In recent year, there has been great concern regarding agricultural land uses and their importance for the conservation of biodiversity. Rice fields are managed unique wetland for wildlife, especially waterbirds. A comprehensive monitoring of the waterbird assemblage to understand patterning changes was attempted for rice ecosystem in South Korea. This rice ecosystem has been recognized as one of the most important for waterbirds conservation.
Biweekly monitoring was implemented for the 4 years from April 2009 to March 2010, from April 2011 to March 2014. 32 species of waterbirds were observed. Self-organizing map (SOM) and random forest were applied to the waterbirds dataset to identify the characteristics in waterbirds distribution. SOM and random forest analysis clearly classified into four clusters and extract ecological information from waterbird dataset. Waterbird assemblages represented strong seasonality and habitat use according to waterbird group such as shorebirds, herons and waterfowl.
Our results showed that the combination of SOM and random forest analysis could be useful for ecosystem assessment and management. Furthermore, we strongly suggested that a strict management strategy for the rice fields to conserve the waterbirds. The strategy could be seasonally and species specific.
논습지(rice fields)는 많은 수의 조류가 이용하는 대표적인 인공습지로 잘 알려져 있다(Elphick, 2010). 개발로 인하여 사라져가는 자연습지를 대신하여 논습지는 다양한 수조류에게 서식지를 제공한다(Fasola and Ruiz, 1996; Elphick, 2000). 백로류 같은 일부 수조류 그룹에게 안정적인 개체군 유지를 위해 취식지로서 기능을 가지는 논습지의 분포는 필수적이다(Czech and Parsons, 2002). 논습지의 형태는 지역농업인의 관리에 의해 다양한 형태가 조성되기 때문에 논습지를 이용하는 수조류 군집의 구성이 연중 다양하게 형성된다(Maeda, 2001).
수조류에게 안정적인 서식지 제공을 위해 서식지 관리 전략이 필요하고 이러한 서식지 관리 전략은 체계적인 보전 계획을 통해 수립된다(Breininger and Smith, 1990; Elphick and Oring, 1998). 일반적으로 수조류의 서식지 보전은 저어새
생물 군집의 분포 패턴 파악은 서식지 관리 전략 수립에 중요한 과정이다(Park
본 연구는 한국 중서부지역 논습지에 도래하는 수조류 군집의 분포 패턴을 파악하는데 초점을 두고 있다. 수조류 군집 데이터를 바탕으로 연중 논습지 이용 패턴의 다양성을 평가하였다. 총 4년의 수조류 도래 자료를 자기조직화지도에 접목하여 수조류를 분류하였고 랜덤 포레스트와 지표종 분석(indicator species analysis)을 통해 분류된 수조류 무리의 세부적인 특성을 확인하였다. 계절성뿐만 아니라 공간적 분포를 고려하여 모델의 결과를 해석하였다. 이를 통해 논습지에 도래하는 수조류의 서식지 보전에 관한 유용한 정보를 제공하고자 하였다.
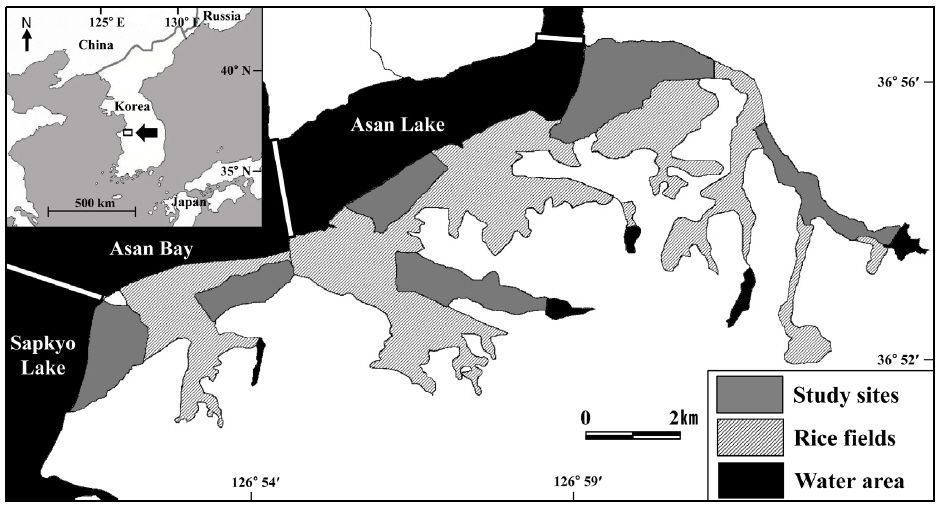
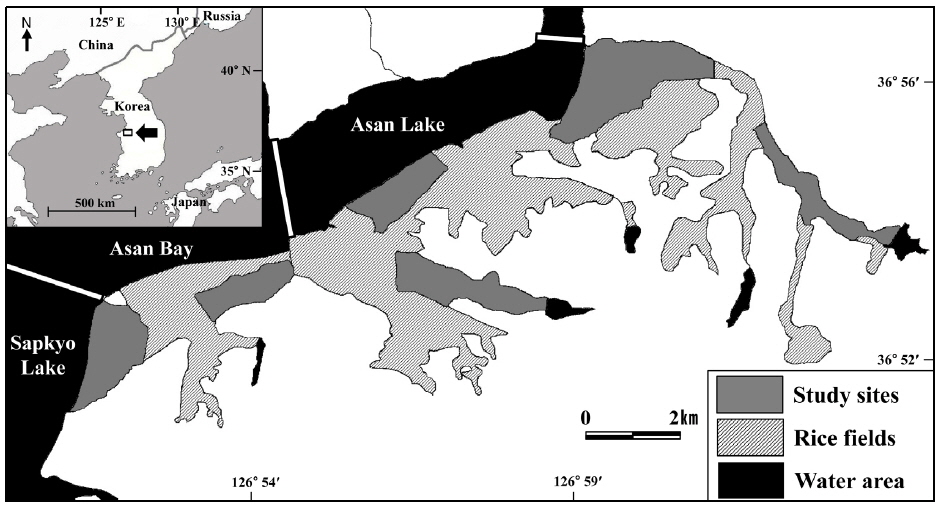
본 연구는 한국 중서부지역에 위치한 아산호와 아산만 남단 지역의 논습지에서 수행되었다(36°54′N, 126°56′E) (Fig. 1). 이 지역은 봄철 이동시기에는 도요물떼새류, 여름철에는 백로류, 겨울철에는 오리기러기류와 같이 다양한 조류가 도래하는 중요한 지역이다. 백로류의 경우 번식 개체군으로 왜가리, 중대백로, 중백로, 쇠백로, 황로, 해오라기 등이 조사 지역 주변에 대단위 번식을 한다(Choi
아산만 남단 지역의 논습지를 모두 조사한다는 건 사실상 불가능하기 때문에 아산만 남단 지역을 대표할 수 있는 6개의 구역을 설정하여 조사를 수행하였다. 각 조사 구역은 약 250개의 필지로 구성되어 있다. 필지는 논둑이나 농로로 나뉘어져 있으며 평균 100 m×45 m 의 크기로 이루어져 있다. 필지는 지역농업인의 관리에 따라 연중 크게 변한다. 4월말이 되면 필지는 수로를 통해 물이 공급된다. 1-2주 뒤 필지는 써레질과 모내기가 이루어지고 9월까지 벼가 성장하여 약 1 m 높이를 형성한다. 9월말에서 10월 중순까지 수확이 이루어지고 수확이 이루어진 후에 필지에 그루터기가 형성된다. 그리고 4월까지 지속적으로 땅을 갈아엎는다.
2009년 4월부터 2010년 3월, 2011년 4월부터 2014년 3월까지 총 4년간 월 2회씩 격주로 수조류 조사를 수행하였다. 조사는 조사 대상 조류가 충분히 활동적인 맑은 날에 오전 6시부터 정오(12:00 pm)까지 실시하였다. 쌍안경(ApoTelevid, Leica, New York, USA)이나 망원경(Ultravid, Leica, New York, USA)을 이용하여 농로를 따라 이동하면서 인접한 필지에 분포하는 조류를 관찰, 동정하였다. 전깃줄이나 전봇대 등과 같은 인공구조물에서 관찰된 조류는 조사 대상에서 제외하였다. 필지의 서식지 타입을 확인할 수 없는 눈이 많이 내린 날에는 조사를 수행하지 않았다.
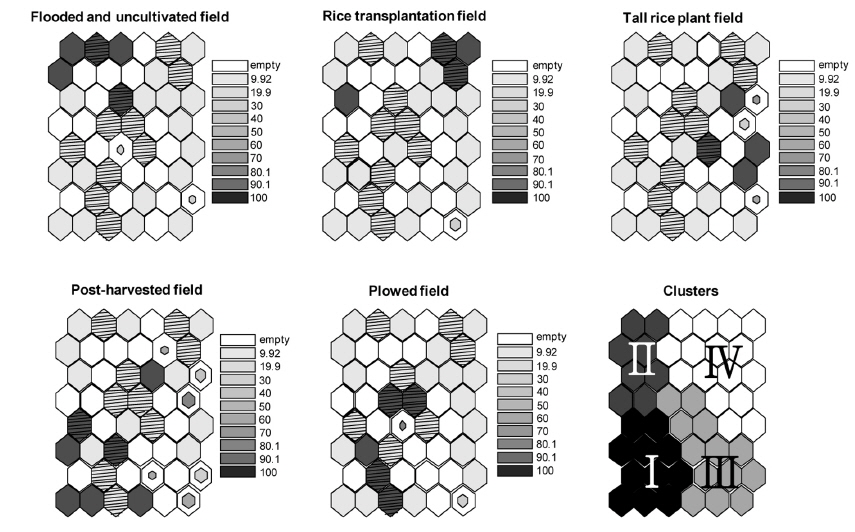
조류 조사 시 관찰된 조류가 이용한 필지의 서식지 타입을 함께 기록하였다. 서식지 타입은 다음과 같이 크게 5가지로 나누었다. (1) 모내기 전 물을 댄 필지(flooded and uncultivated fields, FU), (2) 모내기 한 필지(rice transplantation fields, RT), (3) 벼가 성장한 필지(tall rice plant fields, TP), (4) 수확 후 그루터기 형성 필지(post-harvested fields, PH), (5) 갈아엎은 필지(plowed fields, PL). 서식지 타입은 담수 유무, 토양의 물리적인 상태(평탄화, 논갈이), 벼 식생 성장정도, 수확 후 짚단 처리 방법에 따라 나누었다. “모내기 전물을 댄 필지”의 경우 4월말에서 5월초에 형성되는 형태로 모내기를 위해 물을 댄 필지로 갈아엎은 다음 바로 물을 댄 형태나 물을 대고 써레질을 통해 평탄화 작업을 한 경우 모두를 포함하였다. “모내기한 필지”는 5월초부터 중순 기간 동안 써레질을 통해 평탄화 작업이 끝난 필지에 모를 심은 경우로 정의하였다. “벼가 성장한 필지”는 모내기 후 전체 필지에서 벼의 피복도가 50% 이상인 경우로 하였으며 주로 6월 중순 이후에 나타난다. “수확 후 그루터기 형성 필지”는 10월 초반이나 중반부터 형성되며 논갈이 전 벼를 수확한 필지로 짚단을 그대로 둔 경우와 짚단을 수거한 경우 모두를 포함하였다. “갈아엎은 필지”는 수확 후 논갈이로 필지 표층이 갈아 엎은 형태를 형성하는 경우로 11월부터 3월까지 지속적으로 형성된다.
논습지에 분포하는 수조류 군집의 분류(classifying)와 배열(ordering)을 위해 자기조직화지도를 이용하였다(Kohonen, 1982). 자기조직화지도는 동물집단의 종 분포나 구조를 확인하기 위해 사용하는 분석법으로 탐색적 자료 분석에 효율적이고 강력한 특성을 나타낸다(Chon, 2011). 자율학습 신경망(unsupervised neural network)인 자기조직화지도는 계층적인 시스템이 아니며, 일반적으로 입력층과 출력층으로 구성되어 있다. 출력층은 통상 육각형의 격자단위인 뉴런으로 구성된다. 모든 연결들은 입력층에서 출력층의 방향으로 되어있으며 각각의 뉴런은 출력결과를 전혀 예측할 수 없는 자율학습을 통해 갱신되는 연결강도를 가진다. 이 연결강도는 입력 패턴과 가장 유사한 출력층 뉴런이 승자가 된다. 출력층 뉴런은 승자 독식 구조로 승자만이 출력층 뉴런에 나타나며, 승자와 유사한 연결강도를 갖는 입력 패턴이 동일한 출력 뉴런으로 배열된다. 출력층의 뉴런은 일반적으로 2차원으로 보여준다.
관찰된 각 수조류 종 (총 32종×92회 조사 자료)의 개체수는 각 종별 개체수 차이가 크게 나타났기 때문에 로그치환하여 자기조직화지도의 입력층에 할당하였다. 32종은 조사기간 동안 논습지에서 관찰된 전체 수조류 종을 의미하고 92회 조사는 눈이 많이 내리는 4회의 조사시기를 제외한 조사 횟수를 의미한다. 출력층 뉴런의 크기는 가로 6개, 세로 8개로 나타냈다. 출력층 뉴런의 크기는 휴리스틱 규칙(heuristic rule)에 의해 선택되었다(Vesanto
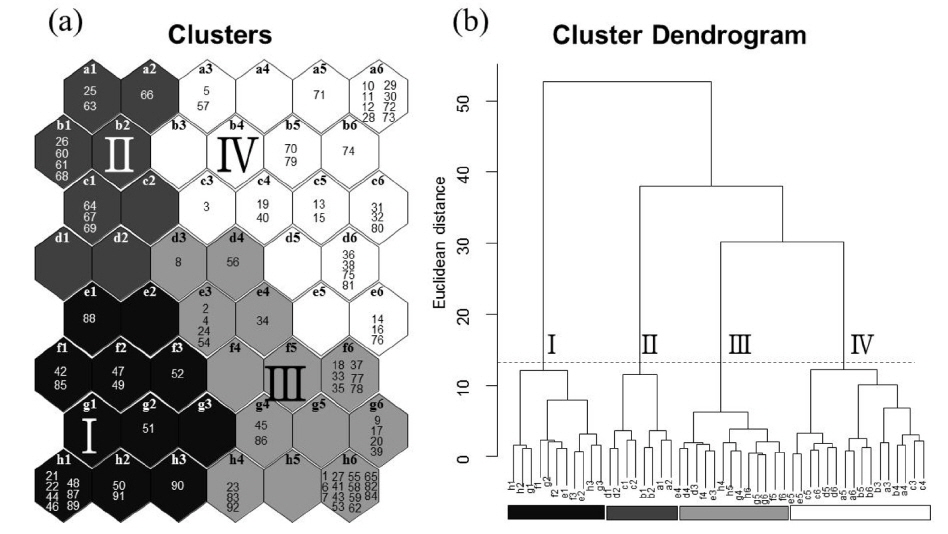
자기조직화지도의 자율학습 후 각 뉴런이 가지는 유사성을 이용하여 각 그룹을 분류하였다. 뉴런의 거리 측정은 Euclidean distance를 사용하였고 그룹의 분류는 Ward linkage 방법을 이용하였다(Park
자기조직화지도의 분석은 R 통계프로그램의
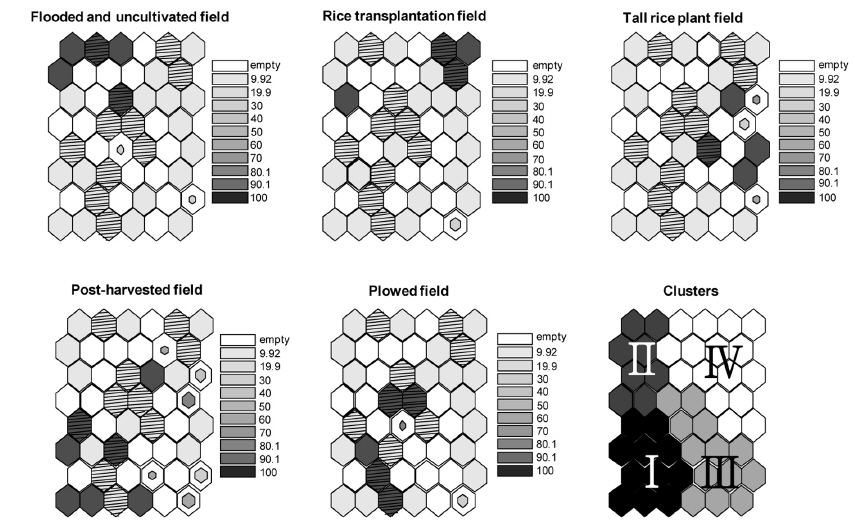
자기조직화지도에 의해 표현된 출력층의 뉴런에 영농법의 중요성을 나타냈다. 각 뉴런 단위 별 영농법의 평균 수치를 grey scale과 뉴런의 크기로 표현하였다. 중요성은 비율 자료로 표현하였으며 짙은 색으로 갈수록 영농법의 중요성이 높음을 의미한다. 뉴런의 크기는 뉴런 단위 별로 계산된 분산값의 역의 비율로 나타냈다. 즉, 뉴런의 크기가 작을수록 분산값이 큼을 의미한다.
자기조직화지도에 의해 나누어진 각 그룹을 대표하는 종을 분석하기 위해 지표종 분석을 수행하였다. 지표종 분석은 출현한 종들의 상대 밀도와 상대 빈도의 곱으로 계산된 지표값(indicator value, IndVal)을 통해 지표종을 평가하는 방법이다. 지표값의 유의성을 결정하기 위해서 몬테카를로 검증을 사용하였다. 총 1,000번의 시뮬레이션을 통해 유의성을 확인하였다.
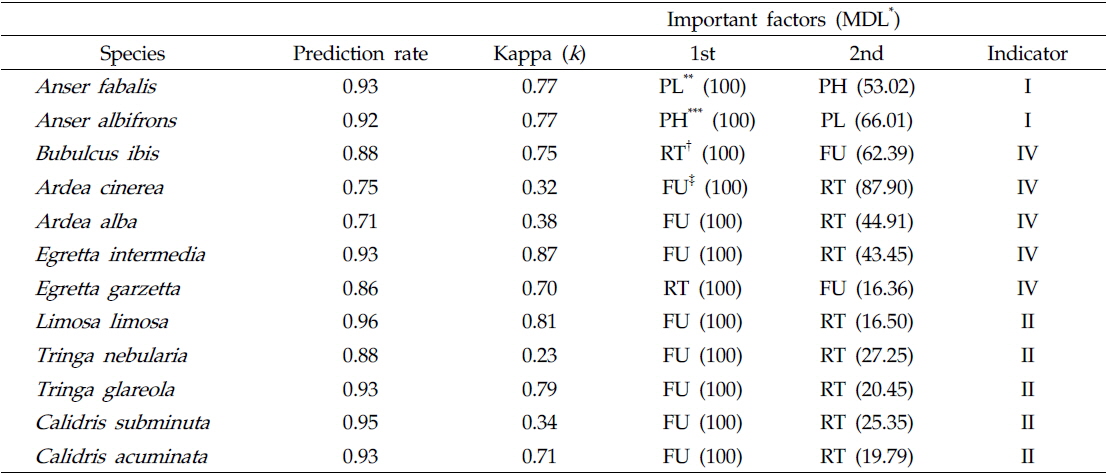
각 그룹과 영농법의 관련성을 확인하기 위해 랜덤 포레스트 모델을 사용하여 할당된 그룹을 평가하였다. 영농법에 따라 출현한 조류 종들을 이분형 자료(0과 1)로 나타냈으며 이를 이용하여 램덤 포레스트 모델에 적용하였다. 랜덤 포레스트 모델은 다수의 분류 나무를 생성시켜 이용하는 방법으로 CART(classification and regression trees)의 불안전성을 보완하고 있으며 정확한 분류를 할 수 있는 학습 알고리즘 중 하나로 간주된다. 랜덤 포레스트 모델은 학습과정을 통해 각 인자의 중요도를 계산한다. 이는 각 분류 나무 마디마다 변수값들이 분류의 정확성에 미치는 정도를 기초로 하여 측정되는 값으로, 변수의 중요도를 파악하는데 유용하게 이용된다. 랜덤 포레스트 모델의 중요도는 minimum description length (MDL)로 나타냈다. MDL의 경우 분류에서 각 변수의 중요성을 서열화로 보여주는 값으로 0에서 100까지 범위의 수치로 변환하여 사용했으며, 모델의 적합성은 Prediction rate와 Kappa 값(
수조류 그룹(도요물떼새류, 백로류, 수금류)별 종수와 개체수의 차이는 Kruskal-Wallis (K-W) 검증을 이용하였고, 사후검증으로 Nemenyi-Damico-Wolf-Dunn 검증을 이용하였다. 모든 분석은 R 통계 프로그램을 이용하였다(R Development Core Team, 2012).
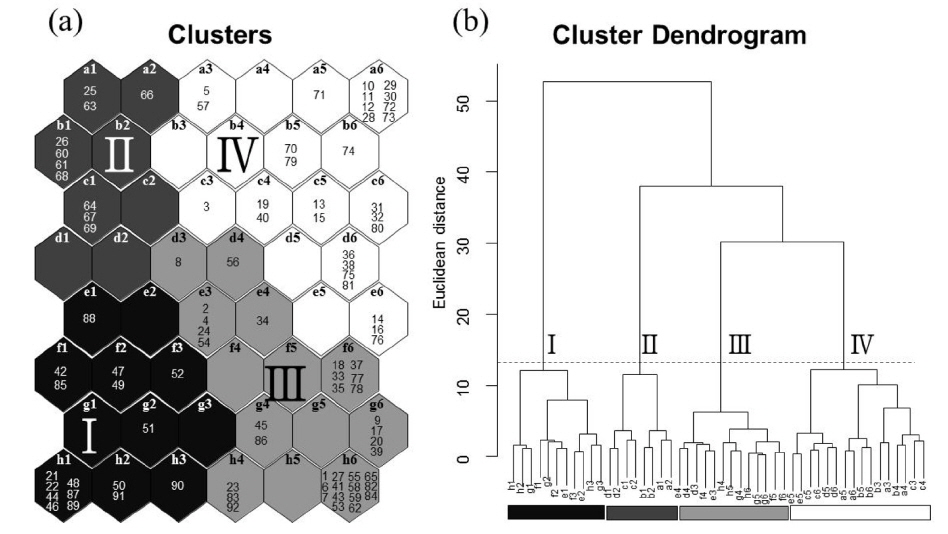
논습지에서 관찰된 32종의 수조류를 기초로 한 92회의 조사 자료를 자가조직화지도에 적용하여 분석한 결과 크게 4개의 그룹(I-IV)으로 나눌 수 있었으며, 이들 그룹들 간에는 유의한 차이가 나타났다(Fig. 2; MRPP, A=0.37,
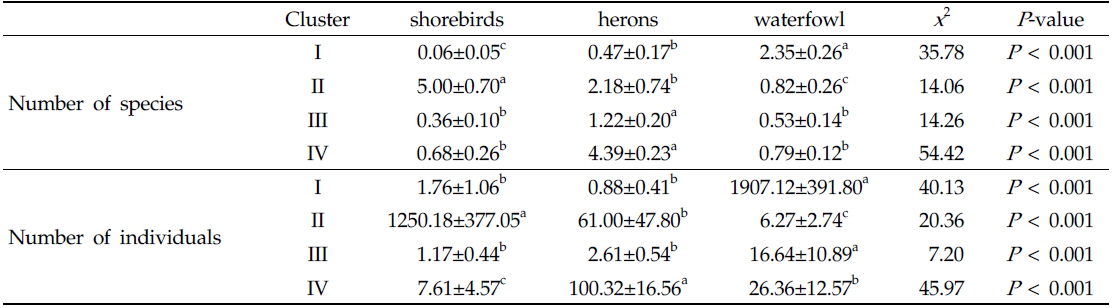
자기조직화지도 분석에서 각 수조류 그룹별 차이는 Table 1에 나타냈으며, 수조류 그룹에서 차이를 잘 반영함을 알 수 있었다. 그룹 I이 위치한 자기조직화지도 결과의 좌측 하단부는 오리기러기류의 종수와 개체수가 높게 나타났고(종수:
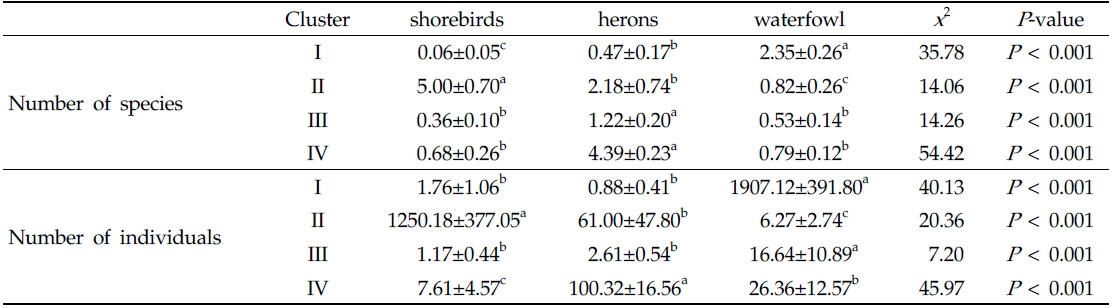
Comparison of number of species and individuals in relation to self-organizing map clusters. Data are expressed as mean±S.E. Different letters (a, b, c) indicate significant differences among clusters based on Nemenyi-DamicoWolf-Dunn test (P < 0.05)
그룹 I의 경우 마른 논의 형태인 수확 후 그루터기를 형성하는 필지와 갈아엎은 필지가 많이 나타났고 그룹 II, III, IV는 담수형태의 논습지의 형태가 많이 분포하는 것을 확인 할 수 있었다(Fig. 3). 특히 그룹 II는 모내기 전 물을 댄 필지가 주를 이루었으며, 그룹 IV는 모내기 한 필지나 벼가 성장한 필지가 우세하게 분포하였다.
겨울철 논습지를 이용하는 큰기러기
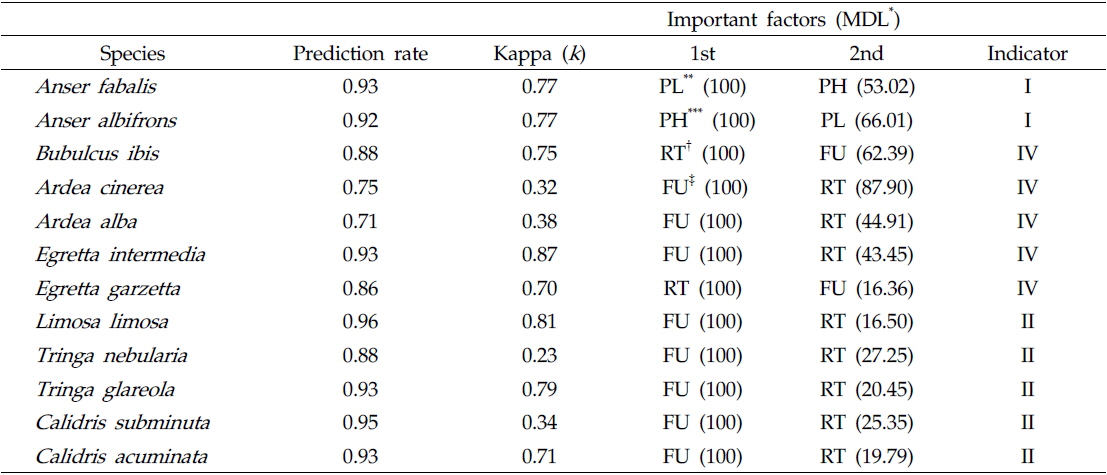
Prediction of waterbird species and evaluation of important feature of farming practices for each indicator species using random forest model. Indicator is based on the IndVal in indicator analysis
본 연구 결과를 통해 논습지를 이용하는 수조류 군집의 분포 패턴은 연중 다양한 형태로 나타나고 각 시기별로 구성된 수조류 군집은 필지의 서식지 타입과 밀접한 관련이 있었다. 논습지는 영농방법에 따라 필지의 상태가 담수논과 건논이 반복하여 나타난다(Elphick, 2010). 이러한 변화 양상이 결국 수조류 군집 분포에 결정적인 요인으로 작용하는 것을 확인할 수 있다(Nam
모내기 전에 이루어지는 논습지의 담수는 수심이 평균 약 10cm 내외로 유지된다(Nam
그룹 III에 나타난 수조류 무리는 논습지에 도래하는 수조류 군집의 강한 계절성을 보여 주며, 필지 타입의 변화의 시점에는 수조류의 이용성이 낮아지는 것으로 나타났다. 그룹 III에서 확인된 도요물떼새류, 백로류, 오리기러기류의 종수와 개체수가 낮았고, 상대적으로 그 차이가 작다(Table 1과 Fig. 2). 또한 특정 필지 타입이 뚜렷이 나타나기 보다는 여러 필지 타입이 분포하는 특성을 가진다(Fig. 3). 따라서 그룹 III의 수조류 무리 특성은 도요물떼새류, 백로류, 오리기러기류와 같은 수조류 그룹들의 도래가 끝나거나 다시 시작되는 시기로 판단된다. 논습지는 인간에 의해 물리적 화학적 요인이 급격히 변하기 때문에 논 생태계는 높은 역동성을 가진다(González-Solís
자연습지와 비교하여 논습지는 공간의 이질성이 적은 단순한 형태를 가진다(Elphick, 2010). 단일 식물 종(벼
자기조직화지도와 랜덤 포레스트 모델, 지표종 분석의 통합적인 사용은 데이터를 탐색적인 접근으로 생태적 정보를 추출하기에 효율적이었다(Bae
본 연구는 논습지에 서식하는 수조류의 시공간적 분포 특성을 확인하기 위해 격주로 조사된 4년간의 수조류 군집 자료를 이용하였다. 총 32종의 수조류가 논습지에서 관찰되었으며, 이들은 뚜렷한 계절성과 함께 논습지에 조성된 공간에 대한 수조류 그룹의 이용 패턴에 차이가 있음을 확인 할 수 있었다. 수조류 군집 자료를 자기조직화지도와 랜덤 포레스트 모델, 지표종 분석에 적용한 결과 명확한 결과를 도출할 수 있었다. 논습지는 담수의 유무로 수조류 그룹이 크게 차이가 있음을 확인하였고 담수논에서도 식생의 밀생 정도에 따라 수조류 그룹의 이용 형태가 달라지는 것을 알 수 있었다. 이러한 수조류 그룹의 서로 다른 분포 패턴은 결국 각 수조류 그룹의 서식지 선호성과 높은 관련성을 가진다.