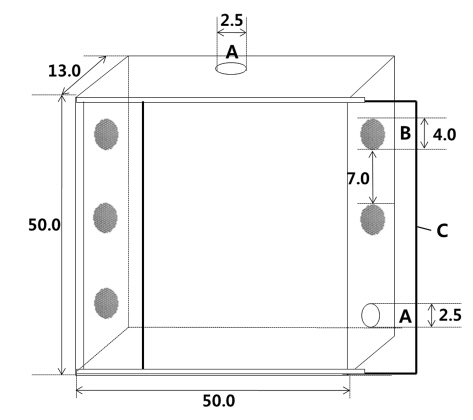
The physical factors of Argiope bruennichi (Araneae, Araneidae) that influence the stabilimentum decoration on the web, a conspicuous white silk structure reflecting much more ultraviolet light than other spider silks in the web, have been poorly understood. In this study, individual variables involved in decorating the webs with stabilimenta by A. bruennichi were examined. The results revealed that the physical condition of the female A. bruennichi affected the behaviors of the stabilimentum decoration on the web. Among the 82 female spiders building their webs, the 49 female spiders adding upper and lower stabilimenta on their web weighed less, and had a narrower cephalothorax and shorter abdomen than the 33 female spiders that did not use stabilimentum. The heavier females decorated their webs with stabilimentum of greater widths. There were also significant positive relationships between the stabilimentum area and the female spider’s cephalothorax width, and between the stabilimentum area and female spider’s abdomen length. Taken together, this study suggests that spiders allocate their resources in stabilimentum decoration as a functional response to the spider's physical conditions, and also supports the “prey-attraction hypothesis,” which states that the use of stabilimentum increases the foraging success by attracting more prey to the web.
Variables influencing the web structures of the spider such as climate factors, prey abundance, competition with the same and other species, predation, individual variables, etc. have been studied (Eberhard 1990, Foelix 2010). However, the spider’s physical factors that influence its decision about whether to add a silk decoration on a web or not have been poorly understood (Eberhard 1990, Schoener and Spiller 1992, Watanabe 1999, Théry and Casas 2009). Many orb-web weaving spiders (Araneae: Araneidae, Tetragnathidae, Uloboridae) decorate their webs with species-typical silk structures, so called stabilimentum (Simon 1895, Herberstein et al. 2000, Bruce 2006).
Stabilimentum is a conspicuous white silk structure that reflects much more ultraviolet light than areas without stabilimentum in the web (Craig and Bernard 1990, Bruce et al. 2005). This is surprising, given that “web-building” spiders’ silk generally has low UV reflectance, presumably to reduce the visibility of the web to insect prey (Blackledge and Wenzel 2000).
Recently, the “prey-attraction hypothesis” has suggested that the stabilimentum functions to increase foraging success by attracting more prey to the web (Craig and Bernard 1990, Tso 1998, Watanabe 1999, Kim et al. 2012). The stabilimentum decoration on the web of the wasp spider,
This study examined whether decorating webs with stabilimenta was affected by individual physical variables in
Adult female
Measurements were conducted on the first web constructed by
To observe the effects of individual variables on the stabilimentum decoration, webs with stabilimenta or without stabilimenta were selected for measurements. For webs with stabilimenta, only webs that had both upper and lower stabilimenta were included.
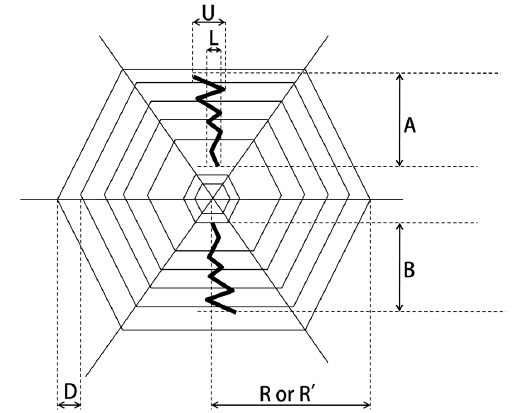
To estimate the web size, the outermost and innermost diameters of each web from the sticky spirals were measured (Fig. 2). The web area was then calculated as: web area= πx longer radius x shorter radius. As the stabilimentum of
Nonparametric statistics in SigmaPlot (version 12.3; Systat Software Inc., San Jose, CA, USA) were used, but the mean and standard deviation of the data were presented instead of medians and inter-quartile ranges on nonparametric statistics. The individual variables (body mass, cephalothorax width, and abdomen length) were compared using the Mann-Whitney rank sum test (Mann and Whitney 1947). Regression analysis with ANOVA was also used to examine the relationship of individual variables and parameters of webs with and without stabilimenta.
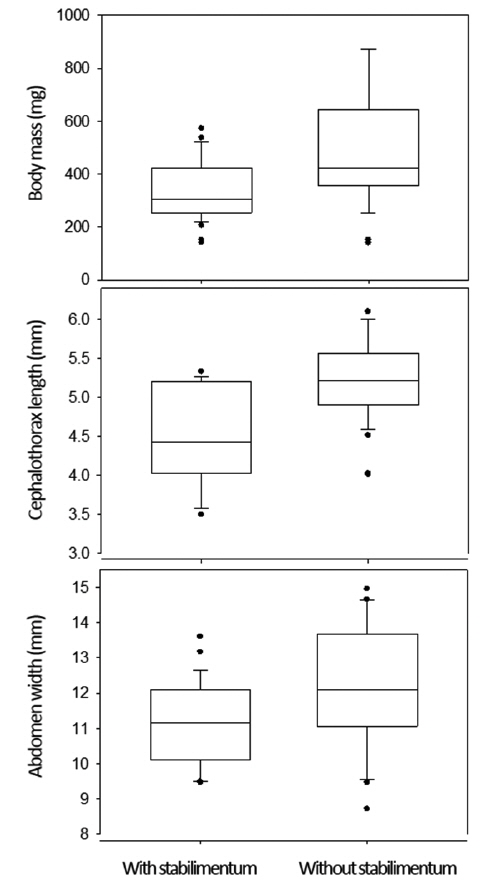
This study investigated the web structures of 82 webs made by female spiders: 49 webs with stabilimenta and 33 webs without. Overall, the body mass of the female spiders (N = 82) was 402.1 ± 174.1 mg (mean ± SD), the cephalothorax width was 4.8 ± 0.7 mm, and the abdomen length was 11.6 ± 1.5 mm. The total web area of the 82 webs was 667.2 ± 285.9 cm2 , the mean radius length was 14.5 ± 3.2 cm, the mesh height was 4.5 ± 1.0 mm, and the number of spirals was 32.5 ± 11.0. The stabilimentum area measured in the 49 webs was 1.76 ± 1.72 cm2 .
There was no significant difference in the web area between webs with or without stabilimenta (Table 1). The mean length of radii and mesh size also did not differ in webs with or without stabilimenta (Table 1). However, the webs with stabilimenta had more spirals on average than the webs without stabilimenta (Table 1). However, the number of spirals showed no positive- or negative relationship with the stabilimentum area (ANOVA test for regression analysis:

Comparison of web structures between webs with stabilimenta and webs without stabilimenta of Argiope bruennichi (mean ± SD)
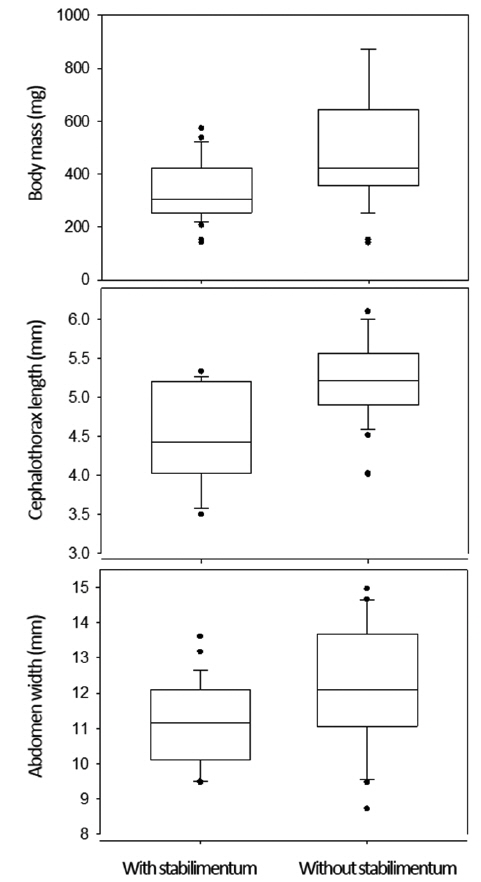
Actually, the stabilimentum decorations reflected the individual variables (Fig. 3). The female spiders that made the webs with stabilimenta weighed lighter than the females that made the webs without stabilimenta (342.0 ± 114.8 mg compared with 491.5 ± 207.8 mg; Mann-Whitney rank sum test, U = 439.0,
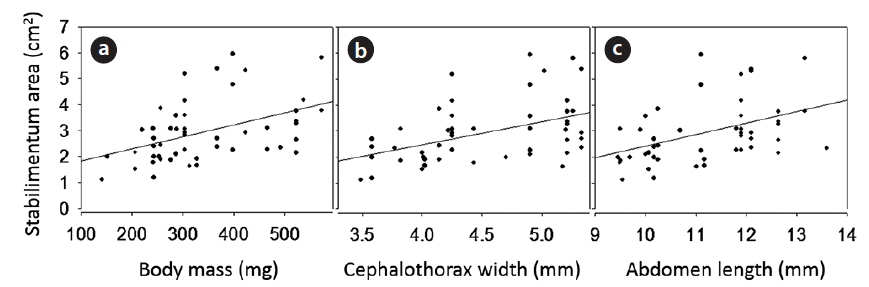
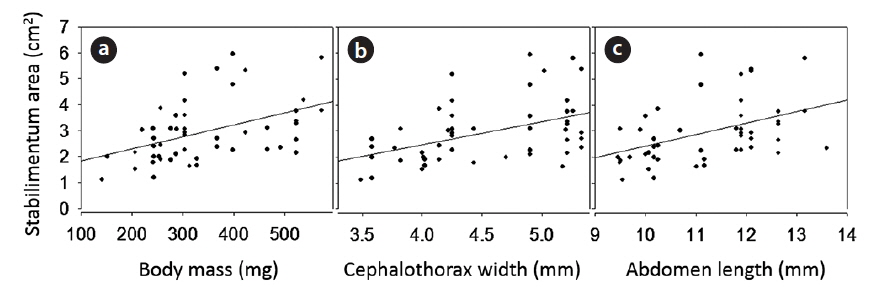
The heavier females decorated webs with wider stabilimentum (Fig. 4A; ANOVA for regression analysis:
The results revealed that the physical conditions of the female
A spider’s decision making on an orb web structure should be associated with its allocation of energy for reproduction or capturing prey (Higgins and Buskirk 1992, Sherman 1994, Herberstein et al. 1998, Watanabe 1999, Théry and Casas 2009). Spiders in very poor physical condition cannot expend much energy for web building. They might not have enough protein for silk production, and thus, it could be difficult to cover the energy costs for web construction. On the other hand, female spiders of a good enough weight to reproduce might not need prey, and therefore may reduce investment of energy in web building to allocate protein and energy to reproduce. But females with average physical conditions could change their investment in web building as a functional response to the changes in the spider’s physical state. Lighter reproductive females should expend more energy in web building to capture more prey, even if this entails the energetically expensive decorating of webs with stabilimentum.
In fact, a previous study on