To evaluate the influence of burrowing substrate on the rearing performance and ambicoloration of cultured flounder,
넙치(
생태적 측면에서, 넙치를 포함한 가자미목 어류(sole, turbot, plaice 및 halibut 등)는 자연계에서 모래나 자갈과 같은 잠입성 기층이 존재하는 곳에서 저서 생활을 한다. 이들은 포식자들에 의한 피식 위험으로부터 자신을 보호하기 위해 서식 공간의 퇴적층에 자신의 색상을 맞추고, 몸을 파묻는 서식 습성을 지니고 있다(Ramachandran et al., 1996; Ellis et al., 1997). 그러나 바닥면이 편평하고 미끈한 경질의 합성수지 또는 콘크리트 재질의 인공 시설에서 사육은 양식산 가자미류의 잠입 행동과 습성을 강제하고, 개체 간 공식 또는 먹이 경쟁을 통한 잦은 충돌을 빚게 함으로써, 방어적 목적에서 체색발현 기전을 이용해 자신을 은폐하거나 위장하려는 기능을 강화하려는 경향을 보일 것이다. 따라서 이러한 일렬의 과정 속에서 가자미류 어류들은 표피 색소발현 장애 및 색소포 이상 과다 발현을 유도 할 수 있을 것으로 추정된다. 그러므로 만약 고밀도로 사육중인 가자미류 등에게 본능적인 잠입 행동과 습성을 발휘할 수 있게 환경 조건을 제공해 준다면, 무안측의 색소포 이상 과다 발현을 제어할 것으로 본다.
이에 본 연구에서 우리는 넙치의 무안측 흑화 현상이 이들의 잠입 행동과 습성에 어떤 관련성이 있는지 알아보기 위해, 자연계와 유사한 환경으로 수조 바닥에 소형자갈을 깔아준 뒤, 사육넙치의 무안측 체색에 변화가 일어나는 지 조사하였다.
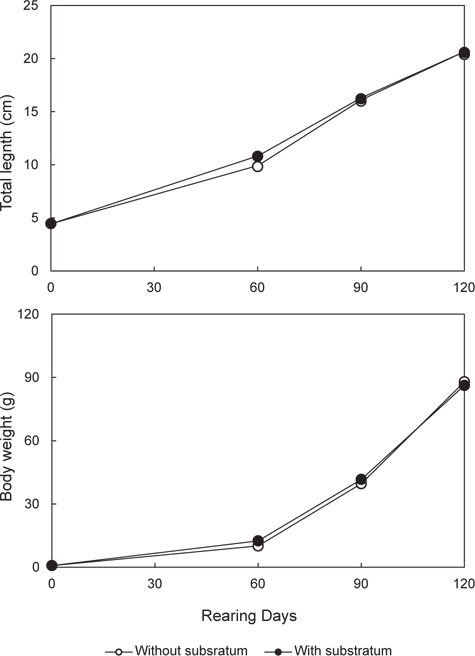
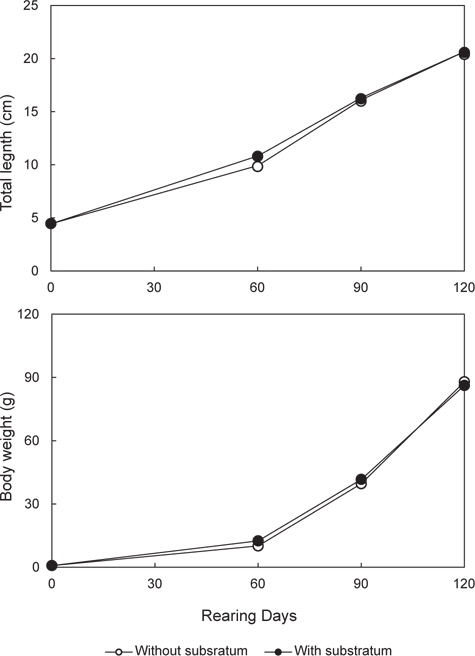
본 실험에서는 무안측 체색발현에 잠입 행동이 어떤 영향을 미치는지 알아보기 위해, 부화 후 2개월 된 넙치 치어(전장 4.46±0.06 cm, 무게 0.77±0.03 g)를 이용해 암녹색 1 ton 수조에서 2반복으로 실시하였다. 실험구는 통상적으로 양식 현장에서 사용되는 바닥면이 매끈하고 편평한 암녹색 FRP 수조를 대조구로 설정하였으며, 비교구(이하 기질구)는 잠입 행동을 발휘하게 하기 위해 수조 바닥에 소형 자갈(직경 4-6 mm)을 잠입 기층으로 제공하였다(Fig. 1). 사육실험은 성장기인 봄에서부터 여름까지 120일간 실시하였다 .
본 연구에서 사육수는 실험구별로 동일하게 1일 약 24회전이 되도록 해주었으며, 수온 및 염분 등 환경 인자들 자연 상태로 맞춰주었다. 환경 인자들은 매일 측정하였으며, 이때 평균 수온은 19.9±4.2℃, pH 7.9±0.7, DO 7.7±0.1 mg/L 및 염분은 30.4±1.1 psu이었다. 실험 기간 중 공급된 먹이는 입자 직경 1.1-6.2 mm의 상업용 EP부상사료(광어 1-6호, 천하제일, KOREA)을 이용하였으며, 먹이는 2회/일(오전10시와 오후4시)로 사육어가 포만 상태에 이를 때까지 제공되었다.
DFI, FE, 생존율 및 성장
매일 2번의 먹이 공급 후 허실 분을 제외한 일간 총섭식량을 기록하고, 그 수치를 바탕으로 아래의 공식으로 일간 개체당 사료섭식량(daily food intake: DFI, mg/fish/day = 소비된 사료중량/[사육일수×수용마리수]) 및 사료효율(feed efficiency: FE, % = [어체의 습중량 증가분/건조 사료 섭취량]×100)을 구하였다. 한편 개시일, 60일, 90일 및 120일에 버니어캘리퍼스를 이용하여 전장은 1 mm 단위까지, 전자저울을 사용하여 체중은 0.01 g까지 측정하였다. 또한 매일 폐사 개체를 수집하여 계수한 뒤 생존율을 산정하였다.
무안측 흑화률
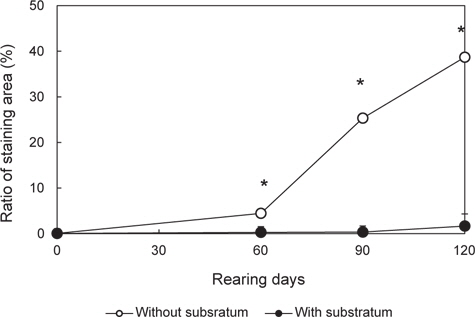
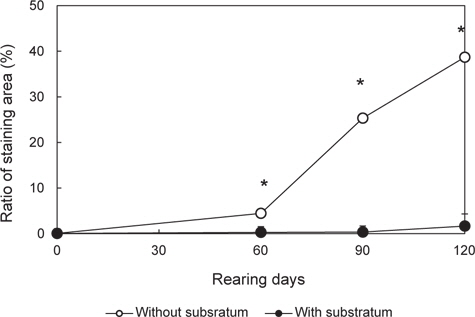
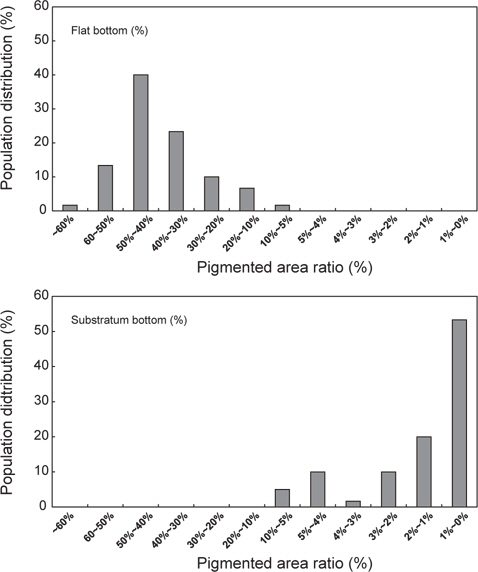
실험구별 무안측 흑화 발현 경향을 알아보기 위해, 실험 0일째 무안측 착색 면적률(0.04±0.01%)를 제시하고, 실험 개시 후 60일, 90일과 120일에 두 실험구의 면적비를 조사하였다. 이를 위해 해당 기간에 실험구별 30마리 넙치를 포획하여, 마취한 뒤, 성장도 측정 후 디지털카메라로 무안측 형태를 실험구별로 촬영을 하였다. 이 영상을 이용해 개체당 무안측 착색면적을 영상분석시스템(LEICA co., Germany)에서 구하였다. 이 값을 바탕으로 개체당 착색 면적률(%=[착색 면적/무안측 면적]×100)을 구하고, 착색 면적비가 1%를 넘어선 개체 수를 계수하여 실험구별 착색 개체비(% = [착색 개체 수/총샘플 수]×100)을 산정하였다. 또한 이 값을 이용해, 실험 종료(120일) 시 그룹별 착색면적의 군집비율을 구하였다. 이를 위해 착색면적률 구간을 60%이상, 60-50%, 50-40%, 40-30%, 30-20%, 20-10%, 10-5%, 5-4%, 4-3%, 3-2%, 2-1% 및 1-0%로 구획화하였고, 해당 범위 비율에 포함되는 개체 수를 계수한 후, 군집 내 비율로 환산하였다.
통계처리
실험결과 데이터들의 통계 처리는 실험구별 평균값의 유의차 유무를 가리기 위해 SPSS 7.0 통계프로그램을 이용하여 사료효율, 사료섭식량 및 생존율은 비모수적 방법인 Kruskall-Wallis test를 통해 검증하였고, 성장과 흑화률은 모수적 방법인 Student t-test를 통해 그룹별 평균 값의 차이 유무를 확인하였다. 이때 사용한 유의 수준은 5%로 하였다.
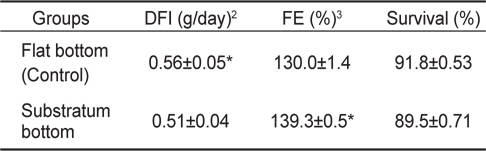
실험구별 DFI 경우 잠입기질이 없는 대조구가 0.56±0.05 g/day으로 기질구 0.51±0.04 g/day보다 유의하게 높은 값을 보였다(
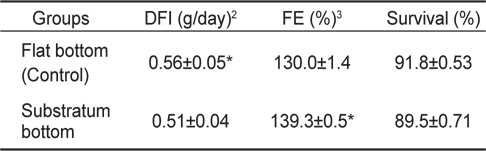
Daily food intake (DFI), food efficiency (FE) and survival rates of fry olive flounders reared in flat-bottom and substratum-bottom aquariums for 120 days1
개시 시, 모든 실험구가 동일한 착색면적률(0.04±0.05%)은 실험 60일에 이르러 대조구(4.44±0.47%)가 유의하게 상승하여 기질구(0.27±0.01%)에 비해 높은 값을 나타내기 시작하였다(
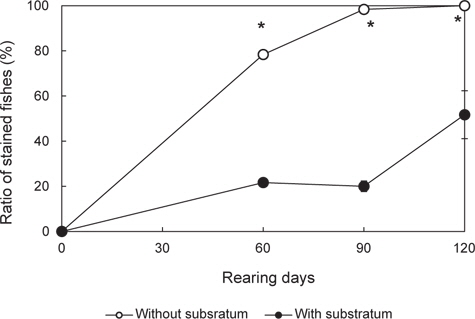
실험구별 무안측 흑화 개체비는 Fig. 4와 같다. 60일에 이르러 대조구 78.4±1.2% 및 기질구 21.7±1.2%로, 두 실험구가 이전에 비해 흑화 개체수가 유의하게 늘어난 것을 확인할 수 있었다. 또한 실험구간 비교를 통해 대조구의 증가 속도가 기질구보다 유의하게 빠르다는 사실을 알 수 있다(
이러한 실험구별 무안측의 착색 경향을 형태학적으로 비교해 보면 Fig. 5와 같다. 그림을 통해 알 수 있듯이, 대조구는 다수의 개체들이 무안측의 표피 전반에 걸쳐 체색이 입혀져 있지만, 기질구는 대부분이 흰색의 비착색 형태를 나타내고 있다. 이것을 분포비율로 비교해보면, 대조구인 경우 가장 낮은 착색면적률은 4.95%로, 대부분(약 60%)의 개체가 착색률 30-50%를 나타내고 있으며, 그 중에서도 가장 많은 개체수가 50-40% 착색률을 보였다. 그에 반해 바닥에 자갈을 깔아 준 기질구의 경우, 10%를 상회하는 개체는 없었으며, 50% 이상의 개체가 0-1%의 착색면적률을 나타내었다(Fig. 6).
본 연구에서는 고밀도 사육 시 넙치의 식욕, 성장, 생존 및 무안측 흑화 발현에 있어 잠입 기질 제공에 따른 영향을 알아보았다. 우선 사육실험 결과, 비록 성장과 생존에는 실험구간 유의 차가 없었지만, 기질구는 대조구에 비해 낮은 DFI 값과 높은 FE 값은 나타내었다. 이는 편평한 바닥 면의 사육수조를 이용한 통상적이 양식기법보다 바닥에 잠입기질을 제공하는 것이 넙치의 단위 생산성을 높일 수 있음을 의미한다. 이러한 잠입 기질의 효과는 자연계에서 서식하는 넙치의 본능적인 습성이 영위될 수 있게 환경조건을 제공해 줄 경우 이들의 성장에 긍정적인 영향을 미칠 수 있음을 보여주고 있다. 자연계에 서식하는 넙치는 해수면 저층에서 단독 생활을 영위하며, 모래 또는 자갈과 같은 잠입성 기질을 이용해 자신의 몸을 숨겨 포식자로 하여금 자신을 보호하고, 먹이 사냥 시에는 위장을 통해 먹이감을 쉽게 포획하는 생활을 영위한다. 그러나 양식 넙치의 경우 집단 사육에 의한 군집생활로 인해 서식공간 제한, 잠입습성 제약, 동족간 먹이경쟁과 신체접촉 등으로 인해 자신의 습성과 다른 생활환경으로 인한 물리적 스트레스를 만성적으로 경험하게 된다. 따라서 이럴 경우 어류는 스트레스성 대사과정을 극복하기 위해 많은 에너지를 요구하게 되고, 이것이 높은 사료섭식량과 낮은 사료효율로 나타날 수 있다. 실제 어류에 있어 만성 스트레스가 대사에너지의 소비를 통해 어류 성장을 저해할 수 있다는 사실은 다수의 연구(Farbridge and Leatherland, 1992; Foo and Lam, 1993; McCormick et al., 1998; Papoutsoglou et al., 2000)를 통해 확인할 수 있다. 특히 스트레스 관련 호르몬인 코티졸은 경골어류의 먹이섭식량 조절(Bernier et al., 2004), 성장호르몬과 Prolactin (Uchida et al., 2004), 및 neuropeptide Y과 corticotropin-releasing factor (Bernier et al., 2004)과 같은 호르몬 분비 활성을 조절함으로써 대사에 관여하는 것은 알려진 사실이다. 그러므로 대조구와 기질구 사이 성장 차가 없음에도 불구하고 기질구가 낮은 DFI과 높은 FE을 보인 것은, 스트레스 관련성을 배제하고라도, 기질구 개체들이 잠입생활로 인한 소비되는 에너지 량이 대조구에 비해 상대적으로 낮음으로 나타난 결과 추정된다.
한편 잠입기질은, 고밀도 사육에서 성장에 의한 전체 생육밀도(biomass)가 급격히 증가하여도, 무안측 체색발현을 효과적으로 억제한다는 것을 확인할 수 있었다. 즉, 총 120일 간의 실험 종료 후 군집 내 흑화 개체 수를 조사해 본 결과, 암녹색 편평한 FRP수조(대조구)는 모든 개체가 기형적인 착색이 이루어진 반면, 잠입 기질구는 약 47%만이 기형적인 착색현상을 나타내었다. 흑화 면적률 역시, 대조구는 가장 많은 비율로 40-50%의 착색면적률을 보였고, 모든 개체가 5%의 착색률를 나타내었지만, 기질구는 53%가 1% 이하의 착색면적률을 보인 정상개체였다. 착색 개체들의 경우는 대부분 10% 이하의 착색면적률을 보였다. 이러한 결과는 고밀도 사육 시 잠입 행동과 습성의 강제가 넙치의 비정상적인 무안측 체색 발현에 아주 큰 영향을 미치며, 더불어 바닥에 자갈과 같은 잠입기질을 제공할 경우 이를 효과적으로 예방할 수 있음을 의미한다. 이러한 결과는 수조 바닥 면에 잠입성 기질로 모래를 제공한 후 저밀도에서 실험을 실시한 Ottesen and Strand (1996), Iwata and Kikuchi (1998) 및 Estevez et al. (2001)의 연구에서도 확인된다. 따라서 육상 수조식 양식장에서 고밀도 사육 시 발현되는 가자미류의 무안측 흑화(Kang et al., 2011)는 수조바닥 면에 잠입 기층 제공을 통해 효율적으로 예방할 수 있을 것으로 사료된다. 또한 사육 관리상 배설물과 찌거기 제거의 어려움으로 지수상태에서 저밀도로 실험한 이전 연구자들의 결과와 달리, 본 연구에서는 산업현장과 동일한 고밀도 및 유수 조건에서 취급과 수질 관리가 모래보다 용이한 자갈을 잠입 기질로 사용한 점에서 실제 산업현장에서 적용 가능성은 높다. 다시 말해 모래는 특성상 유속에 의한 유실 가능성이 높으며, 배설물과 사료찌꺼기 등이 섞여 수질 안정화에 어려움에 있고, 그로 인한 질병 발병률이 높아, 실제 산업현장에서 사용은 어려움이 많다. 그러나 본 연구에 사용된 소형 자갈(직경 4-6 mm) 경우 물리적 특성상, 기술적으로 모래의 단점을 보완하였기에 수질관리와 질병 발생의 위험은 낮다. 하지만, 이 방법의 경우, 역시 유수식 사육 환경에서 중력 이상의 강한 유속이 발생할 경우, 자갈 입자가 한 방향으로 쏠리기 때문에 일정 주기로 이 저면기질을 고르게 펼쳐줘야 하는 번거로움이 있다.
이상의 연구결과, 고밀도 넙치 사육 시 수조 내 잠입기질 제공은 성장도에는 큰 영향을 미치지 않지만, 높은 사료 이용효율로 생산성이 높일 수 있으며, 더불어 비정상적인 무안측 색소포 분화 및 발현을 효과적으로 억제한다는 사실을 확인하였다. 그러나 이 잠입기질의 효과가 사육 넙치에서 잠입 행동을 구현할 수 있게 해 나타난 결과인지, 아니면 무안측 표피에 대한 입자의 물리적 자극이 이러한 효과를 만들어 냈는지에 대한 답은 찾지 못했다. 따라서 무안측 흑화 발현 억제에 있어 잠입기질의 작용 기전을 이해하기 위해, 향후 수조 저면의 표면 굴곡이 가자미류의 무안측 착색현상에 미치는 효과를 연구해 볼 필요가 있다.