The family Diodontidae of the order Tetraodontiformes comprises 19 species in 6 genera worldwide (Nelson, 2006), with 2 species in 2 genera in Korea (Kim et al., 2005) [Diodon holocanthus Linnaeus, 1958; Chilomnycterus affinis (Linnaeus, 1758)]. Diodontid fishes are commonly known as porcupinefishes, burrfishes, and spiny puffers (Gallo et al., 2009). They are able to inflate the body, an important defense mechanism for such slow moving species (Cavaleri, 2000; Leis, 2006; Gallo et al., 2009). The body is covered with well-developed sharp spines, and species are identified according to whether the spines are movable or immovable (Leis, 1986; Aizawa, 2002; Leis, 2006). Some diodontid fishes are poisonous, having tetradotoxin in their ovaries (females) and liver, and can also cause ciguatera (Allen and Erdmann, 2012; Bandyopadhyay, 2013). Most are generally scarce due to their solitary habits; they are found on soft bottoms, on rocky or coral reefs as well as coastal trawling grounds (Leis, 2001; Allen and Erdmann, 2012). Consequently, diodontid fishes have high bycatch rates by trawling and their recovery capacity is low (Stobutzki, 2001). Although Diodontidae fishes are not usually sold commercially for consumption, they are popular in the marine aquarium trade (Leis, 2001; Ellis, 2006). They are a conspicuous and readily captured species, and have been the focus of interest by naturalists for some time (Leis, 2006). However, in Korea, there have been few studies on diodontid fishes other than the brief description of the two species by Kim et al. (2005).
During a survey of fish resources around Jeju Island, Korea, a total of six specimens (57.03-100.72 mm SL) were collected, that were identified as Cyclichthys orbicularis, a new record to the Korean fish fauna. We herein provide morphological and molecular traits of the species and suggest a new Korean name.
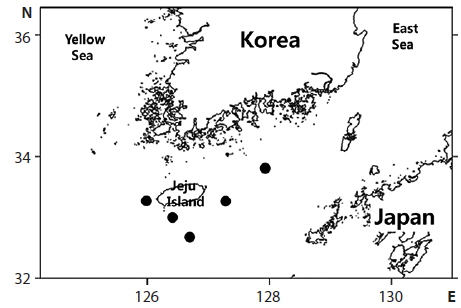
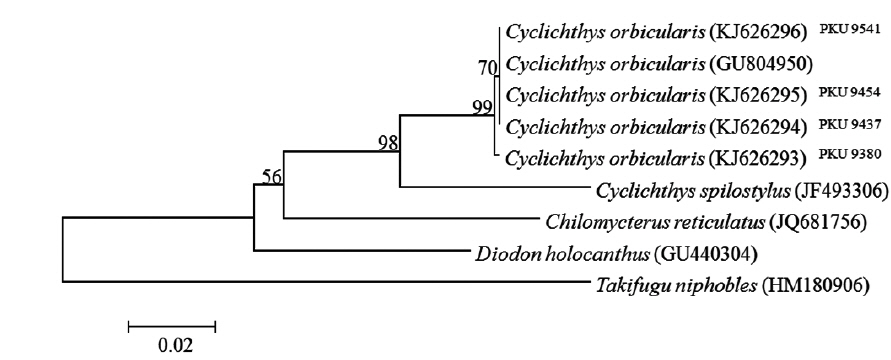
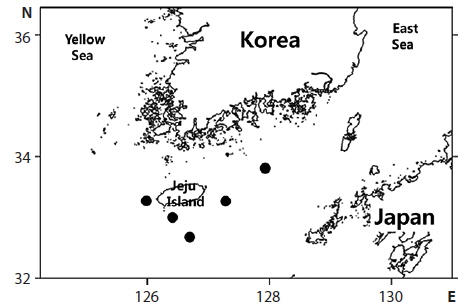
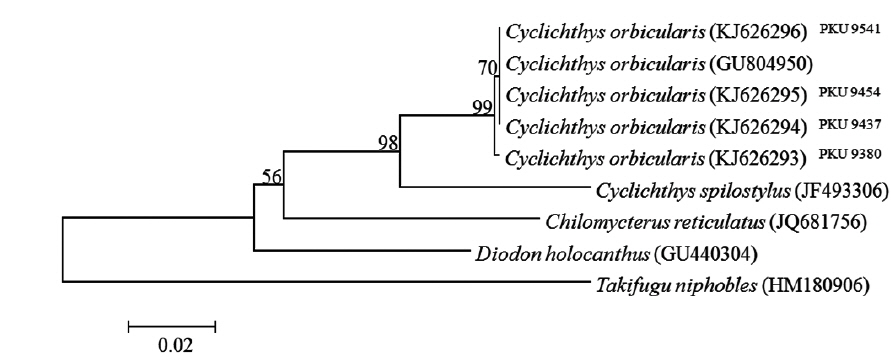
total of six specimens were collected around Jeju island, Korea between July and August 2013 (Fig. 1). Counts and measurements methods followed Nakabo (2002) with a Vernier caliper to the nearest 0.1 mm. Those specimens are deposited in the ichthyology laboratory in Pukyong National University (PKU), Korea. Genomic DNA was extracted from muscle tissue using Chelex 100 resin (Bio-rad, USA), and then polymerase chain reaction (PCR) was performed using VF2 (5´-TCAACCAACCACAAAGACATTGGCAC-3´) and FishR1 (5´-TACAC TTCTGGGTGGCCAAAGAATCA-3´) primers that amplifies mtDNA COI (Ward et al., 2005; Ivanova et al., 2007). PCR was performed in a total volume of 30 μL which contained DNA template 1 μL, dNTP 2.4 μL, 10X buffer 3 μL, Taq polymerase 0.1 μL, reverse primer 1 μL, forward primer 1 μL, and distilled water, and conducted under the following conditions: initial denaturation was for 3 min at 94°C, followed by 35 cycles of 30 s at 94°C for denaturation, 30 s at 50°C for annealing, and 90 s at 72°C for extension, with a final extension at 72°C for 10 min. The PCR products were purified with DavinchTM PCR Purification Kit (Davinch-K, Seoul, Korea). The DNA was sequenced using an ABI 3730XL sequencer and an ABI PRISMR BigDyeTM Terminator v 3.0 Ready Reaction Cycle Sequencing Kit (Applied Biosystems, USA). The nucleotide sequence was deposited in the DDBJ/EMBL/GenBank databases (accession numbers: KJ626293‒KJ626296). The sequence was aligned with ClustalW (Thompson et al., 1994) in BioEdit version 7 (Hall, 1999). Sequences of four diodontid fishes (Cyclichthys orbicularis, C. spilostylus, Chilomycterus reticulatus, Diodon holocanthus), from the National Center for Biological Information database, were used for the comparison, and Takifugu niphobles was used as an outgroup. The genetic distances were calculated with the Kimura-2-parameter model (Kimura, 1980) in MEGA 5 (Tamura et al., 2011). An neighbor-joining(NJ) tree was constructed in MEGA 5 (Tamura et al., 2011) with 5,000 bootstrap replications.
(New Korean genus name: Gud-ga-si-bok-sok)
Cyclichthys Kaup, 1855: 231, Type by subsequent designation; Bleeker, 1865: 48, Type apparently designated first; Matsuura & Sakai, 1993: 372; Leis, 2001: 3959.
Spines of body and head with 3 bases, except in C. spilostylus which has some spines on top of head with 4 bases; no spines on caudal peduncle; 9 caudal fin rays; no spots on fins. The most similar genus, Chilomycterus, is distinguishable from Cyclichthys in having a small spine on dorsal side of caudal peduncle, 10 caudal fin rays, and spots on fins (Leis, 1986).
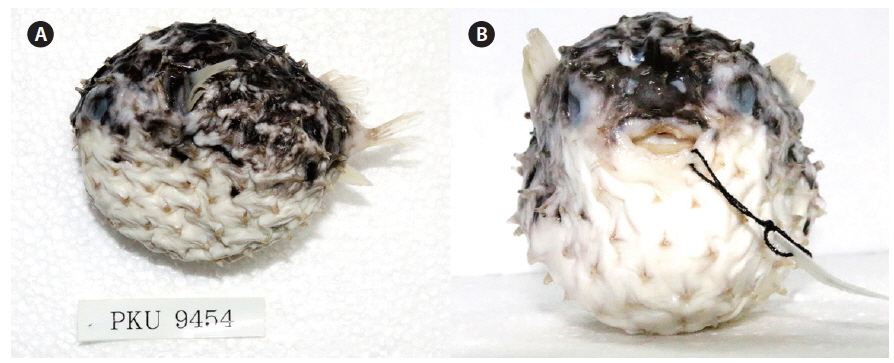
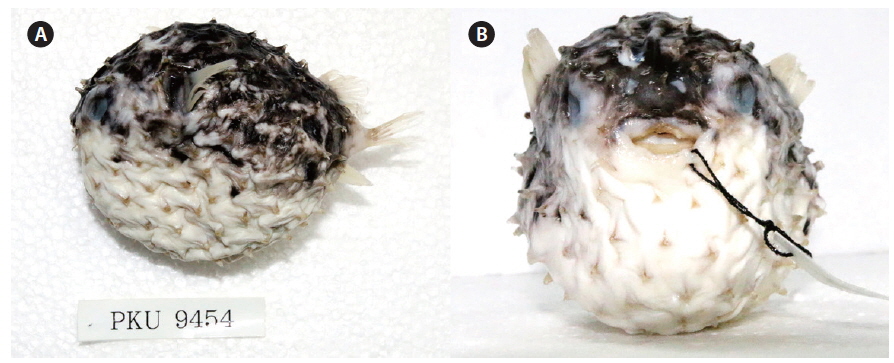
Cyclichthys orbicularis (Bloch, 1785) (Fig. 2)
(New Korean species name: Dung-geun-gud-ga-si-bok)
Diodon orbicularis Bloch 1785: 73 (Type locality: Jamaica); Allen & Swainston, 1988: 156, Australia; Matsuura & Sakai, 1993: 372, Japan; Mishra & Krishnam, 2003: 51, India.
Chilomycterus parcomaculatus von Bonde, 1923: 38 (Type locality: Union of South Africa).
Cyclichthys orbicularis: Leis, 1986: 904; Francis, 1993: 168; Matsuura et al., 1993: 372; Leis, 2001: 3962; Aizawa, 2002: 1434; Allen & Adrim, 2003: 65; Adrim et al., 2004: 127; Heemstra & Heemstra, 2004: 455; Allen et al., 2005: 420; Leis, 2006: 82; Allen and Erdmann, 2012: 1098.
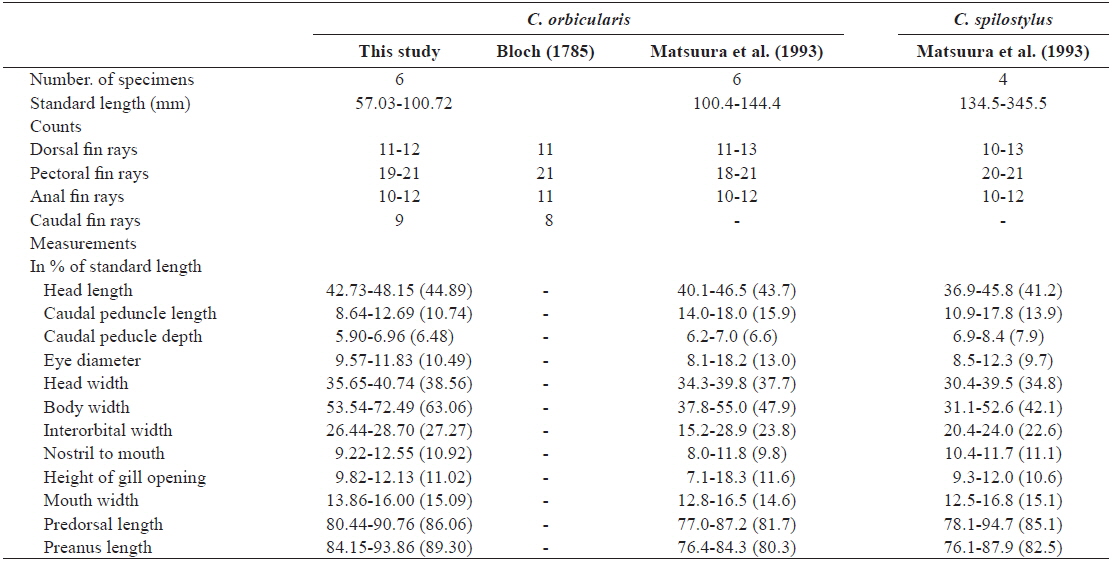
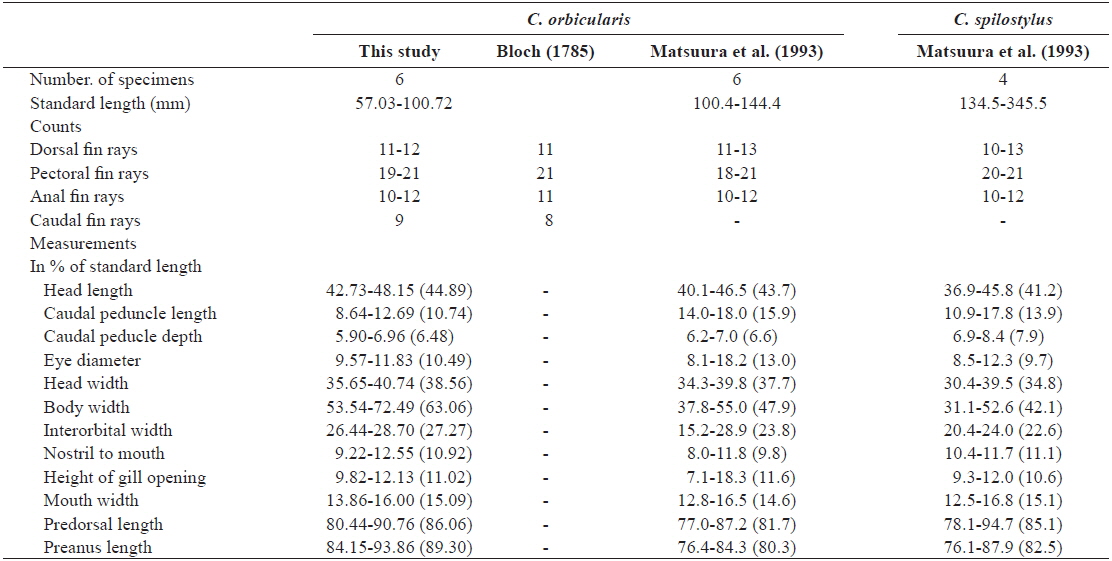
D. 11-12; P. 19-21; A. 10-11; C. 9. Meristic and morphometric characters are shown in Table 1. Body round in cross section and inflatable. Body covered with short spines; spines immovable, fixed by 3-rooted bases. Three spines dorsal to eye, 4 dorsally between pectoral fin bases, 8-9 anterior to dorsal fin base; caudal peduncle without spines. Mouth small, terminal. Posterior margin of lips not reaching to anterior margin of eye. Teeth fused to a strong and beak-like structure. Nostril a short tube with 2 openings. Gill opening a vertical slit immediately before pectoral fin bases. Pectoral fins broad and anterior to the middle of the body. Dorsal and anal fins located posteriorly on body. Pelvic fins absent; caudal fin slightly rounded. All fin rays branched.
Color when fresh. the body greyish dorsally and white ventrally. Several clusters of black spots on dorsal and lateral sides of body, none ventrally. Black spots either absent from fins or present only faintly. Coloration similar in preservation.
Found in the tropic waters of the Indo-West Pacific (Leis, 2001) and Southeast Atlantic (Leis,1986) Oceans, near reef areas, at depths of 9-170 m (Sommer et al., 1996).
Comparison of the 601 bp of mtDNA COI sequences of four specimens with those of other diodontid fishes revealed that the sequences of the specimens aligned best with that of C. orbicularis (d = 0.000-0.002), but clearly differed from those of C. spilostylus (d = 0.066-0.068), Chilomycterus reticulatus (d = 0.105-0.108), and Diodon holocanthus (d = 0.105-0.107). In an NJ tree, the specimens clustered with C. orbicularis (GU804950), with a high bootstrap value of 99% (Fig. 3).
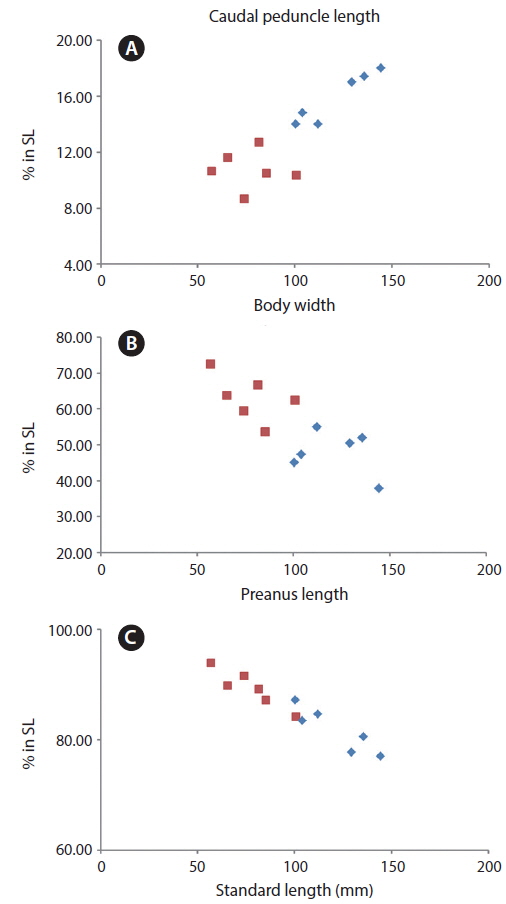
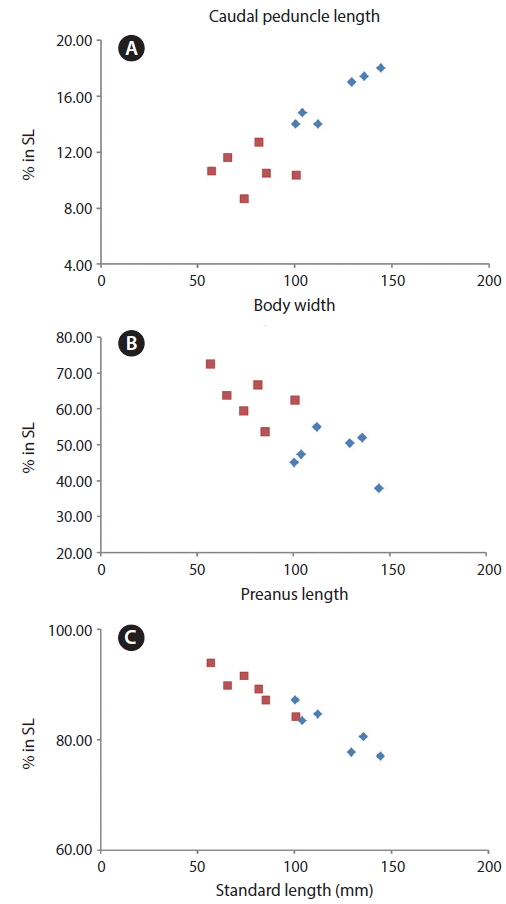
Our specimens collected around Jeju Island are placed in the genus Cyclichthys because they possess immovable spines with 3-rooted bases, 9 caudal fin rays, no spots on the fins, and no spines on the caudal peduncle (Leis, 1986, 2001). The specimens also matched the original description of C. orbicularis in most meristic counts, the presence of black spots in clusters dorsally and dorsolaterally, and three spines on top of eyes (Bloch, 1785; Leis, 2006; Allen and Erdmann, 2012). Molecular analysis supported the identification as C. orbicularis (d = 0.000-0.002). Cyclichthys orbicularis differs from Chilomycterus reticulatus in the number of caudal fin rays (9 in Cyclichthys orbicularis vs. 10 in Chilomycterus reticulatus), the spine on dorsal surface of caudal peduncle (absent vs. present), and black spots on all fins (absent vs. present) (Leis, 1986; Aizawa, 2002; Kim et al., 2005). Although, according to Matsuura et al. (1993), several measurements of Cyclichthys orbicularis overlap with those of the morphologically similar C. spilostylus, the former is well distinguished from the latter by the number of subdermal roots (all spines with 3 roots in C. orbicularis vs. some spines with 4 roots in C. spilostylus), and clusters of dark spots on ventral side of body (absent vs. present) (Leis, 2001; Aizawa, 2002). Most measurements of the specimens in this study overlapped those of C. orbicularis in Japan, but the specimens showed significant differences in body width, caudal peduncle length, and preanus length (Table 1). Generally, body shape in fishes is associated with a variety of environmental factors, type of food or feeding mode, and sexual differences as well as growth period (Marcil et al., 2006; Frederich et al., 2008; Fruciano et al., 2011; Song et al., 2013). But, when standard length was regressed against the various measurements, caudal peduncle length showed positive correlation (Fig. 4a), whereas body width and preanus length were negatively correlated with standard length (Fig. 4b, 4c). Therefore, our results suggested that body shape changes with growth, not due to regional variation. We propose the new Korean name “gud-ga-si-bok-sok” for the genus Cyclichthys, and “dung-geun-gud-ga-si-bok” for C. orbicularis based on the species’ rounded shape.