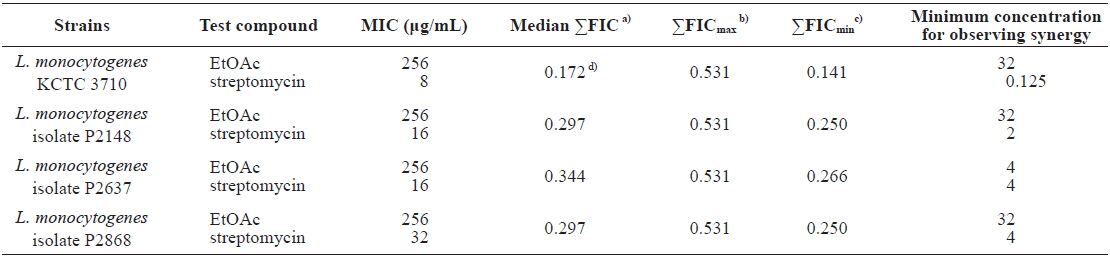
In an effort to discover alternative antimicrobials against Listeria monocytogenes, several marine algae were screened. The methanolic extract of Ecklonia cava exhibited the highest antibacterial activity against L. monocytogenes, with the ethyl acetate (EtOAc) soluble fraction of E. cava methanolic extract having a MIC value of 256 μg/mL and a MBC value of 512 μg/mL. The MIC values of streptomycin in combination with the EtOAc fraction were markedly reduced up to 64-fold, suggesting that the antibacterial activity of the antibiotic was restored when combined with the EtOAc fraction. The interaction between streptomycin and the EtOAc fraction was assessed by fractional inhibitory concentration (FIC) indices. The combination of streptomycin and the EtOAc fraction against L. monocytogenes resulted in ΣFICmin range of 0.141 to 0.266 and ΣFICmax of 0.531 for all strains. The median ΣFIC against L. monocytogenes strains ranged from 0.172 to 0.344. Thus, synergistic ranges of FIC <1 were observed for all combinations of streptomycin and the EtOAc fraction against L. monocytogenes strains. Indeed, the median ΣFIC of streptomycin–EtOAc fraction ranged from 0.172 to 0.344, suggesting a marked synergy.
Recently, consumers have begun demanding foods that are fresh, natural, and minimally processed, along with the requirement for enhanced safety and quality. This perspective has put pressure on the food industry for progressive removal of chemical preservatives, and has fuelled research into alternative natural antimicrobials (Lanciotti et al., 2004). Additionally, increased public awareness of the negative effects caused by synthetic chemicals has led to the search for “green solutions,” such as organic and synthetic chemical-free food products (Abutbul, 2004).
In that regard, seaweeds have become an important source of pharmacologically active metabolites, with a broad spectrum of biological activities with possible applications in food production to prevent bacterial and fungal growth. In fact, compounds with antioxidant, antiviral, antifungal, and antimicrobial activities have been detected in brown, red, and green algae (Eom et al., 2011).
Dried
The contents of total phenolic (TP) compounds in the fractionated
>
Microbial strains and cultures
The bacterial strains used in this study were
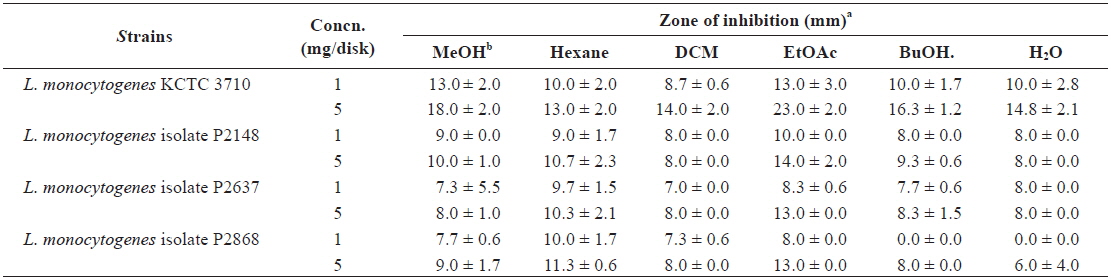
The antibacterial efficacy of marine algae extracts was evaluated by disk diffusion assay followed by the CLSI (2009), as described in our previous reports (Lee et al., 2014; Eom et al., 2011). In brief, bacterial strains were cultured in BHI at 37℃ until an OD at 600 nm of 0.5. One hundred microliters of bacterial culture containing approximately 104 - 105 CFU/ mL were spread on Mueller Hinton agar (MHA; Difco, USA) plates. Paper discs (6 mm in diameter) containing 1 and 5 mg of extract were placed on the MHA plate. After incubating for 24 h at 37℃, the diameter of the inhibition zone was measured. The experiment was carried out three times and the mean values were calculated.
>
Measurement of minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC)
Minimum inhibitory concentration (MIC) is defined as the lowest concentration of antimicrobial that inhibits visual growth of microorganisms after 20 - 24 h of incubation at 37℃ (Gierson and Afolayan, 1999). The MICs of marine algae extracts and some commercial antibiotics were determined by a twofold serial dilution method in Mueller–Hinton broth (MHB; Difco) (CLSI, 2008).
Minimum bactericidal concentration (MBC) is defined as the lowest concentration of an antimicrobial required for a 99.9% reduction in the viable cell population (NCCLS, 2003). For MBC determination, an aliquot of inoculums was taken from a MIC test well that did not show turbidity and was poured onto BHI agar plates. The agar plates were incubated at 37℃ until growth was seen in the growth control plates. The number of colonies on each agar plate was counted. The MIC and MBC experiments were repeated in triplicate.
>
Antibiotic susceptibility test
Sensitivity and/or resistance of
>
Synergic effects between EtOAc fraction of E. cava and antibiotics
The interaction between the EtOAc fraction of
Data are reported as means ± standard errors of the mean. Differences at
>
Antibacterial activity of MeOH extracts from 13 marine algae
MeOH extracts from 13 marine algae, including
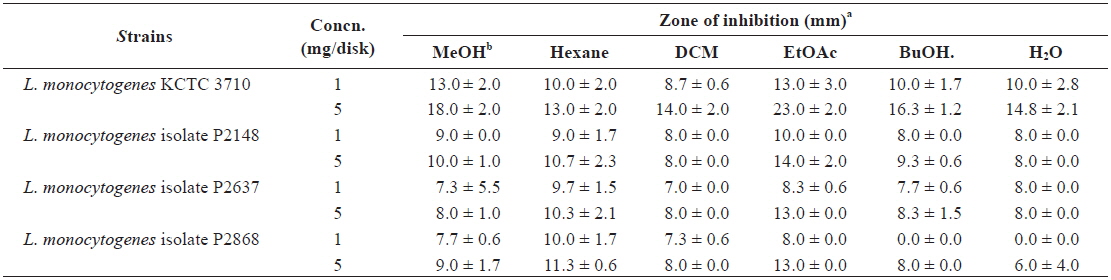
Disk diffusion assay of methanolic extract and its fractions from Ecklonia cava against Listeria monocytogenes strains
To further investigate the mechanism underlying the antibacterial effect against
These results suggested that a substance with antibacterial activity against
>
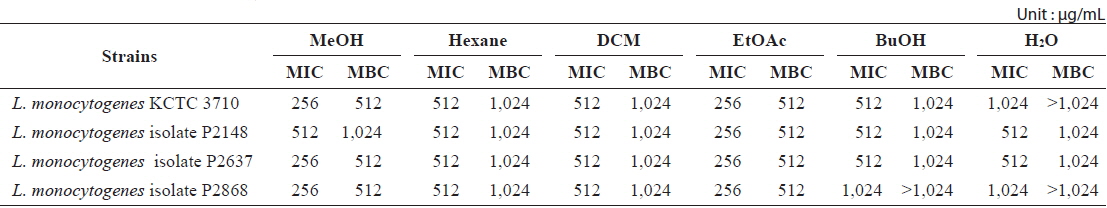
Determination of MIC and MBC of E. cava extract
The MIC assay was carried out to quantitatively evaluate the antibacterial activity of
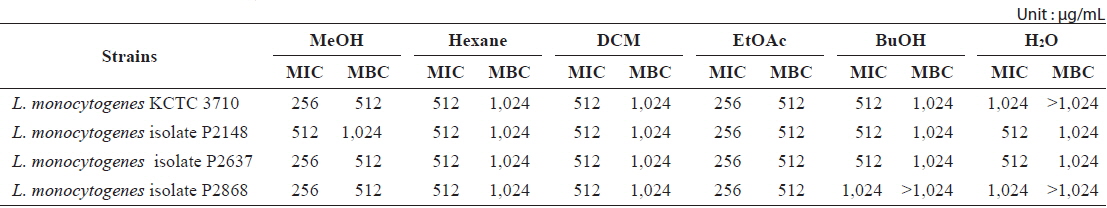
Minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) of the MeOH extracts and its soluble fractions from Ecklonia cava against Listeria monocytogenes strains
The antibacterial activities of
>
Antibiotic resistance of L. monocytogenes strains
Recent reports show an increased rate of resistance to one or more clinically relevant antibiotics in environmental
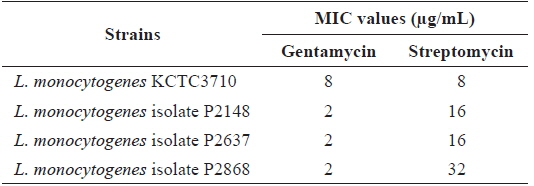
The antibiotic resistance patterns of the type strain and three clinical isolates of
The MIC value for streptomycin against
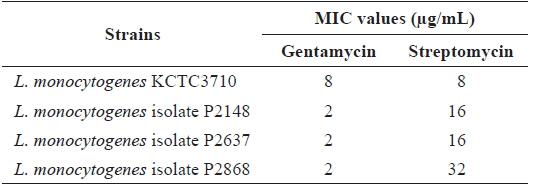
Minimum inhibitory concentration for streptomycin and gentamycin against Listeria monocytogenes strains
>
Synergistic effect between EtOAc fraction of E. cava and streptomycin against L. monocytogenes
It has been demonstrated that one of the more effective strategies in developing new drugs or alternative therapies is the restoration of antibiotic activity, in combination with antibacterial materials derived from natural products and traditional medicines, against drug-resistant bacteria (Eom et al. 2014; Lee et al., 2014). In this study, the interaction between the EtOAc fraction of
As shown in Table 4, the MIC values of streptomycin in combination with the EtOAc fraction of
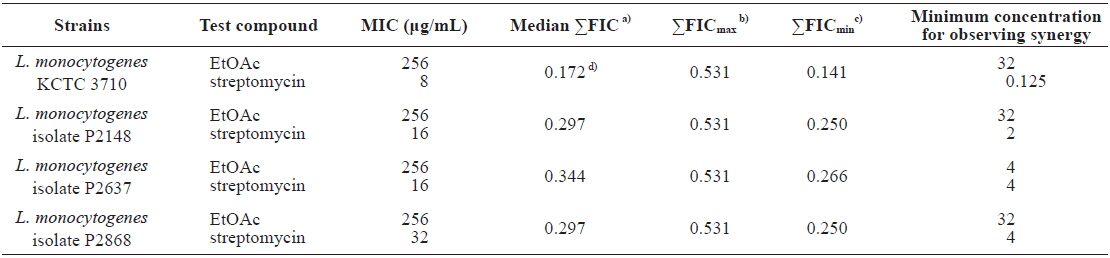
Minimum inhibitory concentrations (MIC) and fractional inhibitory concentration (FIC) indices of the ethyl acetate (EtOAc) fraction of Ecklonia cava in combination with streptomycin against Listeria monocytogenes strains
The synergistic antibacterial activity between streptomycin and the EtOAc fraction of
In conclusion, this study evaluated the antibacterial activity of the edible marine brown alga