Excessive metals in the soil have become one of the most significant environmental problems. Phytoremediation has received considerable attention as a method for restoring the contaminated soils. The microbes having remarkable metal tolerance and plant growth-promoting abilities could also play a significant role in remediation of metal-contaminated soils, because bioaugmentation with such microbes could promote phytoextraction of metals. Therefore, the present study was focused on evaluating the phytoextraction of heavy metals (Co, Pb and Zn) in Helianthus annuus (sunflower) induced by bioaugmentation of a phosphate solubilizing bacterium.
A phosphate solubilizing bacterium was isolated from metal-contaminated soils based on the greater halo size (>3 mm) with solid NBRIP agar medium containing 10 g glucose, 5 g Ca3(PO4)2, 5 g MgCl2⋅6H2O, 0.25 g MgSO4.7H2O, 0.2 g KCl, 0.1 g (NH4)2SO4 in 1 L distilled water. Isolated bacterial strain was assessed for their resistance to heavy metals; CoCl2.6H2O, 2PbCO3.Pb(OH)2, and ZnCl2 at various concentrations ranging from 100-400 μg/mL (Co, Pb and Zn) using the agar dilution method. A pot experiment was conducted with aqueous solutions of different heavy metals (Co, Pb and Zn) to assess the effect of bacterial strain on growth and metal uptake by Helianthus annuus (sunflower). The impact of bacterial inoculation on the mobility of metals in soil was investigated under laboratory conditions with 50 mL scaled polypropylene centrifuge tubes. The metal contents in the filtrate of plant extracts were determined using an atomic absorption spectrophotometer (Perkinelmer, Aanalyst 800, USA).
Inoculation with Enterobacter ludwigii PSB 28 resulted in increased shoot and root biomass and enhanced accumulation of Co, Pb and Zn in Helianthus annuus plants. The strain was found to be capable of promoting metal translocation from the roots to the shoots of H. annuus. Therefore, Enterobacter ludwigii PSB 28 could be identified as an effective promoter of phytoextraction of Co, Pb and Zn from metal-contaminated soils.
Metal contamination of soils has become one of the most significant environmental problems today. Metal uptake by crop plants from the contaminated agricultural lands can have strong adverse impacts on human health through the food chain (Karavoltsos
With the continuous addition of undesirable metals into the environment, remediation of contaminated soils receives increasing attention (Cao
The amount of heavy metals uptake in plants varies with the mobility and the concentration of metals in soil (Chen
>
Isolation of phosphate solubilizing bacterial strains
Heavy metal contaminated soils collected from abandoned mines of Boryeong area in South Korea were used in isolating phosphate solubilizing bacteria. Aliquots of serially diluted soil samples were inoculated on NBRIP (National Botanical Research Institute Phosphate) medium containing 10 g glucose, 5 g Ca3(PO4)2, 5 g MgCl2.6H2O, 0.25 g MgSO4.7H2O, 0.2 g KCl, 0.1 g (NH4)2SO4 in 1 L distilled water (Nautiyal, 1999). The pH of the media was adjusted to 7 ± 0.1. The petri plates were incubated at 300C for 7 days. Morphologically distinct colonies with clear halos were purified by repeated sub culturing. A total of 20 isolates were selected based on the greater halo size (>3 mm) and maintained on solid NBRIP agar medium until use.
>
Assay of heavy metal resistance
Isolated bacterial strains were assessed for their resistance to heavy metals using the agar dilution method (Cervantes
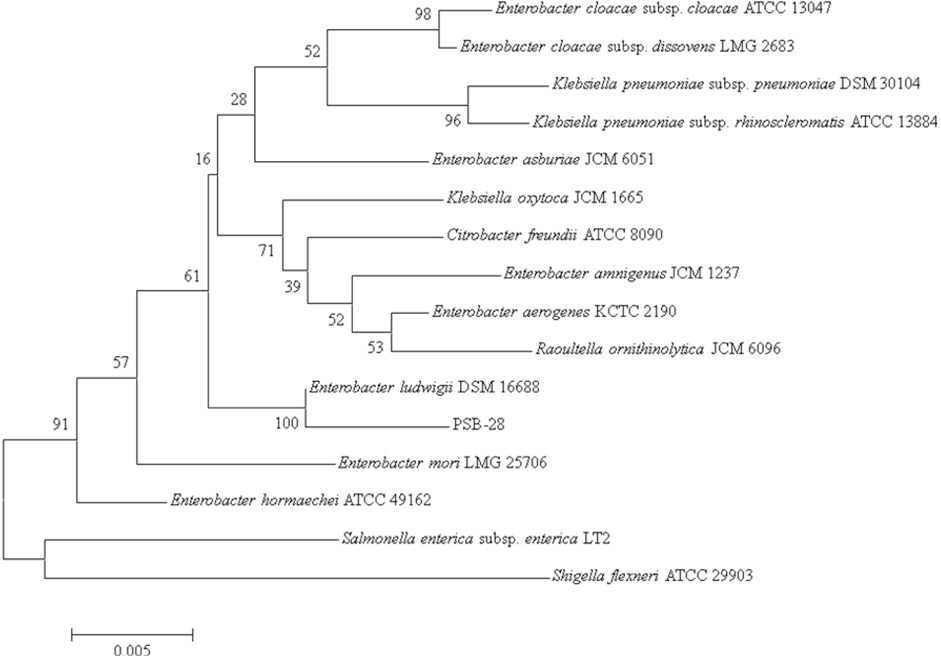
The partial sequencing of 16S rRNA for the bacterial strain was done with the help of DNA sequencing service, SOLGENT, Daejeon, South Korea using universal primers, 27F (5′-AGAGTTTGATCCTGGCTCAG -3′) and 1492R (5′-GGTTACCTTGTTACGACTT -3′). The online program BLAST was used in identifying the related sequences with known taxonomic information available at the databank of NCBI (http://www.ncbi. nlm.nih.gov/BLAST). A Phylogenetic tree was constructed using CLUSTAL X program (Thompson et al., 1997), which involved sequence alignment by neighbor joining method (Saitou and Nei, 1987) and maximum parsimony using the MEGA4 program (Kumar et al., 2001). Grouping of sequences was based on confidence values obtained by bootstrap analysis of 1,000 replicates. Gaps were edited in the BioEdit program and evolutionary distances were calculated using Kimura two parameter model. Reference sequences were retrieved from GenBank under the accession numbers indicated in the trees.
>
Effect of heavy metals on bacterial growth
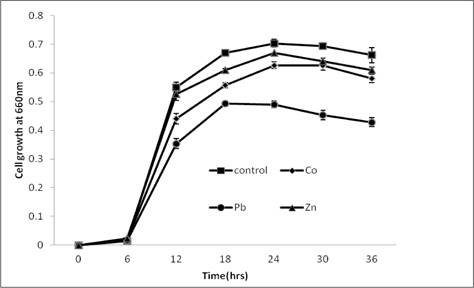
NBRIP medium supplemented with heavy metals (Co, Pb and Zn) at the concentration of 200 mg/L was inoculated with bacterial suspension (106 colony forming units/ml) and incubated with continuous shaking at 300C. Optical density of culture supernatant was measured at definite time intervals using UV spectrophotometer at 660 nm to estimate the cell growth.
>
Assay of inorganic phosphate solubilization
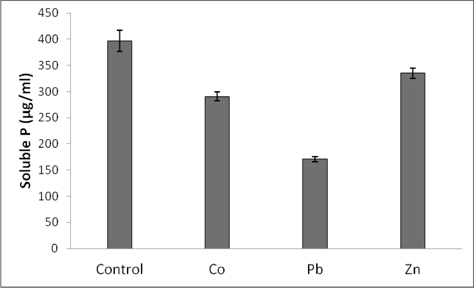
Bacterial culture having 106 colony forming units/mL (2 days old) was inoculated in sterilized liquid NBRIP medium (250 mL) supplemented with different heavy metals (Co, Pb and Zn) at the concentration of 200 mg/L and incubated with continuous shaking at 300C. Sterilized liquid NBRIP medium without supplemented with heavy metals was served as a control. A sample (10 ml) of each cultured and control were taken and centrifuged at 12000 ×g for 15 min. The clear supernatant was used in determining the pH and amount of phosphorous released into the medium.
>
Effect of bacterial strain on growth and metal uptake by H. annuus
A pot experiment was conducted under green house conditions at the College of Agriculture, Chungnam National University in October 2012. The soils collected from several locations of a nearby forest and a waste button mushroom bed in Buyeo-gun area, Chungchungnam-do, South Korea, were mixed with the ratio of 1:1, air dried and sieved (2 mm). Sterilized soil (by steaming at 100℃ for three consecutive days) was amended with aqueous solutions of different heavy metals (Co, Pb and Zn) to achieve the final concentrations of 200 mg/kg soil. They were then kept for 2 weeks in a greenhouse for metal stabilization and used in filling the plastic pots (25 cm diameter, 35 cm height). Seeds of
>
Mobility of the metals in soil
The impact of bacterial inoculation on the mobility of metals in soil was investigated under laboratory conditions with 50 mL scaled polypropylene centrifuge tubes. The bacterial strain transferred into 100 mL flasks containing LB broth was cultured aerobically on a rotating shaker (150 rpm) at 30℃ until reaching the final concentration of 106 colony forming units/mL. The bacterial cells were then harvested by centrifugation at 10000 × g for 15 min and washed in phosphate buffer (pH 7.0) twice. The bacterial pellet was washed in sterile water, re-centrifuged, and finally re-suspended in 5 mL sterile water. Artificially contaminated soil (1 g) in the centrifuge tubes was inoculated with small aliquots (up to 1 mL) of the final washed bacterial culture. After taking the weight the tubes, they were wrapped with brown paper and placed on an orbital shaker at 200 rpm at 25 ℃. At the end of the period of 10 d, the weight of the tubes was recorded and 10 mL of sterile water were added to each tube to extract the soil water soluble heavy metals. The extracts were centrifuged at 10000 × g for 10 min and filtered and the metal contents in the filtrate were determined using an atomic absorption spectrophotometer (Perkinelmer, Aanalyst 800, USA). Artificially contaminated soil without inoculation with the strain served as the control after centrifugation.
>
Isolation and identification of phosphate solubilizing bacterial strain
A total of 20 bacterial strains with the potential ability to solubilize inorganic phosphates were screened based on the greater halo size (>3 mm). They were then assessed for their resistance to heavy metals and the bacterial strain showing the highest degree of metal resistance was selected for the study. According to 16S rRNA sequence analysis, the selected strain showed close proximity with
>
Effect of heavy metals on bacterial growth
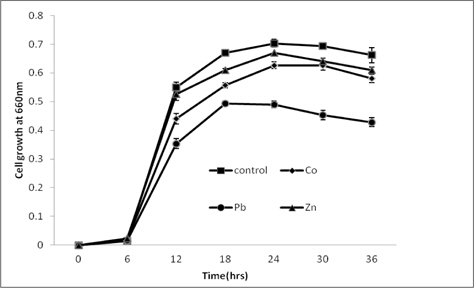
The growth of the strain measured as the optical density of culture supernatant at definite time intervals is given in Fig. 2. During the incubation period of 36 h, none of the metal was found to be highly toxic to the strain. However, compared to the metal free culture medium, slight reductions in bacterial growth were observed in metal supplemented media.
>
Inorganic phosphate solubilization
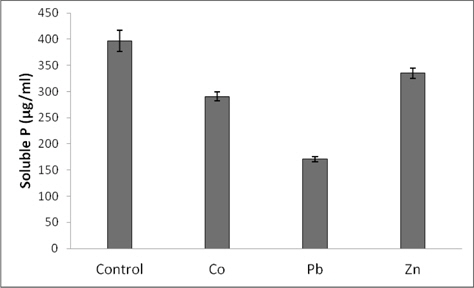
Phosphate solubilization as measured by the amount of phosphorous released into the medium is depicted in Fig. 3. As indicated by the results, the strain was shown to be capable of utilizing tricalcium phosphate as the sole source of phosphate. However, the presence of metals in NBRIP medium (200 mg/L) caused reduction in phosphate solubilizations. Compared with the control, reductions were 26, 56, and 19%, respectively for Co, Pb and Zn.
>
Effect of bacterial strain on growth and metal uptake by H. annuus
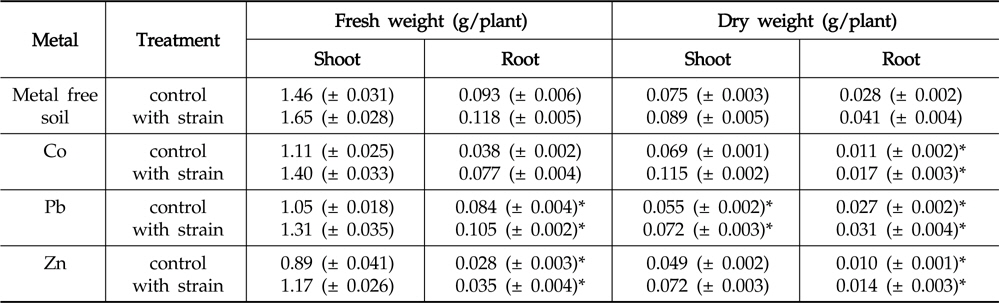
Inoculation with the strain resulted in increased fresh and dry biomass of
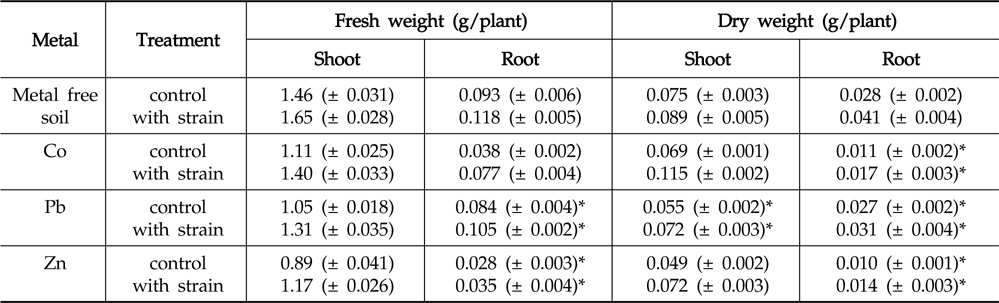
Effect of inoculation with Enterobacter ludwigii PSB 28 on shoot and root weight of Helianthus annuus
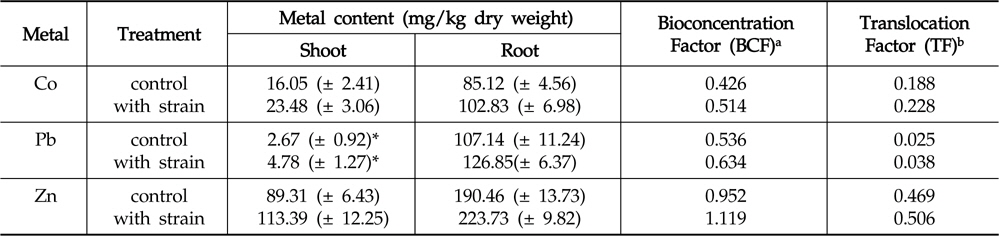
The amounts of Co, Pb and Zn accumulated in the roots and shoots of
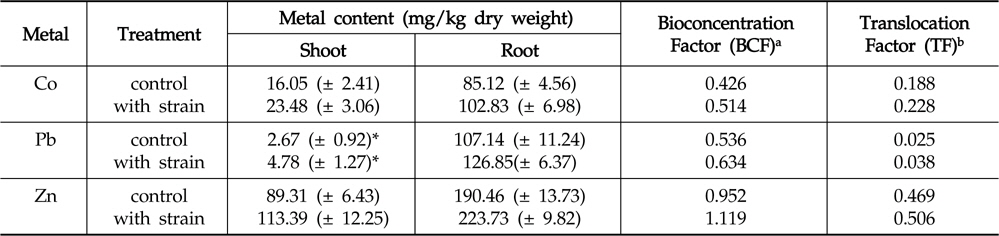
Effect of inoculation with Enterobacter ludwigii PSB 28 on accumulation and translocation of Co, Pb and Zn in Helianthus annuus
>
Mobility of the metals in soil
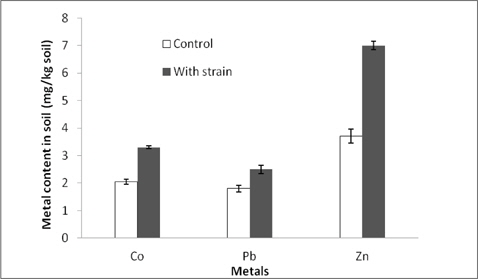
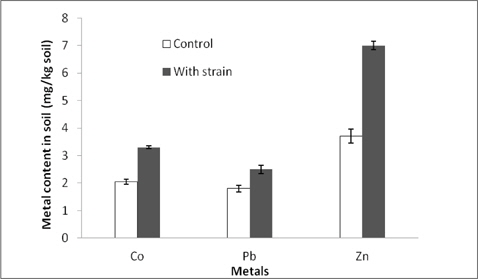
Metal mobilization potential of the strain was assessed in a batch experiment and the results showed that inoculation of the strain could increase the contents of water soluble metals in the soil. The mobilization of Co, Pb and Zn was respectively 41, 27 and 48% higher than those of the control soil (Fig. 4).
Growth response of the present strain under metal contamination conditions is in line with Rajkumar et al. (2008) and Prapagdee et al. (2013), who observed Zn, Cu and Ni resistance in
The effectiveness of the strain as a plant growth-promoter was assessed with
Regardless of inoculation or non-inoculation, the accumulation of metals in root systems was found to be considerably higher than that of in shoots. This could primarily be attributed to the poor translocation of heavy metals from roots to shoots (Rajkumar
As observed by Walpola and Yoon (2013), acidification of the medium could facilitate the inorganic phosphate solubilization by the PSBs. According to them, the acidification occurs mainly through the production of low molecular weight organic acids such as oxalic acid and gluconic acid. Analogous to their findings, an inverse relationship between pH and soluble phosphorus concentration from Ca3(PO4)2 by
The present findings of metal mobilization are in agreement with Wu
Inoculation with