Ulva australis Kjellman (formerly called Ulva pertusa) grows on the rocky shores of Korea, and could play an important role as a primary producer in coastal ecosystems as do many seaweeds. The species occasionally forms extensive mats called green tide in coastal areas around the world due to its fast growth and high reproductive capacity (Fletcher 1996, Han et al. 2003, Hiraoka et al. 2004, Kim et al. 2011, Deng et al. 2012). However, there are no available data on whether U. australis plays an ecological role as a bioengineer, providing habitat, nursing grounds, and food sources for various marine animals, to increase the biodiversity in Korea.
Some macroalgal fronds have innate holes, as found in the two brown algae, Agarum clathratum and Costaria costata, some of which are made by biotic activities (endophytes, grazers) or through wave action (De Bettignies et al. 2012, Kim et al. 2014). For example, Undaria pinnatifida are affected by a pinhole disease that resultings from infection by the harpacticoid copepods Amenophia orientalis and Parathalestris infestus, causing the fronds to have many small holes (Park et al. 2008). Holes are also produced by herbivorous grazers in the process of eating (Menge 1978), and wounds associated with grazing and wave action were shown to increase kelp erosion in Ecklonia radiata (De Bettignies et al. 2012). Ulvella operculata (= Acrochaete operculata) is an endophyte that produces degradative lesions in Chondrus crispus fronds, while green endophytes make holes in the fronds of Grateloupia spp. (Correa et al. 1988, Correa and McLachlan 1991, 1992, Kim et al. 2014).
The effects of environmental factors on the growth and eco-physiological features of Ulva spp. have been thoroughly examined (Talylor et al. 2001, Choi et al. 2010, Mantri et al. 2011). In U. australis, physiological responses to exposure to copper and UV light (Han and Han 2005, Han et al. 2008), as well as various environmental factors (i.e., temperature, light intensity, and nutrients, etc.), have been reported (Floreto et al. 1994). In addition, U. australis were found to display antioxidant, allelopathic, and antihyperlipidemic activities (Floreto et al. 1994). Ecologically, ephemeral algae such as Ulva spp. are food resources for gastropod herbivores (Lubchenco 1983, Geertz-Hansen et al. 1993, Giannotti and McGlathery 2001). However, the growth, hole formation, and ecological role of U. australis as a habitat and food resources of marine animals have not yet been determined. Thus, the aims of this study were to examine the growth and hole formation of U. australis, and to investigate the effects of wave action on the growth of the species and on the abundance of residential animals. Finally, whether hole formation in the U. australis fronds is correlated with wave action and residential animals was discussed.
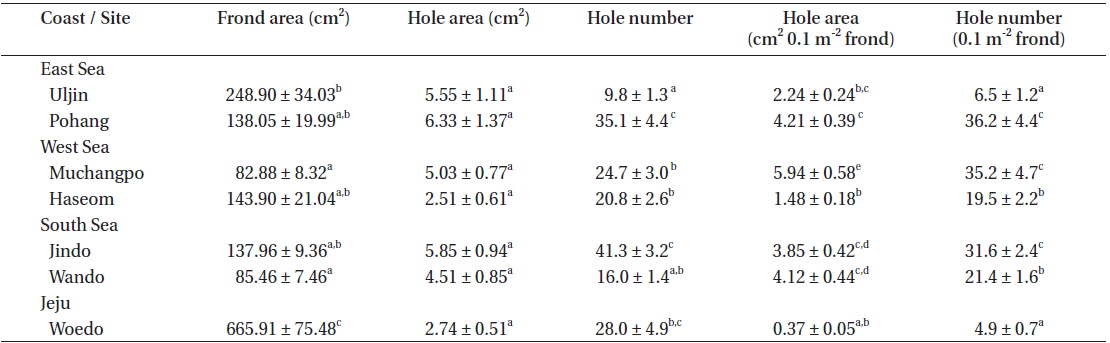
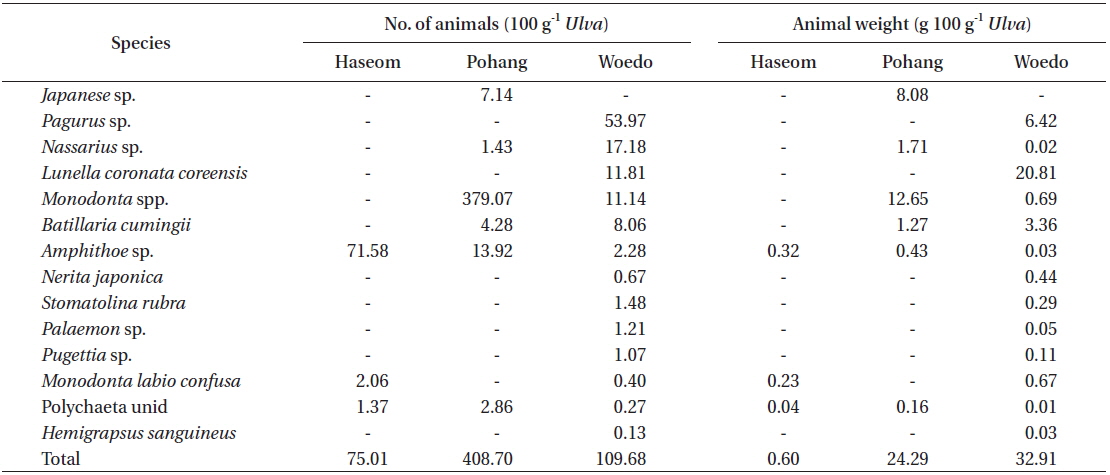
U. australis frond samples were collected at seven study sites located on Jeju Island and along the Eastern, Western, and Southern coasts of the Korean Peninsula between the end of July and early August, 2013 (Fig. 1). At each site, 30 U. australis fronds were randomly sampled. All of the samples were photographed, and Image J software was used to count the number of holes and for measurement of frond and hole area. Hole area (y) and frond area (x) were plotted, and an equation was calculated. The differences in frond area, number of holes, and hole area per 0.1 m2 of Ulva fronds were compared among the seven populations. In addition, more than 100 g wet weight of Ulva fronds were collected from Haseom, Pohang, and Woedo in order to examine the abundance and species composition of the residential animals on the Ulva fronds. The animals living on the Ulva fronds were sorted, identified, counted, and weighed for each population, and then the number and weight of the animals per 100 g of Ulva fronds was determined.
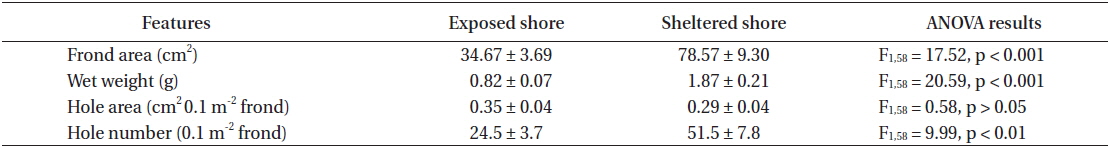
To examine the effects of wave action on the growth and hole formation of U. australis fronds, as well as on the abundance of residential animals, samples were also collected from wave-exposed and sheltered sites of Seongsan, Jeju, Korea on October 19, 2013 (Fig. 1). The levels of wave exposure were measured using a dynamometer, which was made following the protocol of Bell and Denny (1994). Relative levels of wave exposure, from zero (no movement) to 1 (full tie length), were calculated using the distance moved by a rubber indicator connected to a practice golf ball with a nylon cable tie. The wave-exposed site of Seongsan displayed relatively higher wave action (0.7 of 1.0) than the sheltered site (0.2).
Statistical analyses were performed using STATISTICA version 5.0 (StatSoft Inc., Tulsa, OK, USA). One-way ANOVAs were performed separately in order to test for differences in the morphological features of the seven populations, including the frond area, hole area, and hole number per 0.1 m2 of Ulva frond. One-way ANOVA was used to test the effects of wave action on the growth and hole formation of Ulva fronds for the Seongsan populations. Prior to analysis, the homogeneity in the variance was tested using Cochran’s test. Tukey’s multiple comparison was employed, when significant differences between means were detected (Sokal and Rohlf 1995).
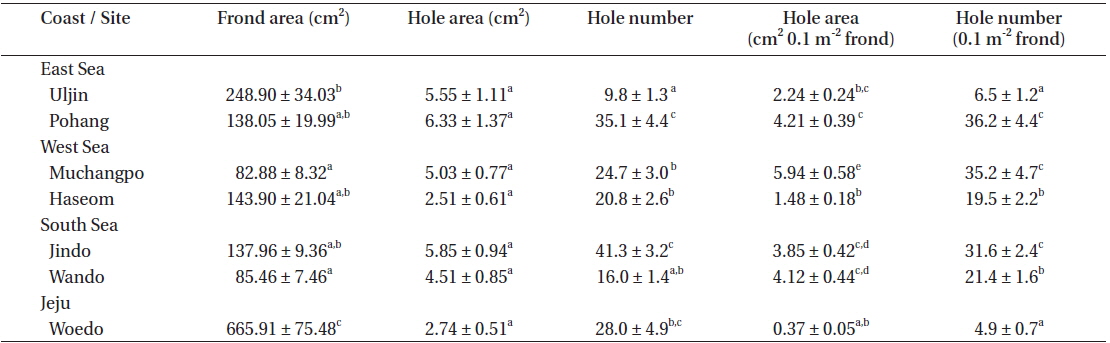
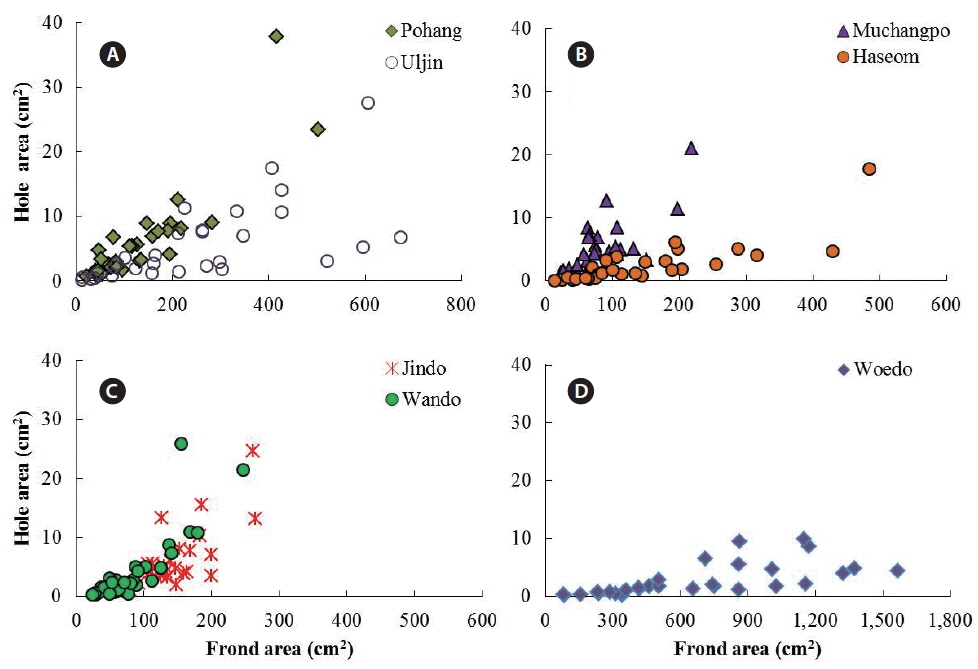
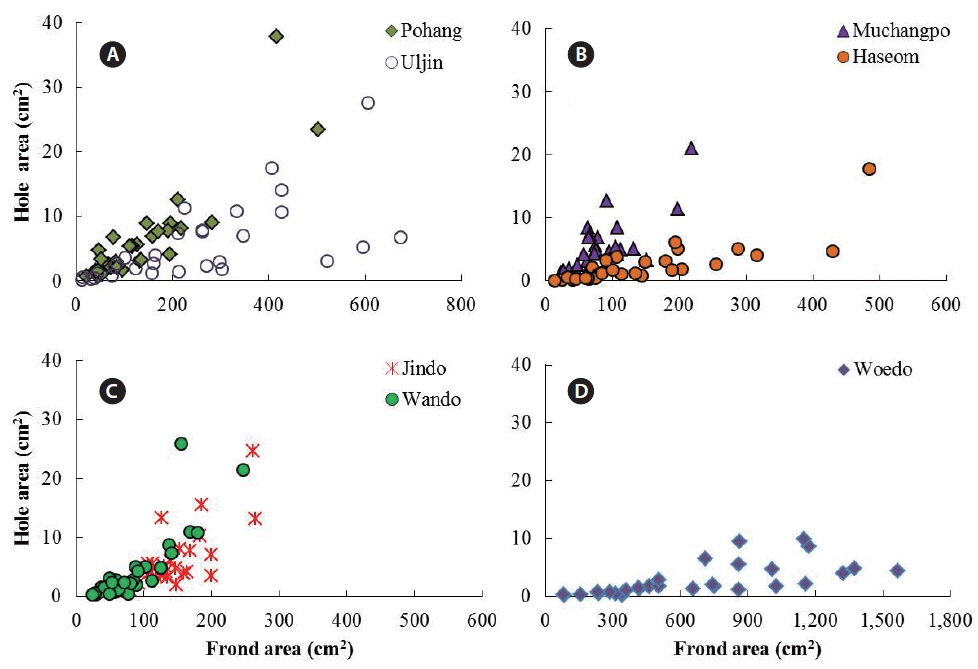
The average frond area of the U. australis population (n = 30 fronds) ranged from 82.88 cm2 at Muchangpo to 665.91 cm2 at Woedo, Jeju (Table 1). The differences in frond area were statistically significant (F6,203 = 37.52, p < 0.001), with remarkably larger values observed in the Woedo population (Table 1). The hole area ranged from 2.51 to 6.33 cm2 per frond, demonstrating the largest values in the Pohang population and smallest in the Haseom population. However, significant differences were not observed in terms of the hole area (F6,203 = 2.44, p > 0.05). An average of 9.8 to 41.3 holes were observed per frond, with the greatest among the seven study sites observed for the Jindo population (Table 1). Hole areas (y) were positively correlated with the frond areas (x) of U. australis (Fig. 2) for the samples taken from Pohang (y = 0.019x1.14, r2 = 0.77), Uljin (y = 0.023x0.95, r2 = 0.70), Muchangpo (y = 0.037x1.08, r2 = 0.53), Haseom (y = 0.002x1.37, r2 = 0.79), Wando (y = 0.001x1.70, r2 = 0.75), Jindo (y = 0.001x1.80, r2 = 0.67), and Woedo (y = 0.001x1.19, r2 = 0.68).
Examination of a fixed area of the frond revealed hole area (cm2 0.1 m−2 frond) to be between 0.37 and 5.94 cm2, with hole numbers (0.1 m−2 frond) in the range of 4.9 to 36.2 (Table 1). The smallest area and number of holes were observed for the Woedo population, even though the frond area was the largest of the seven Ulva populations examined. Hole area (cm2 0.1 m−2 frond) was the largest for the Muchangpo population, while the hole number was greatest in the Pohang population (Table 1).
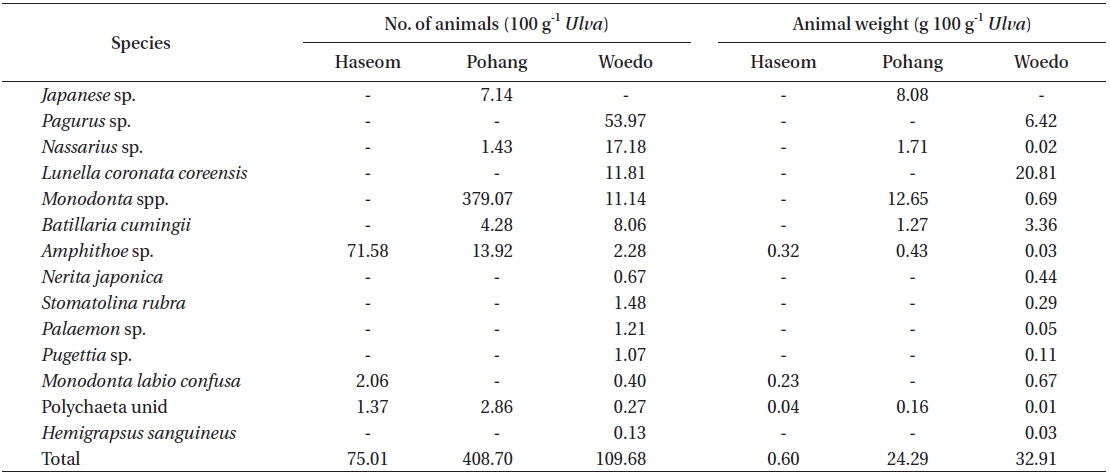
Fourteen species of marine animals were found in the Ulva fronds, with 3 to 13 species found at each location (Table 2). The species richness was the greatest at Woedo and the lowest at Haseom. The number of animals (100 g−1 frond) varied from 75.0 at Haseom to 408.7 at Pohang. In terms of the number of animals, the dominant species differed according to the region, with gammaridea (Amphithoe sp., 95.43%) identified for Haseom, snails (Monodonta spp., 92.75%) for Pohang, and crab (Pagurus sp., 49.20%) for Woedo. The average animal weight (100 g−1 frond) was minimal, making up 0.60 g wet weight at Haseom and 32.91 g at Woedo. The dominant animals in terms of the average weight were also the same, with gammaridea (Amphithoe sp., 54.06%) for Haseom and snails (Monodonta spp., 52.09%) for Pohang. However, Lunella coronate coreensis was dominant in terms of weight at Woedo.
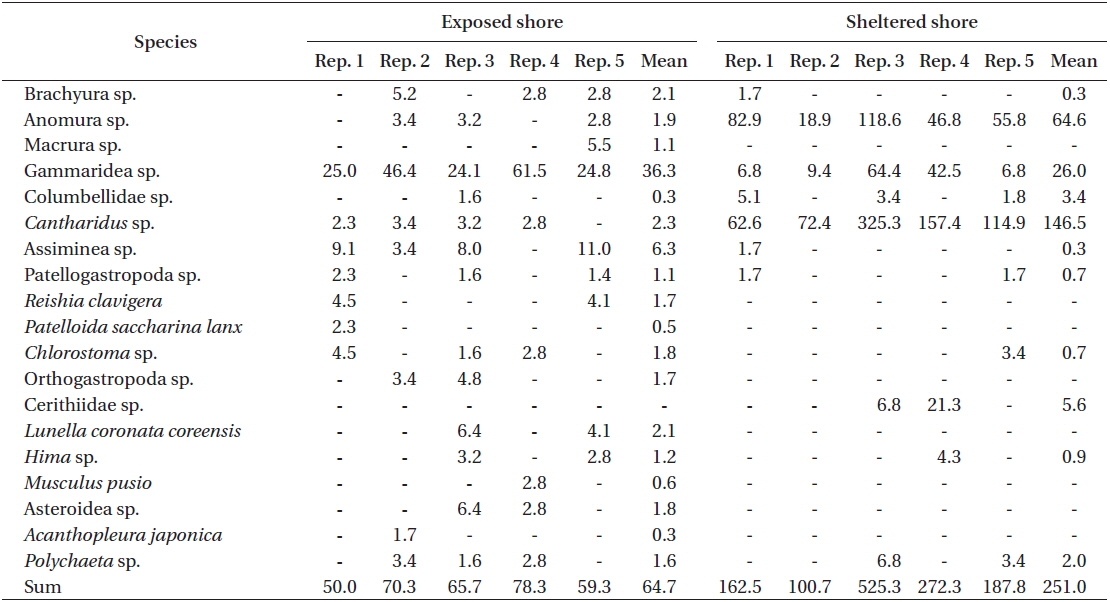
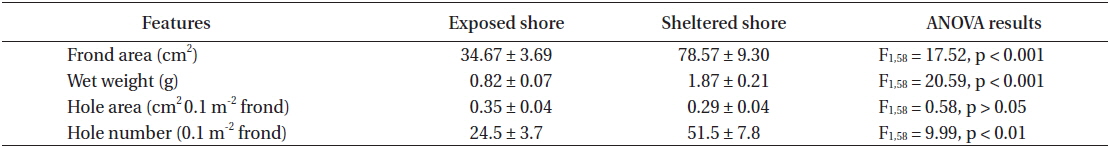
The frond area and weight of U. australis at the sheltered station of Seongsan were twice those at the exposed shores, with significantly difference between the two levels of wave exposure (Table 3). While no significant difference was observed in hole area (0.1 m2 frond), the number of holes was significantly greater in the sample from the sheltered shore than from the wave-exposed shore (Table 3).
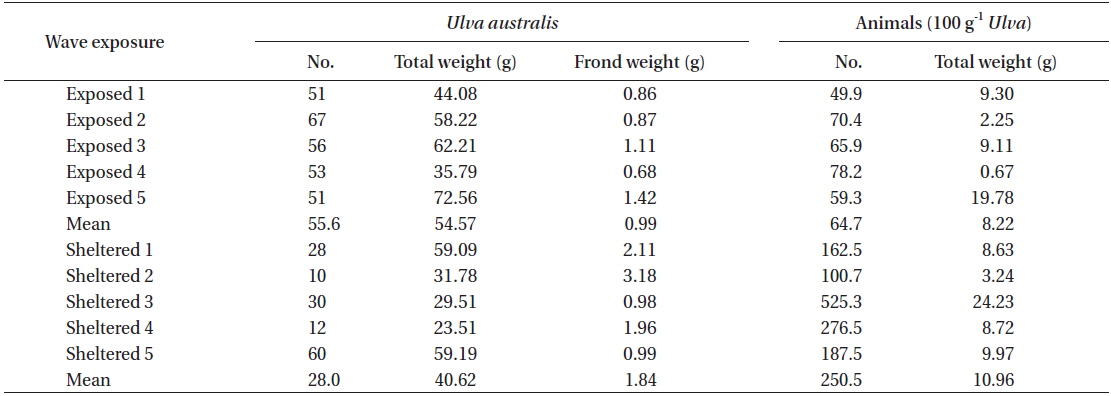
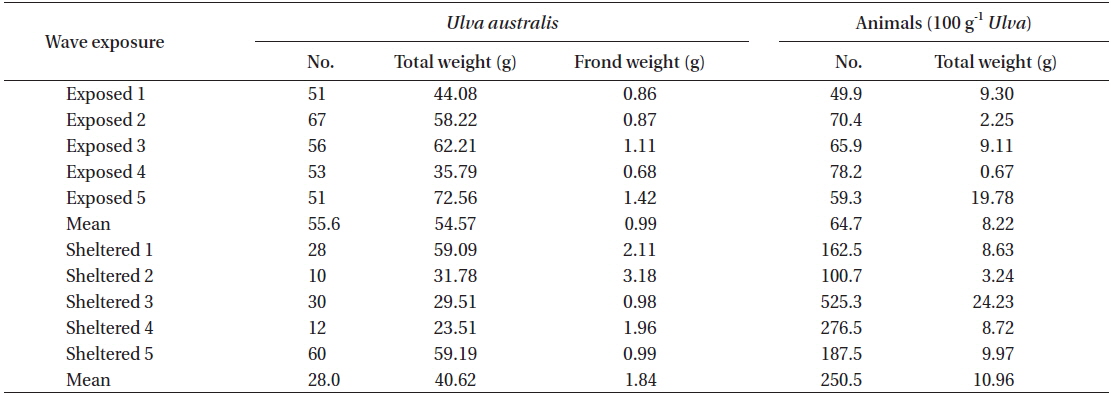
In the U. australis populations of Seongsan, the average frond weight at the sheltered shores was twice as large as that at the exposed shores, measured at 1.84 ± 0.41 ver. 0.99 ± 0.13 g wet weight (mean ± SE, n = 5 replicates) (Table 4). However, no significant differences were observed between those exposed to waves in terms of the individual weight of the Ulva fronds (F1,8 = 3.99, p > 0.05). At the sheltered shore, the number of animals (100 g−1 Ulva) was four times greater (F1,8 = 6.23, p < 0.05) than at the waveexposed shore, while the animal weight was only slightly greater (F1,8 = 0.31, p > 0.05) (Table 4). That is, the animals inhabiting Ulva fronds were abundant in both number and weight at the sheltered shores of Seongsan.
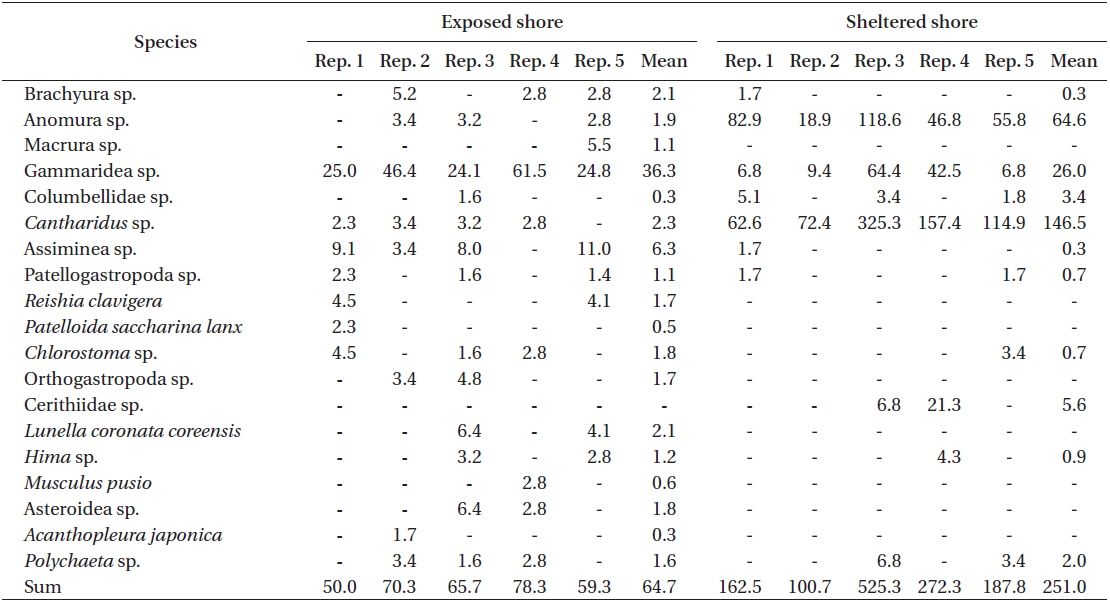
A total of 19 species or taxon was found in the Ulva population of Seongsan, with 18 species observed at the exposed shore and 11 species at the sheltered location. With respect to the Ulva fronds, an abundance of crabs, gammaridea, and snails was observed, irrespective of exposure to waves (Table 5). At the exposed shores, the dominant species were Gammaridea sp. with subdominant species of Cantharidus and Assiminea. However, Anomura sp. was dominant, with Cantharidus sp. as the subdominant species at the sheltered shores.
The growth and hole formation of Ulva australis displayed significant difference among the populations along the coast of Korea. Frond area was eight times greater at Woedo than in the Muchangpo and Wando populations. Ulva spp. is a notorious species that produces green macroalgal blooms in eutropicated or disturbed areas caused by anthropogenic activities on the coast of Korea and around the world (Taylor et al. 2001, Han et al. 2003, Kim et al. 2011). Generally, green tides of Ulva spp. tend to occur at sites with soft bottom, including mud, silt or sand, with floating Ulva species (Hiraoka et al. 2004, Kim et al. 2011). So far, there have been no reports of green tide due to U. australis growth on rocky shores, but this species is becoming dominant in the coastal areas of Korea (Choi et al. 2010). In addition, it is known to grow well in wide range of salinities as do many green tide algae such as Ulva fasciata, U. rigida, and U. lactuca (Zavodnik 1975, Morand and Briand 1996, Giannotti and McGlathery 2001). Thus, it is very important to examine the growth of U. australis in coastal areas of Korea, and to elucidate the environmental factors regulating the growth of the species in the field population. The growth of Ulva spp. is determined by abiotic factors, including temperature, light intensity and nutrients, as well as by top-down control through marine invertebrate grazing (Floreto et al. 1994, Giannotti and McGlathery 2001, Talylor et al. 2001, Choi et al. 2010, Mantri et al. 2011). The present results demonstrated that the growth of U. australis was controlled by wave activity and herbivorous animals in the field population.
In the present study, 14 species of marine animal were observed in three U. australis populations examined, while 19 species were observed in the populations from Seongsan, Jeju, indicating that this species is a bioengineer that increases the biodiversity of various marine animals by providing a habitat, nursery grounds, and food sources, like seaweeds do (Terawaki et al. 2001, Eklöf et al. 2005). The dominant animal species growing on the Ulva populations differed according to the study sites, but gammaridea (Amphithoe sp.), gastropod (Monodonta spp.), and crab (Pagurus sp.) were most commonly observed. The abundance of grazers was positively correlated with the hole number in the Ulva fronds for the Pohang and the Seongsan populations. These results indicate that the Ulva population provides habitats and food resources for various marine invertebrated grazers, which make holes in the fronds. In addition, decayed Ulva fronds can become foodstuff for filter feeders or detritus feeders as a component of particulate and dissolved organic matter, as reported in studies of Ulva lactuca (Giannotti and McGlathery 2001) and kelp (Mann 2000).
Wave activity was also found to influence the growth and hole number of Ulva fronds in the Seongsan populations. U. australis grew better and had more holes at the sheltered shore, where more residential invertebrates were present than at exposed shore. The holes in macroalgal fronds can be produced either genetically, or as a result of biotic or abiotic activities, including those of endophytes, grazers, or wave activity, as observed for C. crispus, U. pinnatifida, and E. radiata (Correa et al. 1988, Park et al. 2008, De Bettignies et al. 2012, Kim et al. 2014). The present results indicate that the holes in the U. australis fronds are mainly related to grazing pressure.
Marine invertebrate grazers, such as gastropods, amphipods, and isopods reduce macroalgal biomass by making holes in the fronds (Giannotti and McGlathery 2001, Martins et al. 2014). In the present study, the dominant animal species in the Ulva populations were gammaridea, gastropods, and crab, which usually make holes or tear the fronds while feeding. Park et al. (2008) reported that small holes of U. pinnatifida fronds are made by the feeding activities of copepods, A. orientalis and P. infestus. Hauxwell et al. (1998) reported that amphipod and isopod grazers consumed significant amounts of macroalgae in the field. Grazing is a form of biological stress that can damage Ulva tissue by producing holes, which may result in loss of biomass due to secondary physical factors, including wave action, pathogens, and abrading sediments. However, a laboratory feeding experiment was not performed on the three dominant residential animals observed herein. Thus, to make the conclusion that residential animals caused the frond holes on the U. australis, more detailed experiments should be conducted in the future. In simulated wound experiments, the biomechanical properties of E. radiata fronds exposed to strong wave activity during the winter exhibited a significant reduction as a result of holes (or cuts), increased kelp pruning rates, and substantial reduction in biomass (De Bettignies et al. 2012).
In conclusion, U. australis populations play an important ecological role as a habitat and as foodstuff for many marine invertebrates, and their population structures differ according the location in terms of frond area, hole number and hole area. In addition, growth and hole number of Ulva fronds around the coast of Korea are affected by wave activity, as well as by the number of herbivorous animals, such as isopods and gastropods.