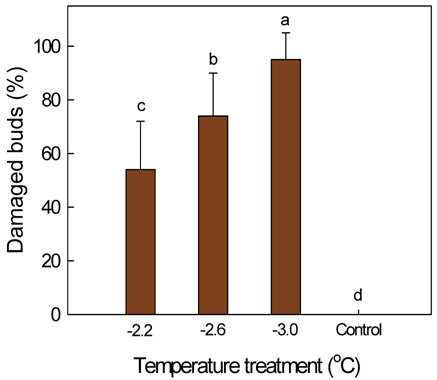
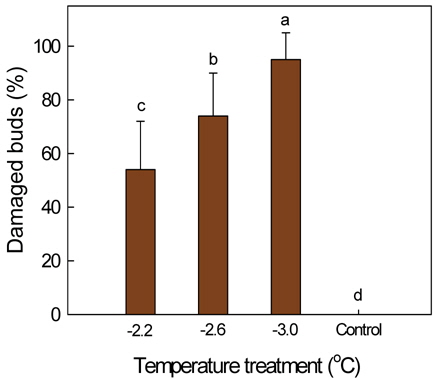
감나무의 발아기에 발생하는 서리피해 양상을 이해하기 위하여 꽃눈의 저온피해 정도에 따른 수체 생장 반응을 조사하였다. 50 L 용기에서 재배한 '부유' 나무를 4월 5일 발아기에 저온저장고에 각각 1시간 동안 -2.2±0.5, -2.6±0.5, or -3.0±0.5℃에 두어 꽃눈의 피해 정도를 달리하였고, 일부 나무는 처리를 하지 않은 대조구로 두었다. 처리온도가 -2.2℃일 때 꽃눈 피해율은 54%였으나 -3.0℃에서는 95%로 증가하여 온도가 낮을수록 꽃눈의 피해율이 유의적으로 증가하였다. 피해 꽃눈은 완전히 죽어 고사한 것, 눈의 주아가 죽고 부아가 생장하는 것, 죽지 않았으나 정상 눈에 비해 생장이 늦고 기형적인 것으로 달리 나타났다. 5월 상순 주당 꽃봉오리수는 꽃눈피해율이 높아질수록 급격히 감소하였다. 그러나 꽃눈피해가 없거나 적은 나무에서는 과다 착과가 되지 않도록 5월 중순에 꽃봉오리 솎기를 하고 7월 상순에 과실 솎기를 하였기 때문에 저온피해주의 수확기 최종 착과수는 꽃눈피해율이 60% 이내일 때 무피해주와 차이가 없었고, 수량의 경우 과중 증가로 피해율 70%까지는 뚜렷한 감소가 없었다. 꽃눈의 저온피해율이 높아짐에 따라 과중과 과피의 적색도는 유의적으로 증가하였다. 꽃눈 피해율이 높아질수록 주당 총신초장과 평균신초장 등의 영양생장이 촉진되었다. 결론적으로 발아기 꽃눈의 저온피해에 따른 수량과 영양 생장 반응은 꽃봉오리수 및 착과 정도에 의해 달라질 것으로 판단되었다.
The buds of persimmon trees are susceptible to cold damage, often with the late frost, at the time of budburst. This study was conducted to determine effect of the cold damage on shoot and fruit growth the current season.
‘Fuyu’ trees, grown in 50-L pots, were placed for 1 h at -2.2±0.5, -2.6±0.5, or -3.0±0.5℃ within a cold storage, at their budburst on April 5. Some trees under ambient temperature at 10-17℃ served as the control. Cold damage of the buds containing flower buds was 54% at -2.2℃, and significantly increased to 95% at -3.0℃. The bud damage included the complete death of all, complete death of main buds only, or the late and deformed shoot growth in the spring. Number of flower buds in early May dramatically decreased as the damage ratio increased. Since the thinning of flower buds in mid-May and fruitlets in early July was done in no or slightly damaged trees, the final number of fruits and yield did not decrease compared with the control when the damage increased by 60% and 70%, respectively. Average fruit weight and skin coloration tended to be better with increasing bud damage. Shoot growth was more vigorous in those trees whose buds were severely damaged by low temperature.
Shoot growth and the yield may depend on the number of flower buds and percent fruit set after the cold damage.
우리나라의 단감재배 안전지대는 연평균기온이 13℃ 이상이고 겨울 온도가 -12℃ 이하로 낮아지지 않는 지역으로 알려져 있다(Kim
감나무의 꽃눈(bud containing flower buds)은 혼합눈 으로서 하나의 꽃눈이 발아하여 잎과 신초가 자라고 1∼5개 정도의 꽃봉오리(flower buds)를 맺는다(RDA, 2001). ‘부 유’의 경우 결과모지의 선단 눈에서 4번째까지 꽃눈이 주로 분포하며, 그 아래는 신초만 자라는 잎눈 또는 잠아가 위치한 다(Harada, 1984; George
저온피해로 꽃눈이 일부 죽더라도 착과수가 부족하지 않을 수도 있지만, 꽃눈이 죽었을 경우에는 그 부위에 신초가 발생하지 않기 때문에 수체 생장 양상이 꽃봉오리 솎기 효과와 다를 것으로 예상된다. 저온피해에 합리적으로 대응하기 위해서는 이러한 수체 생장에 이해가 필수적이지만, 감나무의 발아기 저온피해에 따른 결실, 과실 및 영양생장 반응에 대해 구체적으로 연구한 결과는 없다. 최근 국가기관에서도 기상재해에 대비한 대응책 마련을 위해 이에 대한 기초연구 결과를 요구하고 있다. 본 연구는 감나무 유목에 저온피해를 인위적으로 유발하여 꽃눈의 피해 정도에 따른 수체 생장 반응을 파악하고자 수행되었다.
경남 김해시 진영읍에 위치한 단감연구소에서 50 L 용기에 재배한 5년생 ‘부유'(
처리를 마친 나무는 시험포장에 1.5×1 m 간격으로 완전임의로 배치하였다. 개화 전 무처리 나무 및 저온피해가 적었던 나무에서는 과다 착과를 막기 위하여 5월 16일에 꽃봉오리를 솎아주었다. 꽃봉오리 솎기 기준은 결과지당 1개를 남기고 솎는 것이 관행(RDA, 2001)이나 저온피해를 받은 나무는 결과모지당 결과지 개수가 고르지 않아 결과모지 기준으로 꽃봉오리가 6개 이상 되지 않게 솎아주었다. 생리적 낙과가 끝난 후 7월 9일에는 엽과비(엽수/과실수)가 15 이하로 낮아지지 않도록 과실을 솎아주었다. 4월부터 10월 수확 때까지 주기적으로 자동 관수장치로 물을 공급하였는데, 수분스트레스를 받기 쉬운 용기재배임을 감안하여 매일 관수하되 전엽기에 하루 평균 주당 1 L에서 개화기 이후에는 6 L 정도로 늘였다. 기비로서 4월에 퇴비를 주당 2 kg 씩 시용하였고, 5월 하순부터 9월 하순까지 4회에 걸쳐 단감용 복합비료, 요소, 염화칼륨을 시용하여 주당 N 25.5 g, P 9 g, K 21 g을 공급하였다. 생육기 동안 시험수에 양분의 과다 또는 부족 증상이 나타나지 않았고 병해충 발생도 미미하였다.
저온처리에 따른 꽃눈피해율은 처리온도가 -2.2℃일 때 54%였으나 -3.0℃에서는 95%로 증가하여 처리온도가 낮을수록 꽃눈의 피해율이 유의적으로 증가하였는데(Fig. 2), 본연구의 목적에 부합되도록 총 20주의 꽃눈피해율을 기준으로 각각 수체 반응을 조사하여 결과를 정리하였다. 저온피해주의 꽃눈피해율은 무피해 나무들의 평균 꽃눈수를 기준하여 산출하되, 발아 당시의 눈 형태로 꽃눈 파악이 어렵기 때문에, 5월 2일에 쉽게 구분이 되는 결과지(꽃눈에서 발생하여 꽃봉오리를 맺은 가지)수를 세어 기준 꽃눈수를 정하였고, 기준 꽃눈수에서 저온피해주의 남아있는 결과지수를 뺀 것을 피해 꽃눈수로 간주하였다. 저온피해에 따른 꽃눈의 생장 상태를 4월 23일과 5월 17일에 관찰하였으며, 적뢰를 하기 전인 5월 2일 주당 꽃봉오리수를 조사하였고, 솎기 후 남은 꽃봉오리수를 기준으로 7월 6일까지의 낙과율을 구하였다. 10월 31일에 과실 전체를 수확하여 착과수, 수량, 평균과중을 조사한 다음, 무피해 또는 피해가 적은 나무는 주당 15개, 피해가 심한 나무는 과실 전부를 대상으로 과피색, 경도, 당도를 측정하였다. 과피색은 휴대용색도계(CM-2500d, Konica Minolta Sensing Inc., Japan)를 사용하여 적색도 Hunter a 값을 구하였고, 과육의 경도는 직경 5 mm 탐침을 사용하여 경도계로 측정하였다. 당도는 과실의 적도부에서 과피로부터 약 2 cm 깊이까지 착즙하여 굴절당도계(N1, Atago Co., Japan)로 측정하였다. 11월 9일 나무의 신초수와 총신초장을 조사하여 평균신초장을 구하였으며, 주당 20개의 잎을 신초의 중간 부위에서 채취하여 엽면적을 측정하였다. 온도처리 수준에 따른 꽃눈피해율을 최소유의차 검정으로 통계분석을 하였고, 꽃눈피해율에 따른 수체 반응 성적은 Sigmaplot(Ver. 8.0, SPSS Inc., Ill., USA) 프로그램을 이용하여 회귀분석을 하였다.
Fig. 3은 저온피해를 받은 후 18일이 지난 4월 23일의 꽃눈 생장 상태를 나타낸 것이다. 피해 꽃눈은 완전히 죽어 고사한 것(Fig. 3 B), 주아가 죽고 부아가 생장하는 것(Fig. 3 C), 정상 눈에 비해 생장이 늦고 기형적인 것 등으로 달리 나타났다(Fig. 3 D). 눈 피해가 심한 나무의 가지 수피를 벗겨본 결과, 수피와 목질부까지 변색되지는 않은 것으로 나타났다. Fig. 4는 저온피해 나무의 5월 17일 신초 생장으로, 가지 선단의 눈이 죽은 경우에는 아래쪽 눈의 생장이 왕성하였으며 (Fig. 4 A) 죽은 눈이 많을수록 꽃봉오리를 맺는 신초는 적어졌다. 또한 부아에서 생장한 신초는 꽃봉오리를 맺지 못하고 영양생장만 계속하였다(Fig. 4 B). 눈의 조직 일부가 죽어 기형적으로 생장하는 눈은 자라다가 고사하거나 신초 생장이 빨리 멎어 꽃봉오리가 맺히더라도 정상적인 생장이 어려운 것도 있었다(Fig. 4 C).
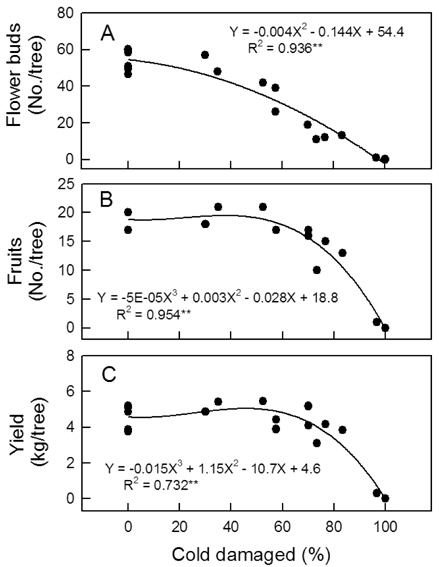
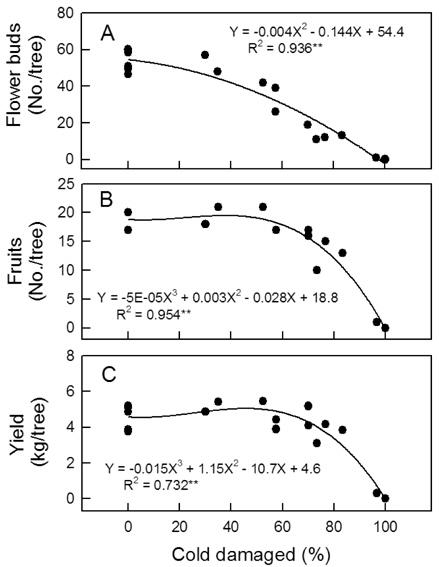
5월 2일 주당 꽃봉오리수는 무피해주에서 45-60개였으나 꽃눈피해율이 높아질수록 급격하게 감소하여 100% 피해를 받은 나무에서는 하나도 없었다(Fig. 5 A). 생리적 낙과가 끝난 후 7월 6일까지 시험수의 낙과율은 3-10% 정도로 낮았고 꽃눈피해율에 따른 차이도 없었다(자료 미제시). 저온해를 받지 않았거나 약하게 받아 꽃봉오리가 많았던 나무들은 7월에 과실 솎기를 하여 엽과비가 15 이하로 되지 않도록 하였기 때문에, 저온피해주의 수확기 최종 착과수는 꽃눈피해율이 60% 이내일 때 주당 17-21개로 무피해주와 차이가 없었다 (Fig. 5 B). 그러나 피해율이 60% 이상으로 높아지면 착과수가 급격히 낮아졌다. 수량의 경우 피해율 70%까지 뚜렷한 감소가 없었고, 73% 이상에서 감소하였다(Fig. 5 C).
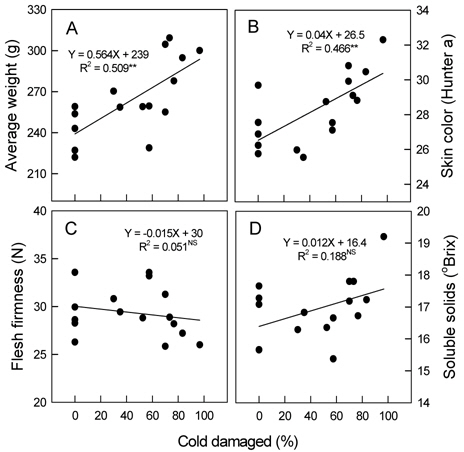
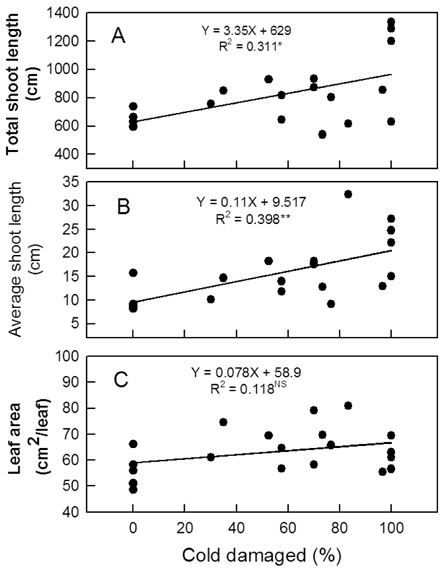
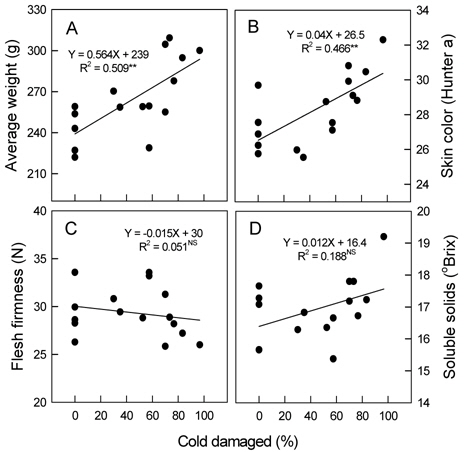
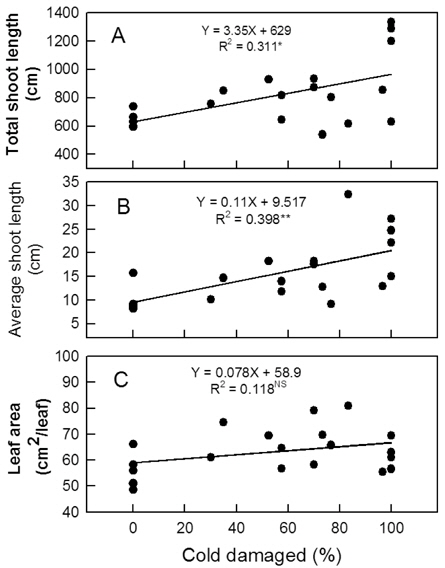
꽃눈의 저온피해율이 높아짐에 따라 평균과중과 과피의 적색도를 나타내는 Hunter a값은 유의적으로 증가하였다 (Fig. 6 A, 6 B). 평균과중의 경우 무피해주에서 222-259 g이었지만, 70% 이상 피해를 받은 나무에서는 278 g 이상으로 증가하였다. 꽃눈피해율 증가와 함께 과육의 경도가 감소하고 (Fig. 6 C) 당도는 높아지는 경향이 있었으나(Fig. 6 D) 통계적인 유의성은 인정되지 않았다. 주당 총신초장과 평균신초장은 꽃눈피해율이 높아질수록 변이의 폭이 커지는 양상이었지만 증가는 뚜렷하였다(Fig. 7 A, 7 B). 단위 엽면적도 유의적인 변화는 아니었으나 피해율 증가와 함께 커지는 경향이었다 (Fig. 7 C).
Fig. 3과 4에 나타난 저온피해 양상은 일반적으로 서리피해를 받은 나무에서 관찰되는 것과 비슷하여, 본 연구의 결과는 자연상태에서 피해를 받은 나무의 생장 반응과 유사함을 시사하는 것으로 볼 수 있다. 발아기 저온피해가 드문 지역에서는 농가가 꽃눈의 저온피해를 인식하지 못하는 사례가 적지 않다. 따라서 이러한 증상 자료는 발아기 저온피해를 진단하는데 참고가 될 수 있을 것이다. 60% 이내의 꽃눈피해율에서 꽃봉오리수 감소가 착과수 감소로 이어지지 않은 것은 피해가 없었거나 적은 나무의 꽃봉오리들을 적정 착과량 유지를 위해 개화 전에 솎았고, 낙과가 끝난 후 과실도 일부 솎았기 때문이었다. 과실 솎기를 일찍 하고 많이 할수록 단위 과실에 공급되는 양분량이 증가하므로 과실 비대는 촉진되는 것으로 알려져 있다(Palmer
착과수가 적으면 과실로 공급되는 탄수화물이 많아져 비대가 증가할 뿐만 아니라 착색이 빨라지기도 한다(Walter, 1967; Embree
종합하면 발아기에 꽃눈의 저온피해가 있더라도 목표 착과수에 필요한 꽃봉오리수가 부족하지 않고 생리적 낙과가 심하지 않다면 수량 확보에 큰 어려움이 없을 것으로 판단되었다. 반면, 꽃봉오리가 부족하여 착과량 감소가 심한 나무에서는 신초 생장이 지나치게 강해질 수 있으므로 수세 관리에 유의해야 할 것이다.