Although rotifers have been considered the best feeding option for several species of fishes in aquaculture, they are sometimes larger than appropriate for the early larval stage of some marine fishes. Thus, we aimed to determine whether size-based selection of the parents could affect the average body size of their progeny in two clonal populations of the rotifer Brachionus rotundiformis. From each of the clones, 20 individuals were bi-directionally selected toward both smaller and larger sizes and each individualbased selection was conducted for 10 consecutive generations. The results showed that although there were sometimes differences in mean body size between parents and their progeny, no directional trend was observed in all selected lines of both clones. We demonstrated that artificial selection in a rotifer stock cannot lead to an expected size range although they appear to exhibit a large degree of body size polymorphism.
Size-based selection of food in fish (Ghan and Sprules, 1993; Deudero and Morales-Nin, 2001; Shaw et al., 2003) and crustaceans (Harvey and Epifanio, 1997) has been demonstrated in both natural and cultural environments. Shirdhankar and Thomas (2003) reported that the food digestibility of larval or juvenile fish was determined to a great extent by the size of food particles in relation to the mouth size of the predator, and Shaw et al. (2003) indicated that the relationship between prey size and mouth size was the primary determinant of prey selection. With the worldwide development of aquaculture industries, a main challenge for aquaculturists has been to provide live food with suitable characteristics, including the proper size, during the larval stage of fish development. This has been a critical challenge with some groups of fish. For instance, a high mortality rate in grouper larvae was related to their sensitivity to prey size (Kohno et al., 1997).
Rotifers are used as live food in the larval rearing of more than 78 species of marine finfish and crustaceans. The demand for rotifers is still increasing (Fu et al., 1997). However, even small strains of rotifers have sometimes been found to be too large for smaller-mouthed larvae of aquaculture candidates such as grouper and rabbit fish (Rodriguez and Hirayama, 1997). In order to supply cultured rotifer as a cost-effective feed for fish larvae, it is important to provide the appropriate size of rotifers to the larvae (Hagiwara et al., 2001). A preliminary approach for this is the selection of rotifer strains with suitable size ranges (Kotani and Hagiwara, 2003). Several investigators have studied the effect of various environmental conditions on rotifer body size. Fukusho and Iwamoto (1981) examined the influence of various feeds on the size and shape of L-type rotifers and reported that body size increased when rotifers were fed with ω-yeast or a combination of baker’s yeast and formula feed for prawns. In contrast, Yufera (1982) concluded that rotifer body size was primarily determined by genetics and not greatly influenced by environmental conditions, e.g., dietary manipulation. Snell and Carrillo (1984) conducted a more intensive investigation of body size variability to determine the effects of salinity, temperature, and rotifer strain on lorica length. They concluded that while lorica size was largely determined by the genetics, small modifications of lorica size were possible by environmental manipulation, although an independent effect of either temperature or salinity was not statistically significant. Glavic et al. (2000) proposed that it was possible to produce rotifers with required lorica size by changing environmental conditions such as temperature and salinity.
Although rotifers have been used as live food in aquaculture for nearly 50 years, there is an apparent lack of information on its quantitative genetic aspects. Selective breeding, as used in farm animals and plants, is a time-consuming genetic manipulation technique that can play a major role in the development of lines having required traits. However, quantitative analyses such as monitoring the selection pressure and tracing the genetic pathways and heritability in rotifers seem to be limited by life history characteristics such as microscopic size, short life span, parthenogenetic reproduction, short generation length, and short time to maturity. Different types and strains of the genus
The objective of this study was to evaluate the response of
>
Source and maintenance of rotifers
Two colonial populations of a laboratory stock of the rotifer
>
Measurement of body size and estimating agedependent size variation
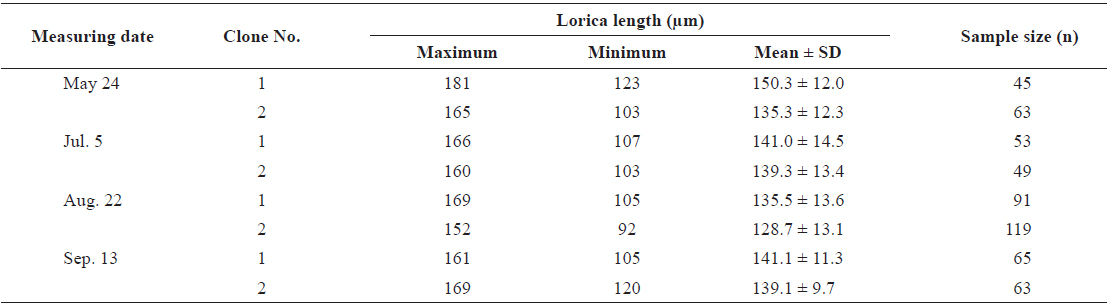
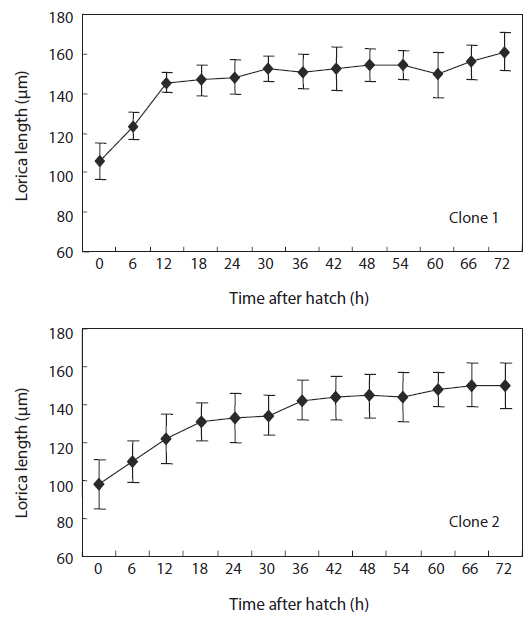
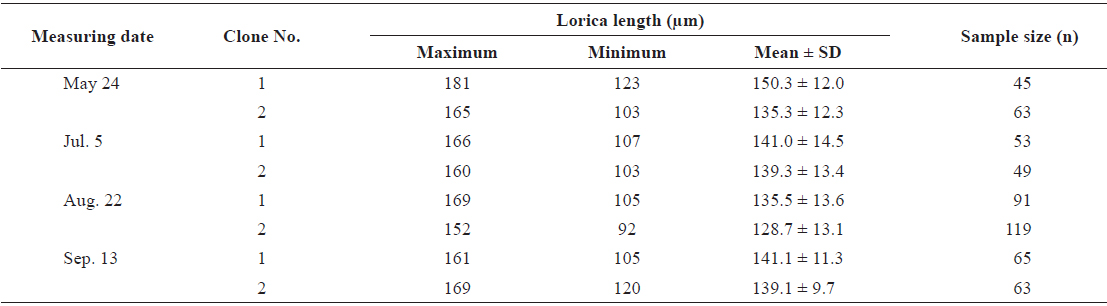
From cultures of each clone containing 400 rotifers/mL, a volume of 20 mL was randomly sampled and fixed with 5% formalin to allow the measurement of mean lorica length of each population using a stereo-microscope at 100X magnification. The sizes of two experimental clones at different measuring times are shown in Table 1. To estimate size increments during early life, 30-40 rotifers were sampled and fixed with 5% formalin at 6-h intervals until the age of 72 h after hatching.
According to the size range of the rotifers (Table 1), and in a preliminary attempt to select rotifers with small lorica length, plankton nets of different mesh sizes (80, 100 and 124 μm) were used to select for neonate rotifers of similar size and of the same generation. Several trials of batch filtration failed to collect rotifers at the expected size ranges. This was due to the characteristic inequality of the length and width of the rotifer body and its unpredictable orientation while passing through the mesh, the soft and flexible body texture, the presence of attached eggs, and difference in body size even at the same age. An additional disadvantage of this method is that, because of the short generation time to maturity, several generations of rotifer were present at any given time, and prevented the isolation of individuals of the same age, tracing of descendancy, and eventual calculation of genetic factors such as heritability.
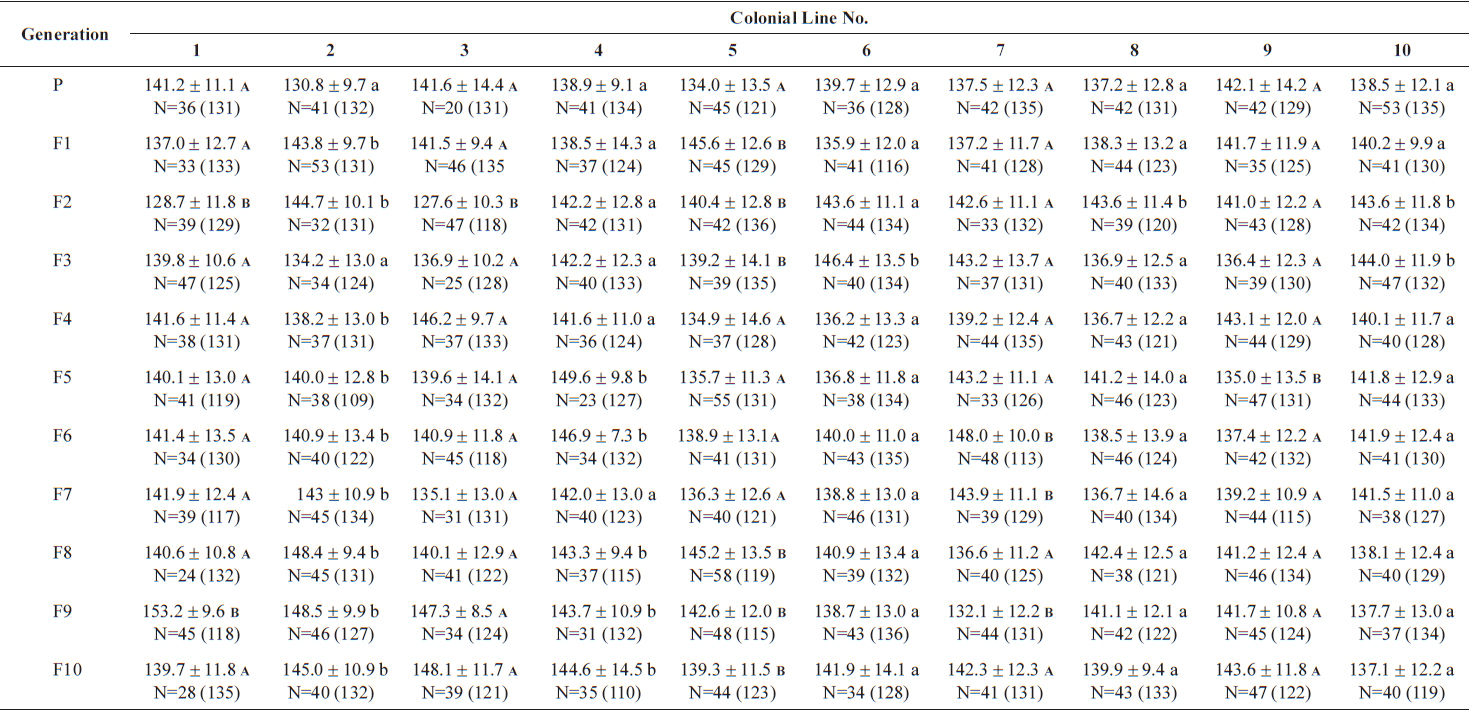
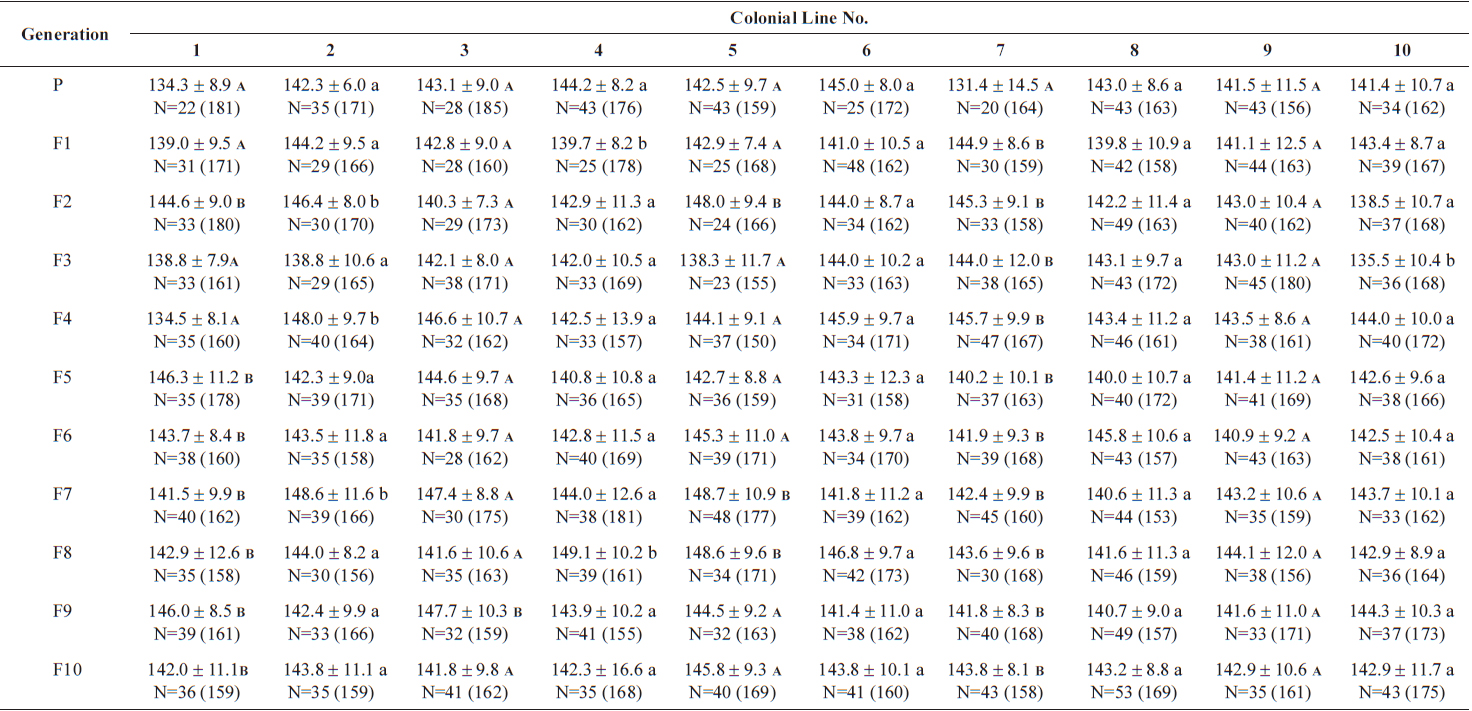
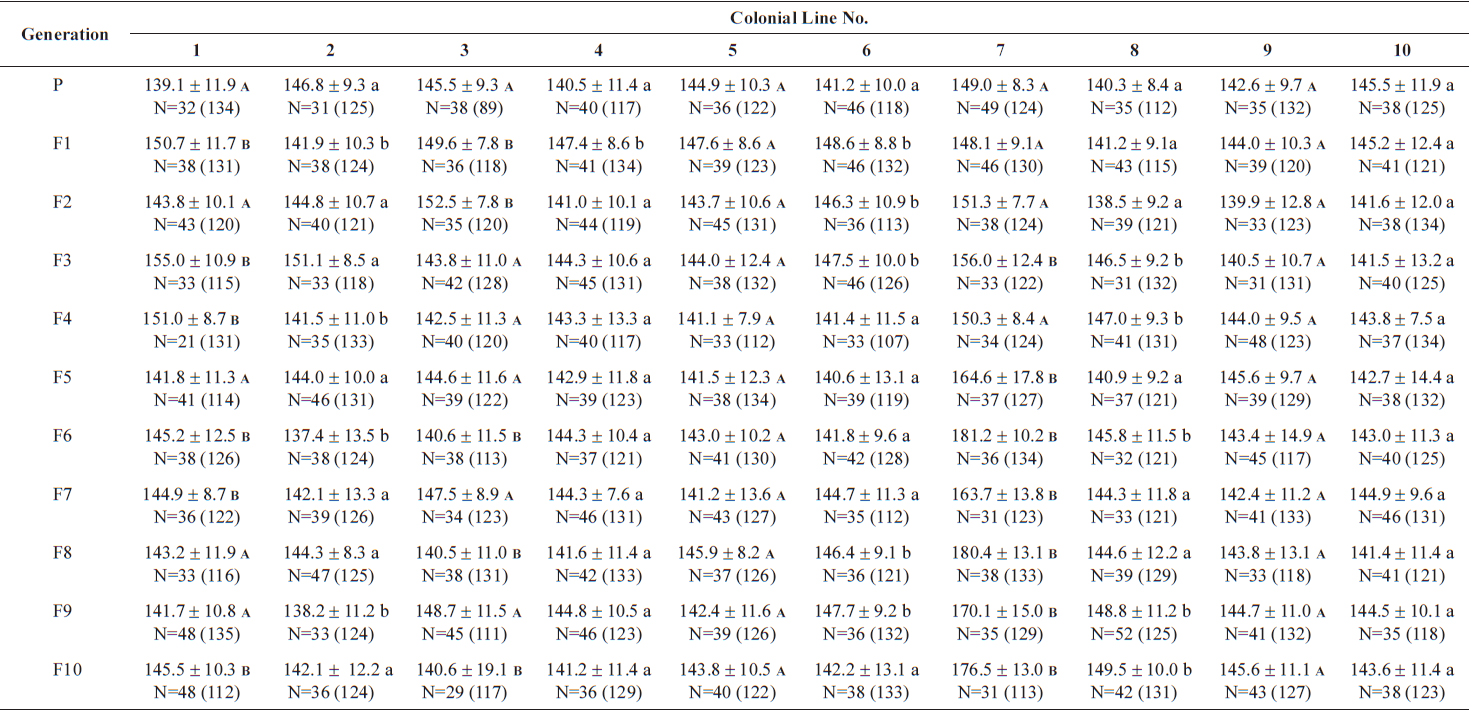
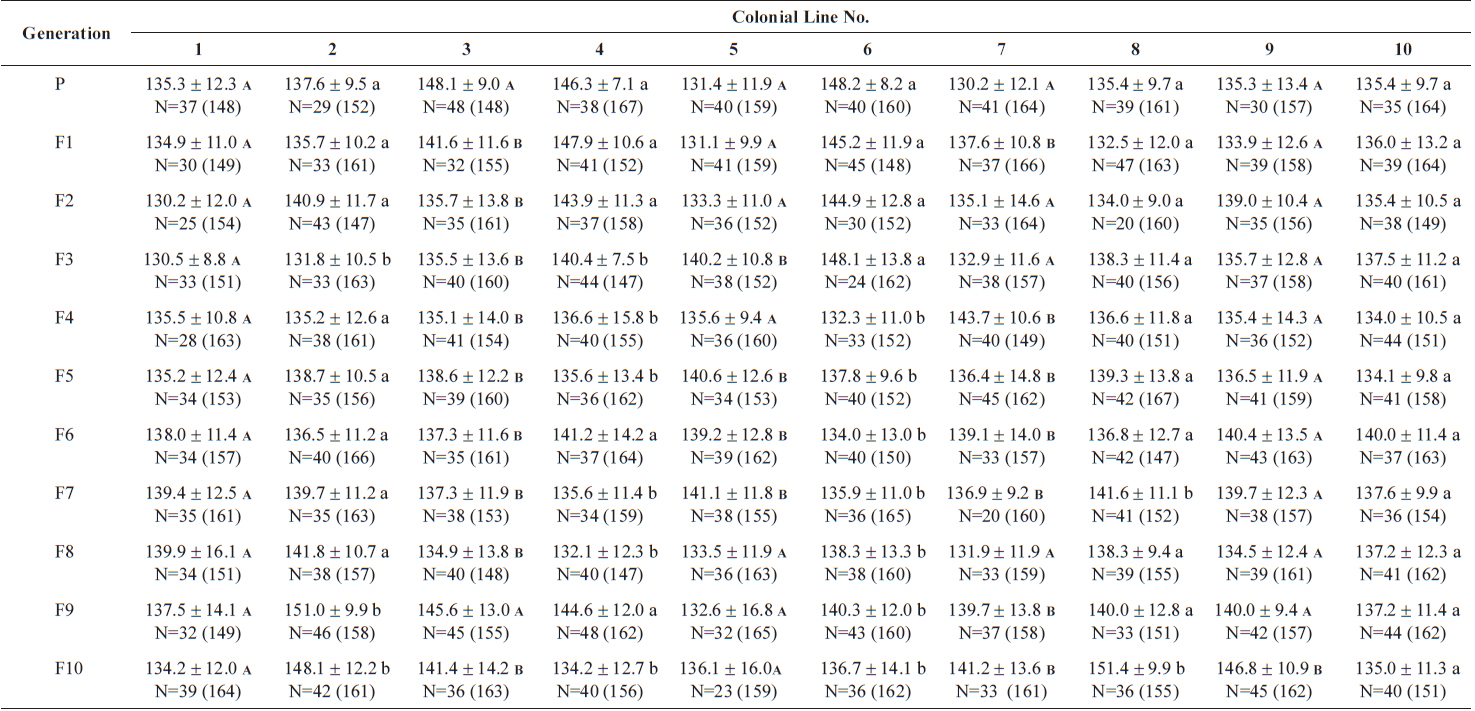
Size values of two rotifer Brachionus rotundiformis clones chosen for size-based selection at different times
An alternative method of individual selection was adopted to minimize the aforementioned disadvantages. Isolated individuals of an appropriate size range were used to examine selection effects in successive generations. A brief description of this method is as follows: When good cultural conditions were maintained, many egg-bearing female rotifers were observed under the microscope at 40X magnification. Parts of each colonial culture of rotifer were transferred to a Petri dish for microscopic viewing. With careful observation, the smallest and largest females carrying eggs were removed by pipetting them from the culture medium and transferring them individually to single wells of a 24-well tissue culture plate. Each well was supplied with 2 mL of autoclaved seawater and the culture conditions and feeding rate were the same as that used in the initial culture. For each selective direction (small and large body size) of each clone, 10 parental individuals were monitored for 10 generations. The first parental females (P) selected for both small and large size were checked for progeny (F1), and the second selection was conducted in the same manner with F1 individuals. After the F1 selection, the parental individuals were fixed in formalin and their sizes were measured. The remaining progeny of each parent were left to grow and make their own population. When a large enough number of rotifers were grown in each generation, an aliquot was fixed for size measurement.
The collected data were analyzed with the SPSS software (Ver 14.0 SPSS Inc.). Both analysis of variance (ANOVA) and Student’s t-tests were performed to examine the bi-directional size-based selection effect on the rotifers. As the latter test turned out to be more robust than the former, most of the statistical analyses consisted of Student’s t-tests. Simple correlation analyses were performed to understand the relationship between some morphological traits by calculating Pearson correlation coefficient (r). Intra- and inter-colonial size variations were compared using two statistical factors such as the coefficient of variance (CV) and ANOVA.
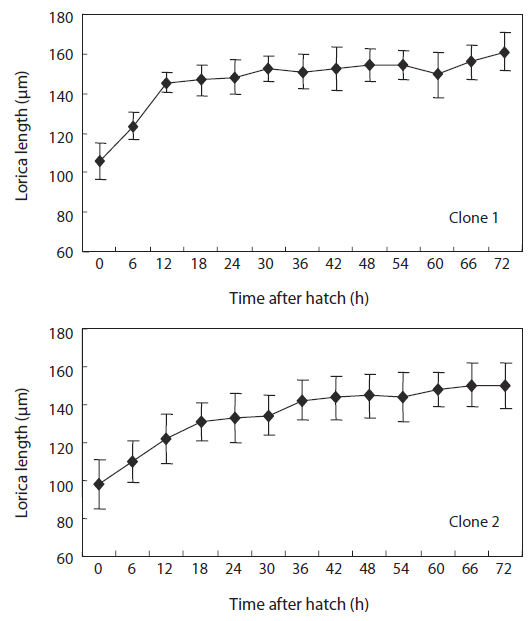
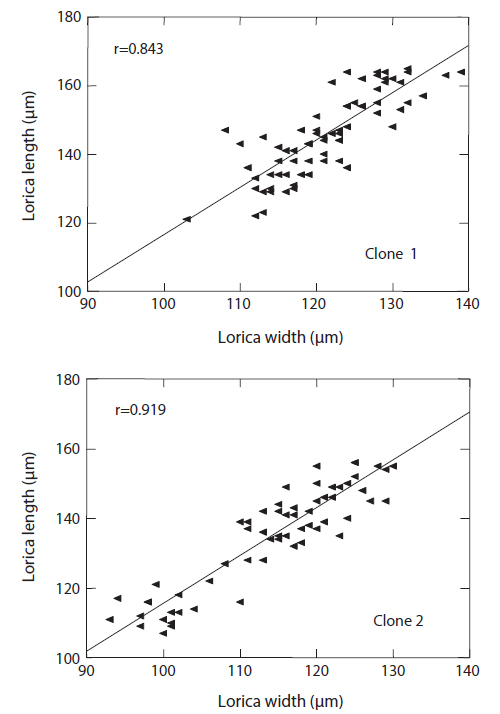
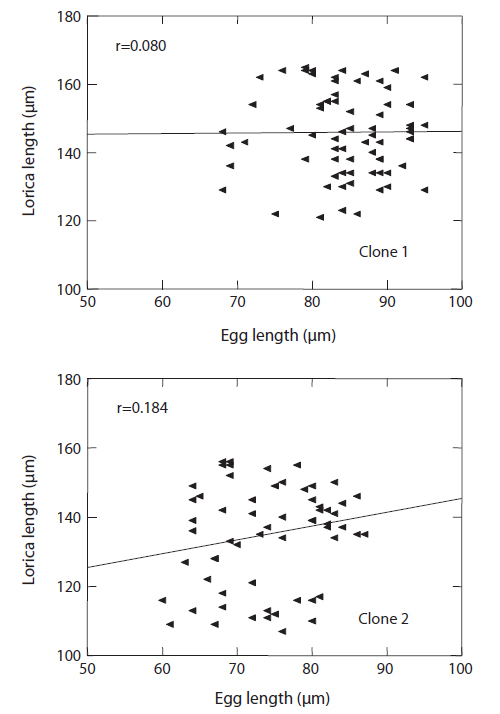
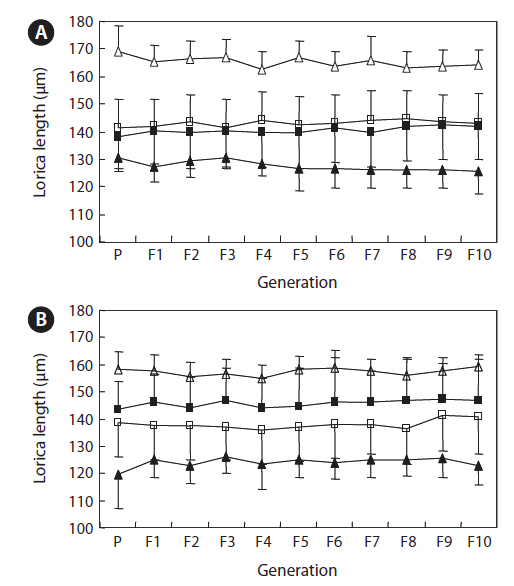
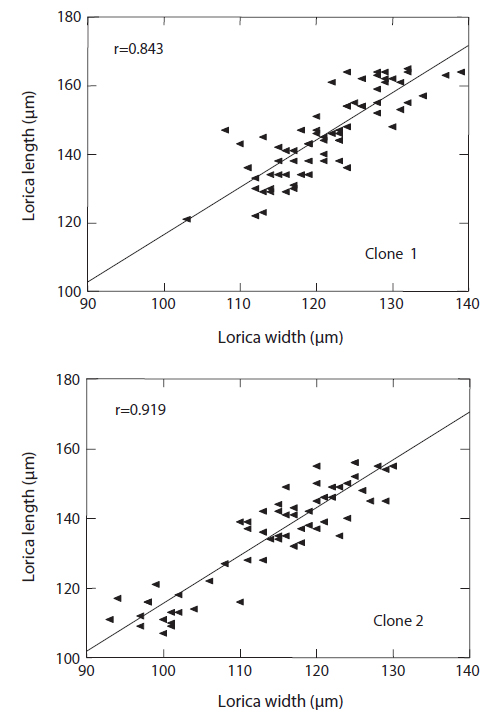
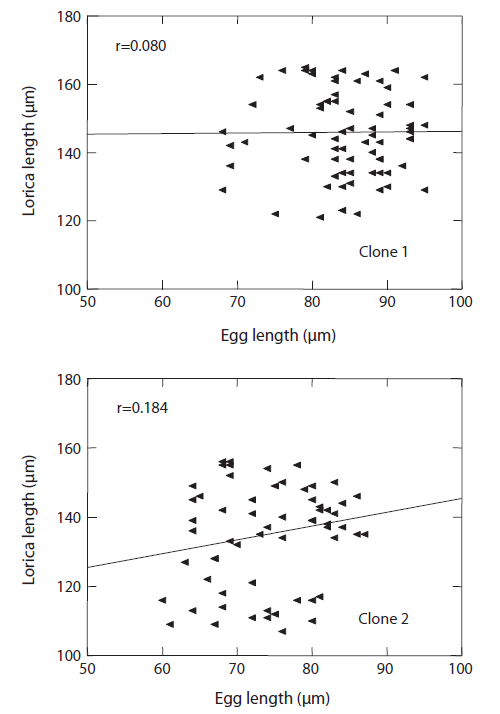
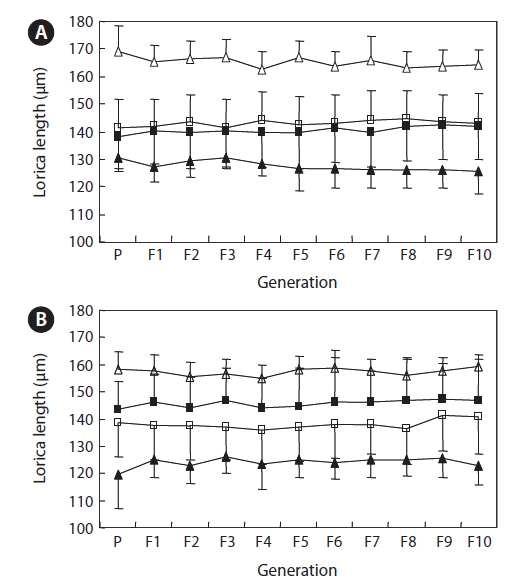
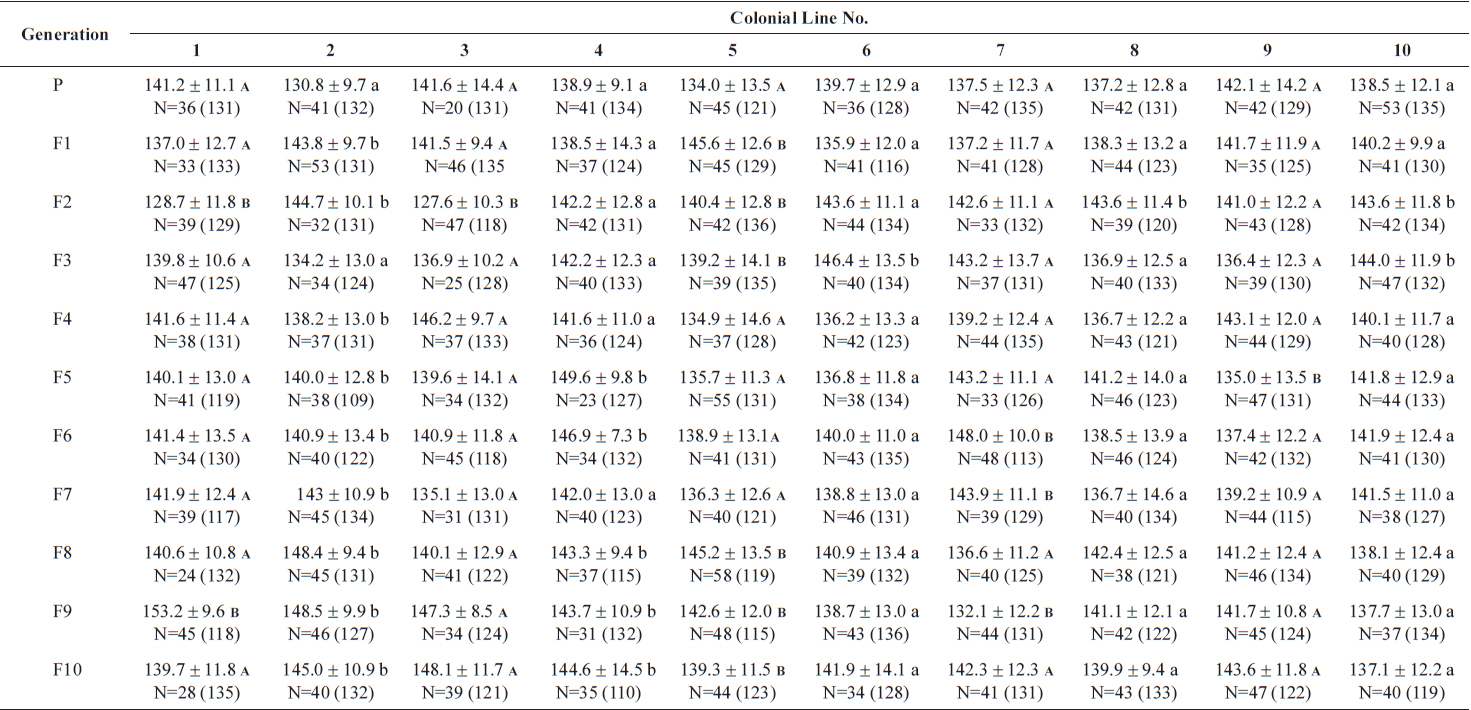
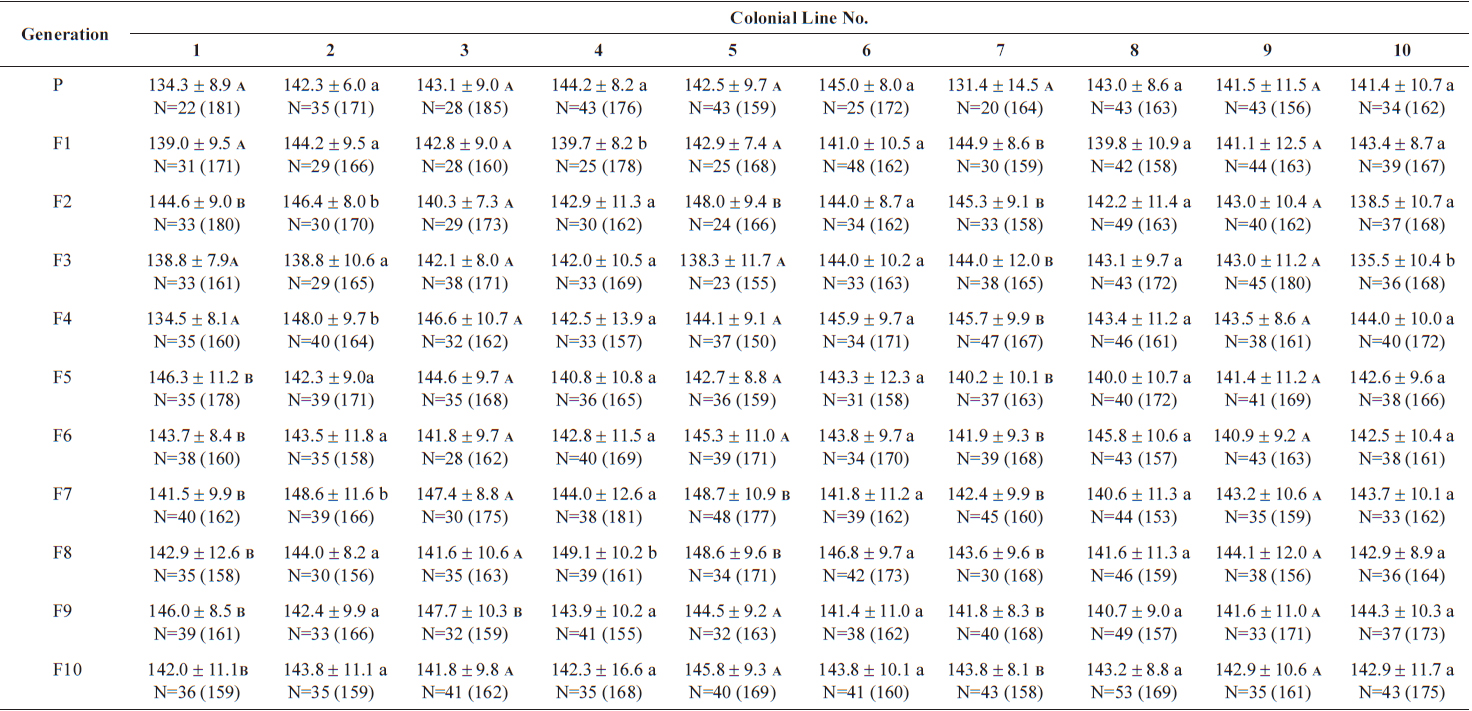
The changes in body size of the two clones were examined until 72 h post-hatching, and the results are shown in Fig. 1. Growth curves indicate that the rotifers grew faster during the first 12 to 18 h after hatching than any other point in their life span, and that their body size increased even after adulthood. The first eggs are usually produced 12–18 h post hatching and hatched rotifers reach their maximum size after about three days. Changes in lorica length was correlated with body width in both clones (Fig. 2), so that body width increased linearly with length (r = 0.843 and 0.919 for clones 1 and 2, respectively). Conversely, lorica length of the parental rotifers was not likely correlated with egg length because the correlation coefficient for these two properties were r = 0.080 and 0.184 for clones 1 and 2, respectively (Fig. 3). In order to determine the effect of size-based selection, mean body sizes and standard deviations (SD) were estimated for 10 consecutive generations. Each generation selected for either small or large body size produced populations using two clones. The results are presented in Tables 2 to 5. The sizes of the smallest rotifers chosen as the first parental individuals were 121–135 μm for clone 1 and 89–134 μm for clone 2, while the sizes of the parents selected for large size ranged from 156–185 μm and 148–167 μm for clones 1 and 2, respectively. In clone 1, the mean lorica length of the small-sized parental population ranged from 130.8 ± 9.7 μm to 142.1 ± 14.2 μm, while those at the 10th generation were between 137.1 ± 12.2 μm and 148.1 ± 11.7 μm (Table 2). The large-sized parental population from clone 1 ranged from 131.4 ± 14.5 μm to 145.0 ± 8.0 μm, while those of the 10th generation were between 141.8 ± 9.8 μm and 145.8 ± 9.3 μm (Table 3). In clone 2, the mean lorica length of the small-sized parental populations ranged from 139.1 ± 11.9 μm to 149.0 ± 8.3 μm, while those at the 10th generation were between 140.6 ± 19.1 μm and 176.5 ± 13.0 μm (Table 4). However, the largest mean lorica length (176.5 ± 13.0 μm) of the 10th generation seemed to be unusual since it was observed only in some generations of the selected colonial line No. 7. The mean lorica length of the large-sized parental population of clone 2 ranged from 130.2 ± 12.1 μm to 148.2 ± 8.2 μm, while those at the 10th generation were between 134.2 ± 12.0 μm to 151.4 ± 9.9 μm (Table 5). In Tables 2 through 5, the mean body size of each generation having the same designation was not statistically different from that of the first parental population (
The lorica length (mean ± standard deviation) of small-sized parental populations (clone 1) and their descendants
The lorica length (mean ± standard deviation) of large-sized parental populations (clone 1) and their descendants
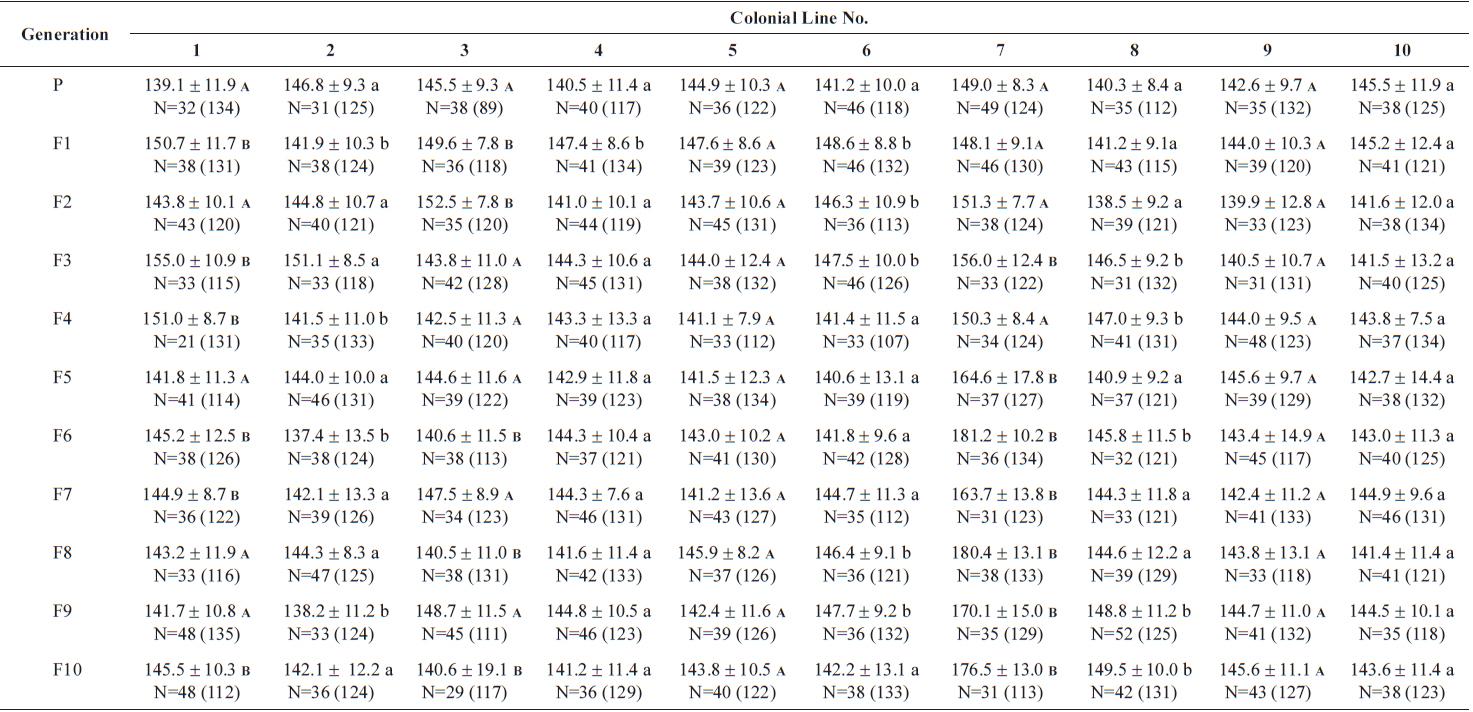
The lorica length (mean ± standard deviation) of small-sized parental populations (clone 2) and their descendants
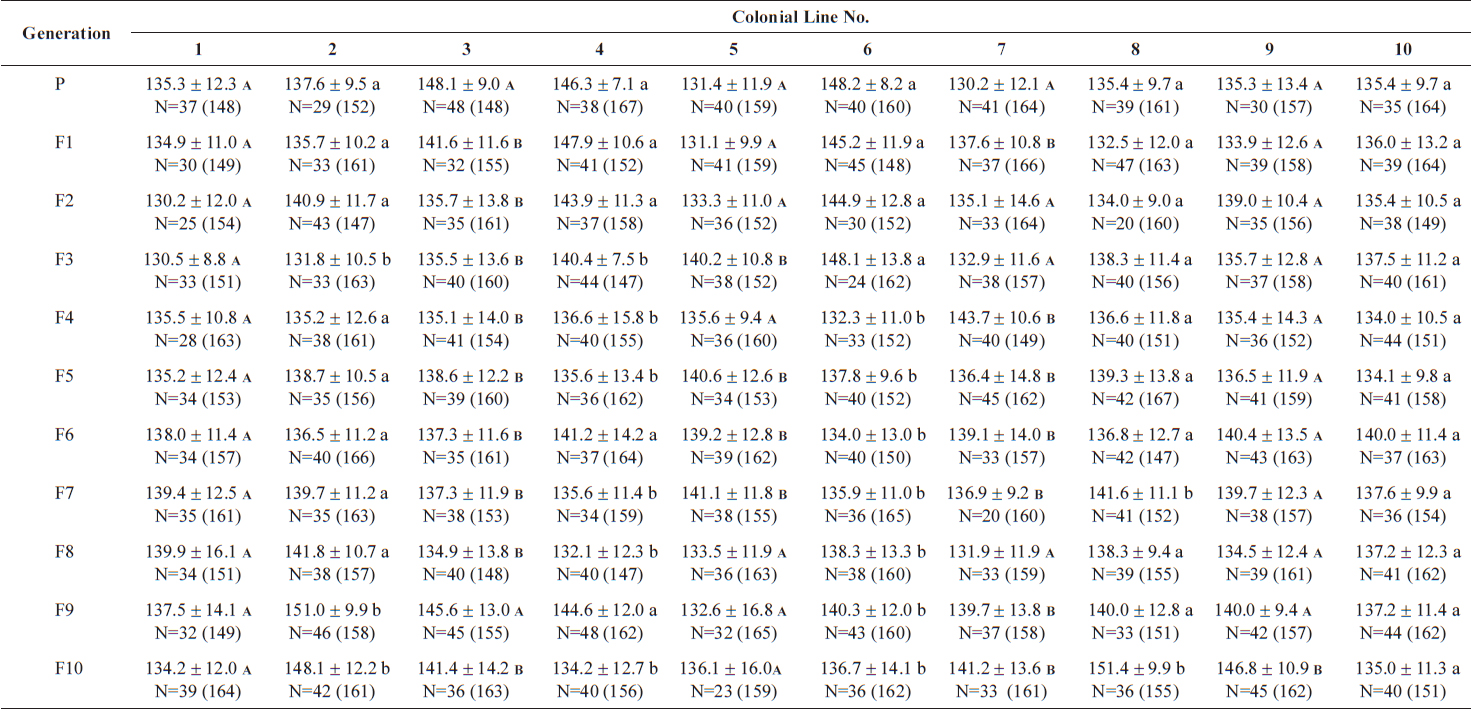
The lorica length (mean ± standard deviation) of large-sized parental populations (clone 2) and their descendants
The body size of rotifers is considered a critical characteristic and determines the adequacy of rotifers as food for young larvae (Rumengan et al., 1998). Rotifer body size was found to be primarily determined by genetics and the influence of environmental conditions was negligible (Yufera, 1982; Snell and Carrillo, 1984). Song et al. (1999) found that the lorica length of the rotifer
In conclusion, the results of this experiment show that: (1) the rotifers of the same age were a variety of sizes in a clonal population, (2) there is no correlation between rotifer body size and the size of its parthenogenetic eggs (see Fig. 3), and (3) no directional trend via size-based selection was observed in all of the selected lines of both clones, although there were sometimes differences in mean body size between parents and their progeny. Overall, we demonstrated that artificial selection cannot lead to an expected size range in a rotifer stock with an identical genetic background due to parthenogenetic reproduction, and that such population exhibit a large polymorphism in body size.