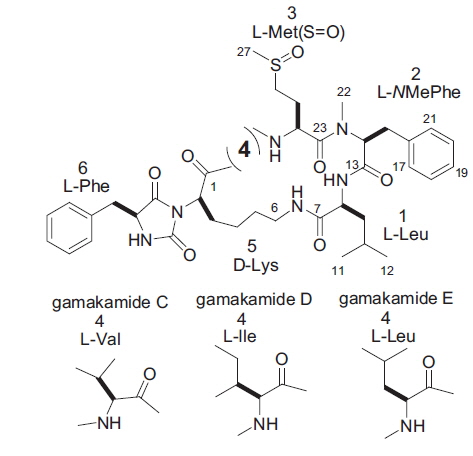
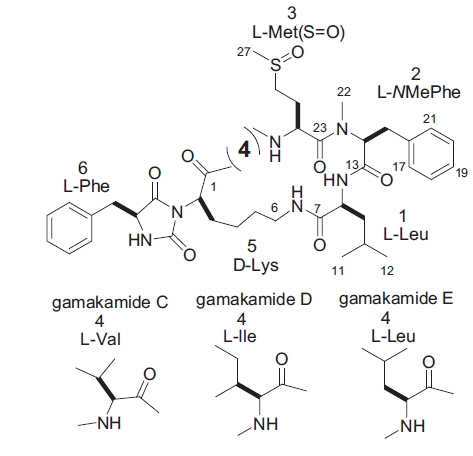
Two new bitter-tasting cyclic peptides comprising six amino acids, namely gamakamide C and D, were isolated from cultured oysters Crassostrea gigas. Dimethylaminoazobenzene sulfonyl–amino acid analysis detected Val and Leu in gamakamide C and Ile and Leu in gamakamide D. The molecular formula of gamakamide C was determined as C43H60N7O8S by high-resolution fast atom bombardment mass spectroscopy (HR FAB-MS) ([M + H]+ m/z 822.4200 Δ –2.4 mmu), and that of gamakamide D was determined as C43H62N7O8S by HR FAB-MS ([M + H]+ m/z 836.4379 Δ –2.0 mmu). Comparison of amino acid analyses and fragment ions by MS/MS among gamakamide C, D, and E (known), the structures of gamakamide C and D were confirmed as –ʟ-Val-ʟ-Met(SO)-ʟ-NMe-Phe-ʟ-Leu-ᴅ-Lys-ʟ-Phe– and –ʟ-Ile-ʟ-Met(SO)-ʟ-NMe-Phe-ʟ-Leu-ᴅ-Lys-ʟ-Phe–, respectively.
The oyster
Oysters take up and accumulate nutrients from plankton and organic matter in the water by filter feeding. Therefore, oyster quality depends on the cultivation conditions, such as the quantity and species of plankton available, degree of pollution, and water quality. The discoloration of oyster meat, such as greening of the intestine by chlorophyll (Kimura, 1969), red coloration (Hata et al., 1982), and yellowish coloration (Hata et al., 1987), occurs due to the plankton on which the bivalves feed. Hatano et al. (1990) also reported an unacceptable taste and coloring of cultured oysters caused by some plankton in Japan. Furthermore, Parry et al. (1989) reported a bitter taste in cultured oysters during a bloom outbreak of the diatom
There have been no previous reports regarding the deterioration of oyster quality due to plankton in the natural setting, but shellfish farmers commonly believe that sudden changes in flavor or color are due to the feed plankton. Deterioration of the quality of cultured shellfish, including oysters, occurs sporadically every year. Therefore, it is necessary to monitor and control the feed plankton and water quality in culture farms during the harvest season for maintenance of both good quality and good food hygiene.
Lee (1995) first reported bitter taste compounds contained in oysters cultured in Gamak Bay located on the southwestern coast of Korea. All of the oysters harvested at that time from Gamak Bay had a very bitter flavor, and even when smoked, boiled, and canned, these oysters remained bitter. Although no reports of illness followed the consumption of these bitter oysters, the unpleasant flavor adversely affected local oyster processing companies.
Five bitter peptidic compounds, tentatively named gamakamides A to E (named after Gamak Bay), were isolated from the bitter-tasting oysters collected at Gamak Bay (Lee, 1995). The structure of gamakamide E was subsequently determined by spectrometry (Lee et al., 2012). We herein report the structures of two new analogues, gamakamides C and D.
Bitter taste compounds were separated and isolated from the side fractions of cultured oyster
Constituent amino acids of gamakamide were evaluated by dimethylaminoazobenzene sulfonyl (DABS)-amino acid analysis after acidic hydrolysis (Syu et al., 2008). Aliquots of 20 μL of methanolic gamakamide C, D, and E solutions (50 μg/mL) were added to separate sample tubes and dried under a stream of nitrogen gas. The tubes were set on a hydrolysis rack after addition of 700 μL of 6 N HCl solution, and the samples were hydrolyzed for 24 hours at 110℃. Chlorine was removed under vacuum, and 20 μL of sodium hydrogen carbonate buffer (NaHCO3, pH 9.0) was added to the hydrolysate and mixed with 40 μL of DABS (1 mg/mL in acetonitrile) reagent and reacted for 12 minutes at 70℃. The reaction was stopped in an ice bath, and the solutions were diluted with 440 μL of sodium hydrogen phosphate buffer (pH 9).
DABS-amino acids were analyzed by high-performance liquid chromatography using an Ultrasphere DABS column (5 μm, 250 × 4.6 mm; Beckman Coulter, Fullerton, CA) at a wavelength of 436 nm. Solvent elution was A: 4% DMF in 10 mM citric acid buffer (pH 6.5) and B:A (3): 4% DMF in acetonitrile (7) ratio. Gradient elution (flow rate, 1.4 mL/min) was carried out starting with 25% B to 56% B solution over 17.2 minutes, followed by gradient elution of 86% B for 23.2 minutes, with a final change to 100% B solution after 29.2 minutes.
Low-resolution fast atom bombardment-mass spectrometry (FAB-MS) and high-resolution FAB-MS (HR FAB-MS) of gamakamides were performed in positive ion mode using glycerol as the matrix on a JEOL JMS 700 MS spectrophotometer (JEOL, Tokyo, Japan).
MS/MS measurements of gamakamides were based on collision-induced dissociation occurring in the collision cell (Q2) of the triple quadrupole at a collision energy of 40 eV using an API2000 mass spectrometer (MDS SCIEX; Applied Biosystems, Foster City, CA) equipped with an electrospray ionization source in positive-ion mode. Electrospray ionization was evoked by a spray voltage of +5.5 kV, and the heated capillary temperature was maintained at 500℃. Nitrogen was used as the target gas. Gamakamide D (5 μL in methanol) was introduced into the mass spectrometer by flow injection with methanol at a flow rate of 0.2 mL/min.
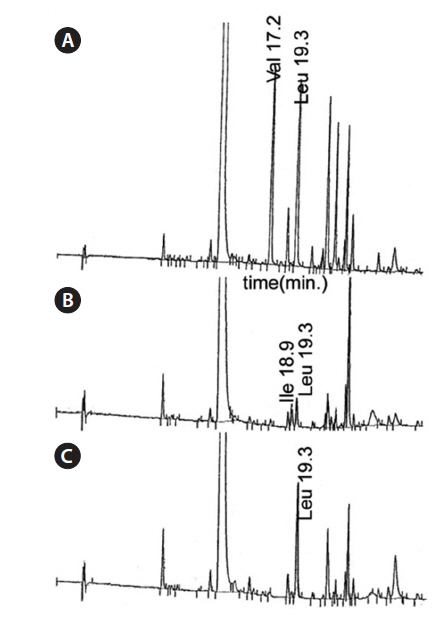
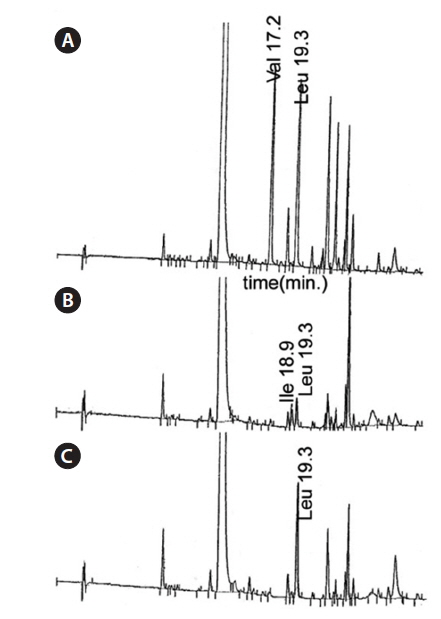
The amino acid constituents of the gamakamides were investigated by highly sensitive DABS-amino acid analysis after hydrolysis of amino acids with HCl and comparison with the retention times of authentic amino acids. DABS-Val (17.2 min) and DABS-Leu (19.3 min) were detected at a ratio of 1:1 in gamakamide C (Fig. 1A). Gamakamide D showed DAB-Ile (18.9 min) and DAB-Leu (19.3 min) at a ratio of about 1:1 (Fig. 1B), while only a large peak of DAB-Leu (19.3 min) was detected in gamakamide E (Fig. 1C). These results indicate that in contrast to gamakamide E, which contains only two Val residues, gamakamide C contains Val and Leu while gamakamide D contains Ile and Leu.
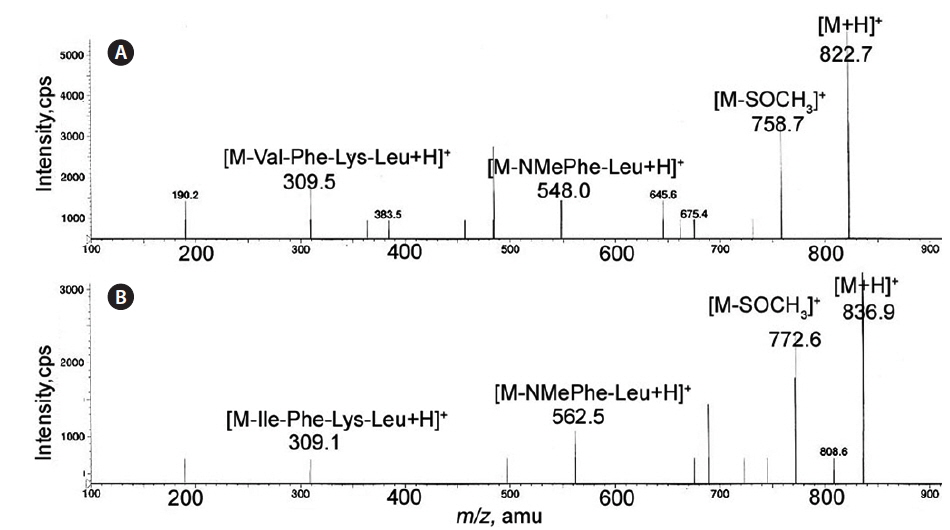
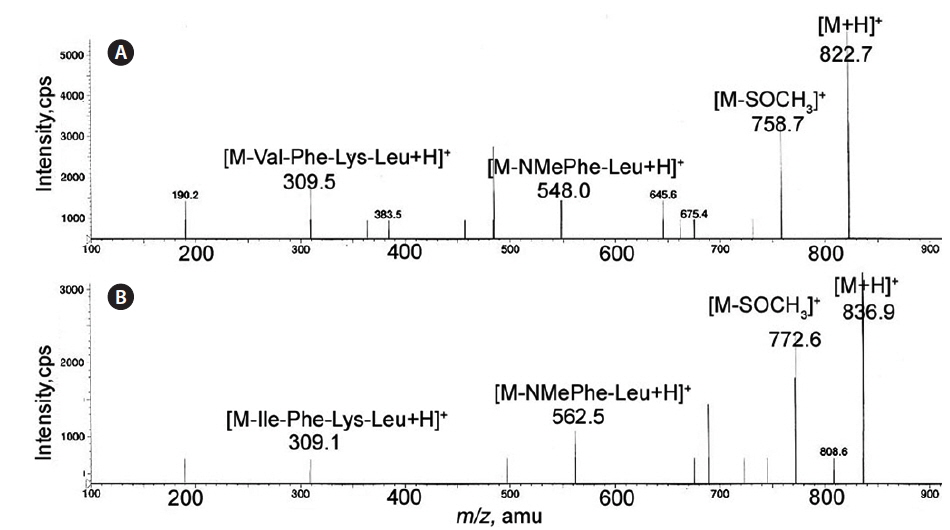
Val was also confirmed in gamakamide C on the mass spectrum. On FAB-MS measured in positive-ion mode, [M + H]+ ion peaks were detected at
In gamakamide D and E, [M + H]+ ion peaks were detected at the same mass unit,
Two Leu residues were connected at positions 1 and 4 in gamakamide E, as shown in Fig. 3 (Gamakamide E), which was confirmed by nuclear magnetic resonance and MS analyses (Lee et al., 2012). Comparing the MS and MS/MS data of these gamakamides and the arrangement of amino acids in gamakamide E, structural differences among gamakamide C, D, and E were presumed to only exist at position 1 or 4 (Val in gamakamide C and Ile in gamakamide D). Therefore, gamakamide C possesses Val and Leu at position 1 or 4, respectively, while gamakamide D possesses Ile and Leu at these positions.
Fig. 2 shows a comparison of the MS/MS spectra of gamakamide C and D. The [M – S(O)CH3]+ ion peak showed
The stereochemistries of Val in gamakamide C and Ile in gamakamide D were presumed to be of the l-type because of confirmation of a Lys in the gamakamide E peptide as l-type by Marfey’s method (Lee et al., 2012). Based on these results, the structure of gamakamide C was presumed to be a six-membered cyclic peptide, –ʟ-Val-ʟ-Met(SO)-ʟ-
Hydrophobic or branched amino acids, such as Phe, Tyr, Arg, Leu, Ile, Val, and Met, are known to give a bitter taste (Food, 2015). Therefore, it was presumed that Phe, Val, Leu, and Ile, alone or in some combination, in gamakamides produce a bitter taste. Enzymatic hydrolysis or processing of protein using endopeptidase frequently leads to the production of an undesirable bitter taste due to the production of hydrophobic oligopeptides, including the bitter amino acid residues described above (Kim, 2013).
Separate from these changes during metabolism, bitter compounds were presumed to accumulate in the digestive organs of oysters because of their consumption of plankton present in seawater. Therefore, it is necessary to identify the plankton responsible for the bitter taste of the oyster meat.
Many cyclic peptides with strong and varied biological activities have been isolated from sponges and tunicates (Fusetani and Matsunaga, 1993) but rarely from shellfish (Reese et al., 1996). Although gamakamides C, D, and E have yet to be found to have any biological activity except bitter taste, they are expected to have some activity due to their structure. Hydantoin, which is similar to gamakamide, has a unique imidazolidine 2,4-dione with biological activity. Many chemical synthesis methods for hydantoin groups have been used for production of pharmaceuticals and pesticides (Hydantoin, 2015).
The structure of gamakamides is similar to that of keramamides, anabaenopeptins, and oscillamides isolated from sponges and cyanobacteria with a ureido carbon and ᴅ-Lys in the molecule (Harada et al., 1885; Nagasawa et al., 1991; Marsh et al., 1997). However, little is known about the biosynthesis of the hydantoin ring in nature. Further studies using living cells are required to determine the mechanism of synthesis of the hydantoin structure of gamakamides, with a terminal carboxyl group and amino group of Phe and amino group of ᴅ-Lys.