 각각 14개와 17개 정점으로 구성되어 시간경과에 따라 3개 정점이 증가하였다. 반면 중간 오염상태인 ‘moderate (중간)’는 9개 정점에서 5개 정점으로 감소하였고, 심한 오염과 매우 심한 오염상태를 반영하는 ‘poor (불량)’ 와 ‘bad (가장 불량)’는 각각 3개와 1개 정점에서 2개와 3개 정점으로 증가하였다. 그러나 연구해역은 ‘high (가장 양호)’에서 ‘moderate (중간)’의 등급을 나타내는 정점이 춘계와 하계에 각각 24개와 23개로 차이가 없었다. 다만 시간경과에 따라 ‘poor (불량)’와 ‘bad (가장 불량)’가 각각 4개와 5개 정점을 나타내 1개 정점이 증가한 수준에 불과하였다. 결국 AMBI 지수는 춘계와 하계에 등급분류에 있어서 큰 차이를 보이지 않았다. 반면 연구해역의 M-AMBI 지수는 춘계에는 건강도가 좋은 환경상태를 반영하는 ‘high (가장 양호)’는 없었고, ‘good (양호)’은 3개 정점이 포함되었다. 다음으로 ‘moderate (중간)’는 11개 정점에서, 마지막으로 건강도가 좋지 않은 상태를 반영하는 ‘poor (불량)’와 ‘bad (가장 불량)’는 각각 12개 정점과 2개 정점이었다. 그러나 하계에 ‘high (가장 양호)’는 1개 정점도 포함되지 않아 동일하였으나, ‘good (양호)’은 1개 정점이, ‘moderate (중간)’는 8개 정점이, 마지막으로 ‘poor (불량)’와 ‘bad (가장 불량)’는 각각 14개와 5개 정점으로 크게 증가하였다. 따라서 M-AMBI 지수를 고려한다면, 연구해역은 시간경과에 따라 환경상태가 악화된 것으로 추정할 수 있었다(Table 5). 한편 본 연구에서는 일부 서식처에 따라 다소 차이는 있을 수 있으나, 우점 생물군인 다모류를 대상으로 AMBI와 M-AMBI 지수를 산출하여 비교를 시도하였다. 다모류는 대형저서동물 군집에서 출현 종수나 개체수에서 가장 우점하는 생물군이며(Fauchald and Jumars, 1979). 환경교란에 반응하는 능력과 영양단계의 유연성으로 인해서 연성기질에서의 환경변화를 잘 반영하기 때문이다(Tomassetti and Posello, 2005; Jung et al., 2013). 또한 다모류는 퇴적물과 수환경 경계에서 주로 서식하기 때문에 영양염 순환에 지대한 영향을 미칠 뿐만 아니라, 생활사적인 측면에서도 다양한 환경조건에 직면했을 때 능동적으로 회피하는 능력이 미약하여 건강도 평가에 있어서 유용하게 이용되고 있기 때문이다(Borja and Dauer, 2008). 분석 결과, AMBI 지수는 조사시기에 따라 일부 정점(정점 1과 5)에서 차이가 있었으나, 전반적인 수치와 경향성은 모든 분류군을 대상으로 실시한 경우와 동일하였다(Fig. 7). 또한 M-AMBI 지수도 춘계에는 정점 5, 8, 15 및 20 에서, 하계에는 정점 2, 3, 4, 18, CJ2 및 CJ3 등에서 차이를 보였으나, 기타 정점에서는 모든 분류군을 대상으로 분석했을 때와 동일하였다(Fig. 8). 따라서 상기에서와 같이 다모류 군집만을 대상으로 하였을 경우에 일부 결과에서 다소 차이를 보여주고 있으나, 해역의 전반적인 건강도 평가에서 큰 무리는 없는 것으로 추정된다. 결국 건강도 평가에서 일부 특정 생물군(다모류 및 단각류)을 이용한 건강도 평가가 이루어지고 있는 현시점에서 다모류 군집만을 대상으로 하는 분석과 검증도 의미있는 시도로 판단된다(Gomez Gasteira and Dauvin, 2000; Dauvin and Ruellet, 2007). 마지막으로 연구해역에서 환경보전도 지수는 춘계에 0등급(bad)과 Ⅰ등급(poor)은 각각 1개와 3개 정점이, Ⅱ등급(moderate)은 21개 정점이, Ⅲ등급(good)과 Ⅳ등급(high)은 각각 2개와 1개 정점이 포함되었다. 반면 하계에는 0등급과 Ⅰ등급이 각각 1개와 9개 정점으로, Ⅱ등급이 15개 정점으로, 그리고 Ⅲ등급은 3개 정점으로 나타났고, 최고 등급인 Ⅳ등급은 없었다. 따라서 M-AMBI 지수의 경향과 동일하게 시간경과에 따라 건강도는 점진적으로 불량해지는 동일한 결과를 나타내었다(Table 5). 이렇듯 본 지수는 1회성 또는 단기간의 결과만을 나타내는 이화학적인 시험법과는 다르게 장기간에 걸친 생물의 서식환경을 반영한다는 점에서 장점이 있다. 특히 평가에 필요한 항목수를 최소화하고, 생물학적으로도 전문화된 지 식을 지나치게 요구하지 않는다는 점에서 간편하다고 볼 수 있다. 결국 본 연구에서 처음 소개되고 있는 환경보전도 지수는 해역생태계의 건강도 평가를 위해 다른 해역에서 산출된 결과와 다각적인 비교를 통해 그 효율성을 검증해 볼 필요가 있을 것으로 판단된다. 이상에서와 같이 본 연구에서 적용된 3개의 건강도 지수와 다양도 등을 종합적으로 고려한 결과, 연구해역은 3개의 건강도 평가 지수와 비교해 AMBI 지수에서 다소 차이를 보이고 있으나, 전반적인 경향은 춘계에서부터 시간경과에 따라 점진적으로 건강도가 불량해지는 것으로 파악할 수 있었다. 다만 본 연구가 춘계와 하계라는 두 시기에 현장조사가 이루어졌다는 점에서 현재 건강도 악화가 지속적으로 이루어질 것이라는 결론을 도출하기에는 한계가 있다. 그럼에도 불구하고, 공간적으로 어의도와 서도를 기준으로 서쪽에 위치하고 있는 모든 조사시기의 정점 1, 4, 5, 9, 10, 12, 13, 14, 16, 17, 21 및 23에서 공통적으로 불량한 것으로 나타났다. 특히 이들 정점 가운데 정점 5, 9, 10, 12, 13, 14 및 16은 하계에 0.24-0.60 mg/L의 매우 낮은 용존산소량을 나타내는 특징을 보였다. 결국 본 연구해역은 저층수의 용존산소량이 대형저서동물 군집에게 지대한 영향을 미치고 있음을 생물 평가지수를 이용하여 다시 한 번 잘 입증하고 있다.