To elucidate the variation of species composition, biomass and length distribution of flatfishes by the period and region in the Uljin marine ranching area, bottom trawl survey was investigated at the six stations from Feb. 2008 to Dec. 2010. During the survey period, a total 4 families 17 species in the Pleuronectiformes, average 69,158 ind./km2 and 5,625 kg/km2 were identified, in 2008 appeared in 14 species, average 25,798 ind./km2 and 2,333 kg/km2 and in 2009 appeared in 13 species, average 102,360 ind./km2 and 5,634 kg/km2 and in 2010 appeared in 14 species, average 75,704 ind./km2 and 8,632 kg/km2. The individual dominant species, occupying over 10% of total individuals, was
연안어장은 그 동안의 과도한 이용과 해양오염 및 연안의 매립과 간척사업 등으로 인하여 어장환경이 훼손되어 어획량이 감소하는 경향을 보이고 있는데, 우리나라의 경우 연근해 어업의 총어획량은 1980년대 이후 감소하고 있는 실정이다 (Fujita, 1987; Yotsui and Maesako, 1993; MIFAFF, 2008). 이러한 수산자원 감소를 방지하기 위한 여러 방안 중에 바다목장 사업은 인공어초, 해조장 등 인공구조물을 설치하여 수산자원의 서식장 및 산란장을 제공하여 어류자원을 지속적으로 증대시키고, 대상해역 생태계의 해양특성과 환경수용력을 우선적으로 고려하여 방류대상종과 방류규모를 결정하고 안정적인 수산물 생산 및 공급체계를 구축하여 잡는 어업에서 기르는 어업으로 전환하는 것을 목적으로 하는 사업이다 (MOMAF, 2005; MIFFAF, 2008; Lee et al., 2012). 또한 바다목장 사업은 어장을 적극적으로 선택하고 조성하는 과정에서 생물자원의 현존량 및 생태특성에 관한 자료를 축척함으로써 효율적인 어류자원의 관리모델을 개발하는데도 그 목적이 있다.
가자미목(Pleuronectiformes)의 넙치과(Paralichyidae)와 가자미과(Pleuronectidae) 어류 중에서 넙치(
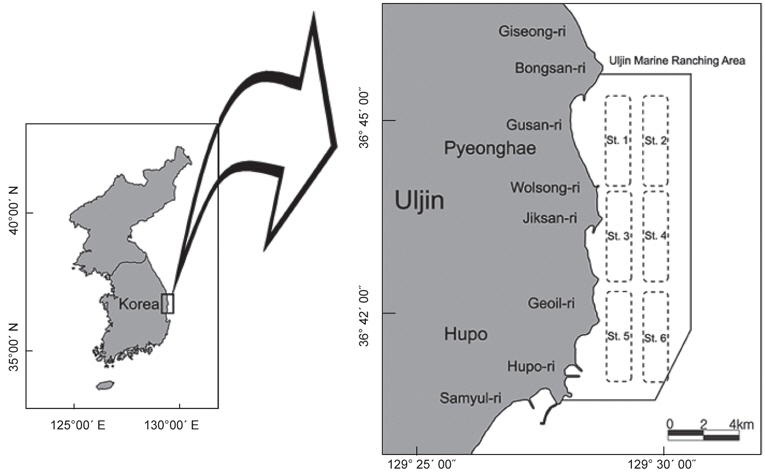
울진바다목장 해역은 경북 울진군 근남면 구산리에서 후포면 후포리 일대 20 km2로 한류와 난류가 교차하기 때문에 계절별로 여러 어종이 출현하는 것으로 잘 알려져 있다. 울진바다목장에서 거일리를 중심으로 한 남쪽 해안은 암반이 잘 발달되어 있고, 직산리, 월송리 및 구산리 일대의 수심이 낮은 연안은 다른 해역에 비해 사질의 함량이 높은 특징을 가지고 있다. 본 조사는 2008년 2월부터 2010년 12월까지 월별로 6개의 정점에서 소형저인망을 사용하여 채집을 실시하였다(Fig. 1). 본 조사에 사용된 어구의 크기는 길이가 약 10 m, 망폭이 약 2.6 m였으며, 각각의 정점에서 1.5~2.5 k't의 속도로 10~30분간 예망하였다.
각 정점으로부터 얻은 가자미목의 종별 개체수 및 생체량 자료는 단위 면적당 (km2)으로 환산하였으며, 가자미목 어류군집의 구조를 이해하기 위하여 출현 종 수 및 개체수 자료를 이용하여 종다양도지수(
정점간 유사도분석(similarity analysis)은 정점별로 출현한 종수와 개체수 자료를 바탕으로 유사도를 분석하여 백분율(%)로 나타내었다. 유사도분석 시에 생물자료간의 편중을 피하기 위해 모든 자료는 fourth root로 변환하였으며 Bray-Curtis 지수(Bray and Curtis, 1957)를 사용하여 정점간 유사도를 표현하였다. 정점간 유사도를 바탕으로 하여 군집분석을 실시할 때에는 PRIMER 6.0 프로그램을 사용하였으며, 그룹간의 연결 방식에는 complete linkage를 사용하였고, 그룹간 유사도를 바탕으로 수지도(dendrogram)를 작성하여 집괴분석(cluster analysis)을 실시하였다. 또한 정점간 그룹간의 공간적 분포 특성과 유사성 차이를 알아보기 위하여 다차원척도분석(MDS, Multi Dimensional Scaling)을 실시하여 2차원 공간에서의 군집구조의 차이를 파악하였다. 군집분석 결과 분리된 그룹간의 종수 및 개체수의 유의한 차이를 파악하기 위해서 one-way ANOSIM 분석을 실시하였다. 또한 각 정점군과 정점군 사이의 생물군집 차이를 유발하는 종을 분석하고 이들이 군집의 차이에 기여하는 정도를 알아보기 위하여 SIMPER분석을 실시하였다(Clarke and Warwick, 2001).
시간의 변화에 따른 가자미목 어류의 개체수, 생체량, 종다양도와 종우점도의 변화를 분석하기 위해서는 SPSS version 12.0 for Windows로 paired t-test를 실시하였고, 정점별 개체수 및 생체량의 차이와 연도별 체장분포의 차이를 분석하기 위해서는 ANOVA test를 실시하고 사후분석으로는 Turkey-T 분석을 실시하였다.
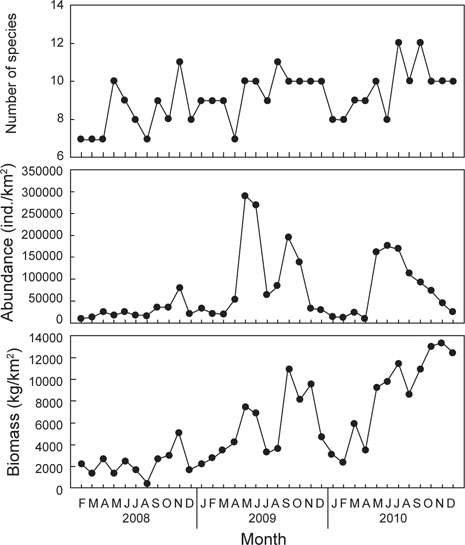
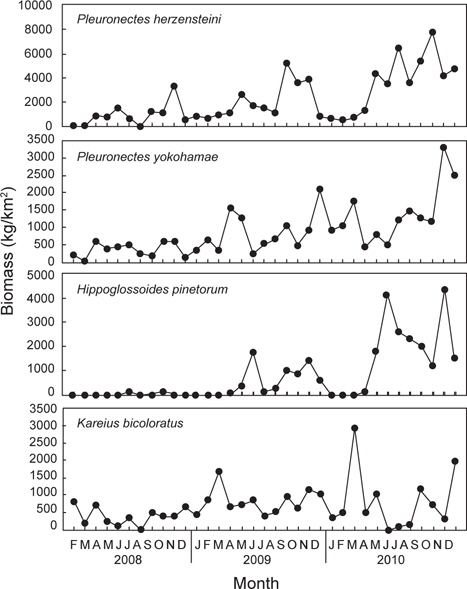
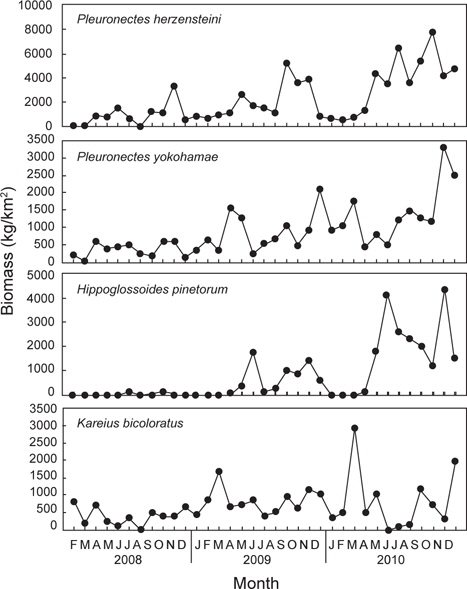
2008년 2월부터 2010년 12월까지 울진바다목장 해역 6개 정점에서 출현한 가자미목 어류는 총 4과 17종으로 가자미과(Pleuronectidae)가 12종으로 가장 많았고, 넙치과(Paralichthyidae)는 3종, 납서대과(Soleidae)와 참서대과(Cynoglossidae)는 각각 1종이 출현하였다. 2008년에는 총 3과 14종, 2009년에는 총 3과 13종, 2010년에는 총 3과 14종이 출현하였는데, 월별로 출현한 종수를 연도별로 비교한 결과 2008년에 출현한 종수에 비해 2009년과 2010년에 출현한 종수가 증가한 것으로 나타났지만(
조사기간 동안 출현한 가자미목 어류의 평균 개체수는 69,158 ind./km2이었는데, 2008년에는 평균 25,798 ind./km2로 2009년과 2010년에 비해 적은 개체수를 보였던 반면(
조사기간 동안 출현한 가자미목의 평균 생체량은 5,625 kg/km2로 2008년에는 평균 2,333 kg/km2로 생체량이 가장 적었고(
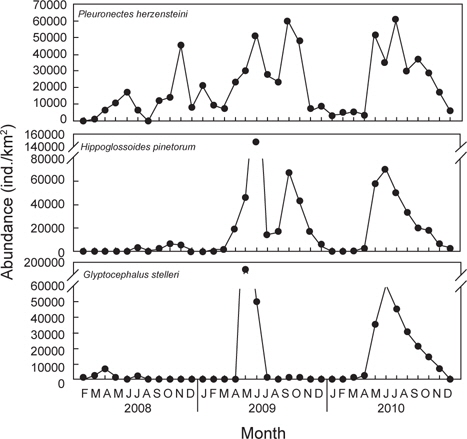
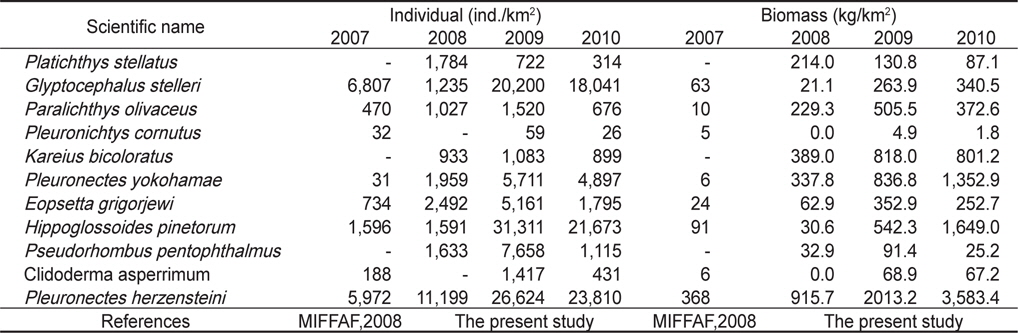
조사기간 동안 채집된 생물들 중에서 전체 개체수의 10% 이상을 차지하는 주요 우점종을 분석한 결과 참가자미(
조사기간 동안 채집된 생물들 중에서 전체 생체량의 10% 이상을 차지하는 주요 우점종을 분석한 결과 참가자미가 평균 2,207 kg/km2 (39.3%)로 가장 높은 생체량을 보였는데, 2008년에는 평균 916 kg/km2, 2009년에는 평균 2,013 kg/km2, 2010년에는 3,583 kg/km2로 시간이 경과함에 따라 생체량이 지속적으로 증가하는 경향을 보였다 (
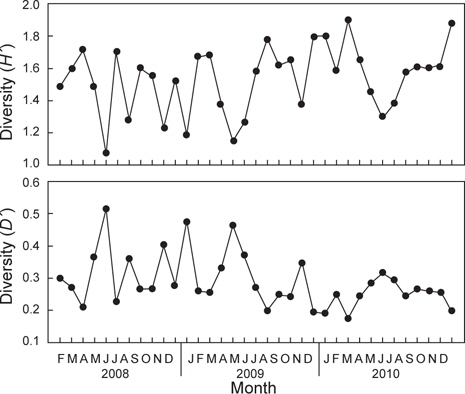
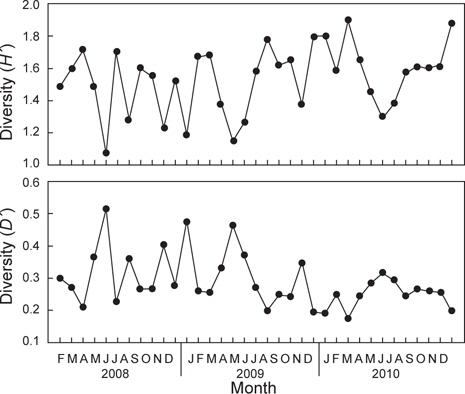
조사기간 동안 출현한 종수와 개체수를 바탕으로 월별 생태지수를 분석한 결과 종다양도(
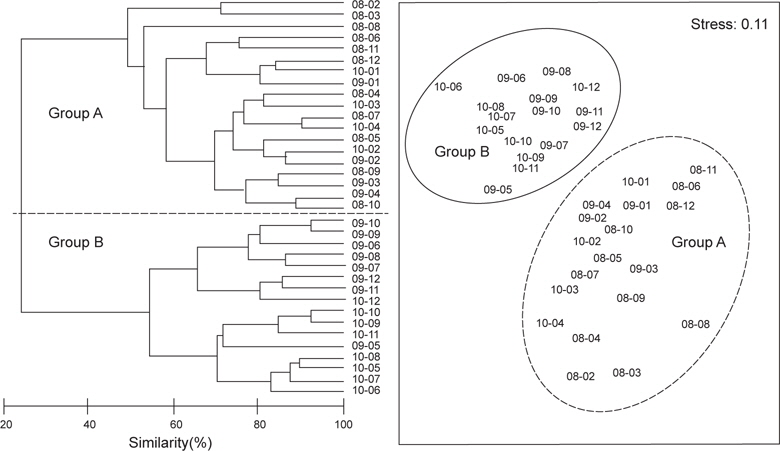
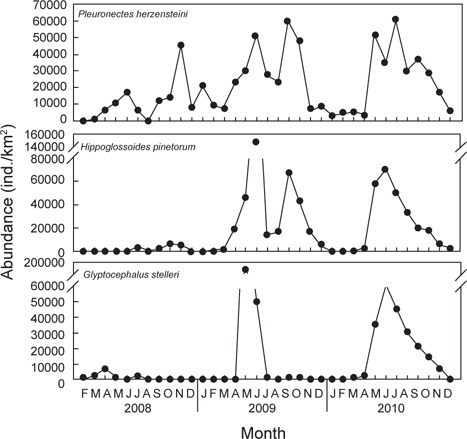
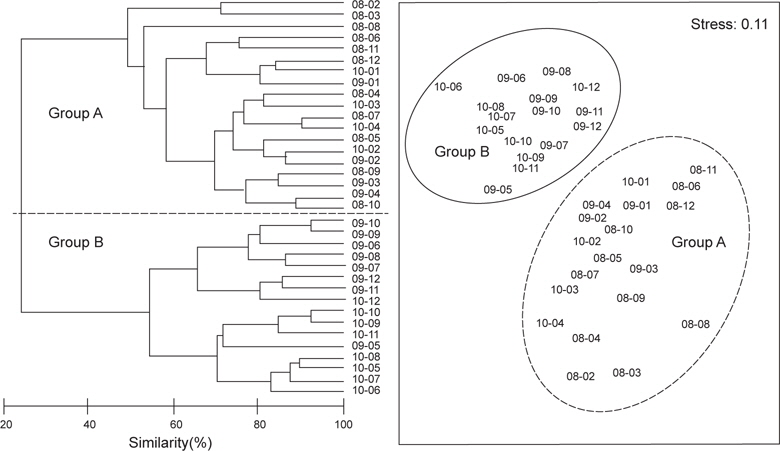
2008년 2월에서 2010년 12월까지 월별로 5회 이상 출현한 종수와 개체수를 바탕으로 월별 유사도를 구하고 집괴분석과 다차원척도분석을 통해 생물군집의 월변화의 유사성을 분석하였다. 그 결과 생물군집은 크게 2008년과 2009년 1~4월, 2010년 1~4월에 출현한 생물군집(Group A)과 2009년 5~12월과 2010년 5~12월에 출현한 생물군집(Group B)으로 구분되는 경향을 나타냈다(Fig. 7,
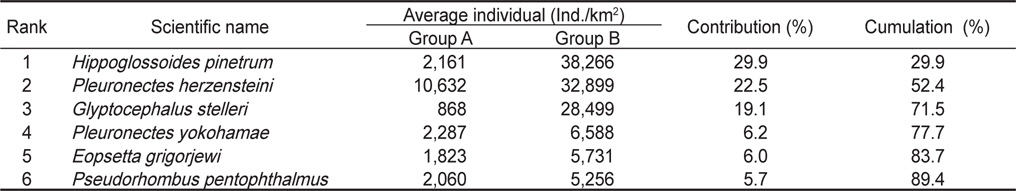
군집분석에서 Group A와 Group B 사이의 차이를 유발하는 것에 대한 종기여도를 분석하기 위하여 SIMPER분석을 실시한 결과 기여도에서 10% 이상을 차지하는 종들은 총 3종으로 이들은 Group A에 비해 Group B에서 개체수가 급격히 증가한 경향을 보였다. 용가자미(
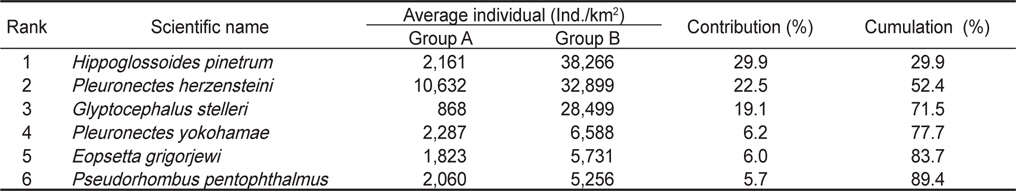
Species contributions to dissimilarity between Group A and Group B in the Uljin marine ranching area from Feb. 2008 to Dec. 2010
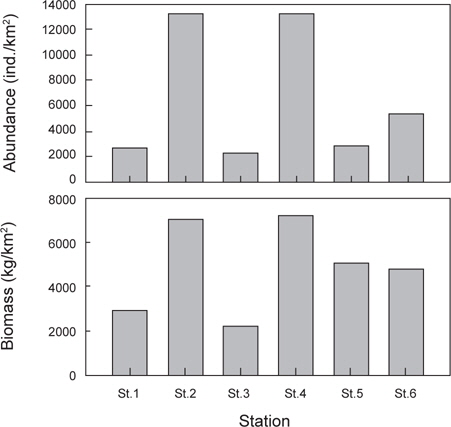
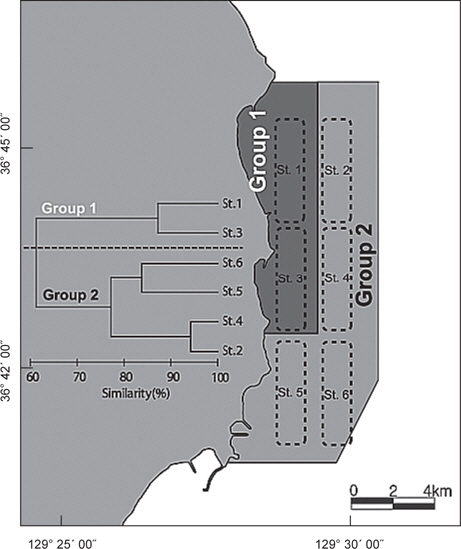
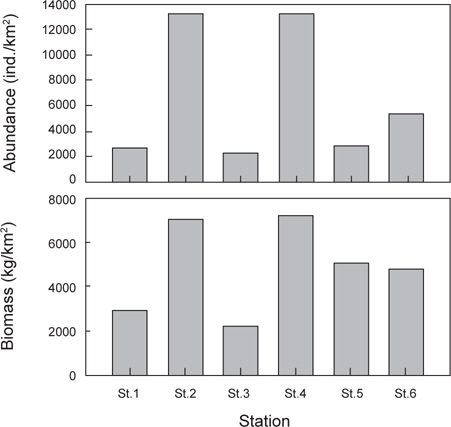
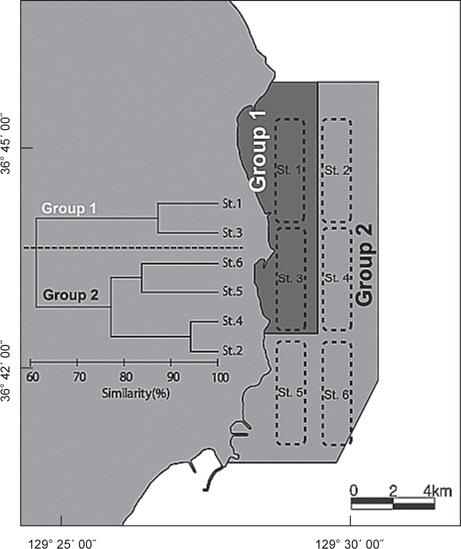
정점별로 출현한 생물군집을 바탕으로 정점간 유사도를 구하고 집괴분석과 다차원척도분석을 실시한 결과 조사해역은 크게 두 개의 그룹으로 분리되었다(Fig. 8). Group 1은 수심이 낮고 모래의 함량이 높은 지역에 해당하는 정점 1과 정점 3이 포함되었고, Group 2에는 수심이 상대적으로 깊고 니질의 함량이 높은 정점 2, 정점 4, 정점 5와 정점 6이 포함되었다.
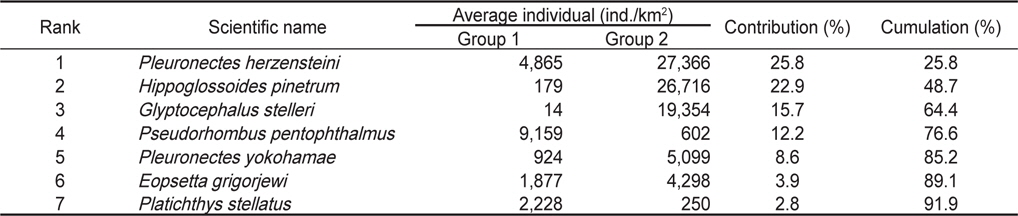
군집분석에서 Group 1과 Group 2 사이의 차이를 유발하는 것에 대한 종기여도를 분석하기 위하여 SIMPER분석을 실시한 결과 기여도에서 10% 이상을 차지하는 종들은 총 4 종으로 나타났는데, 참가자미(
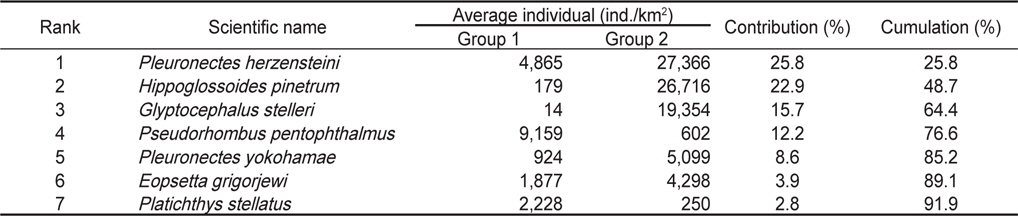
Species contributions to dissimilarity between Group 1 (St.1 and St.3) and Group 2 (St.2, St.4, St.5 and St.6) in the Uljin marine ranching area from Feb. 2008 to Dec. 2010
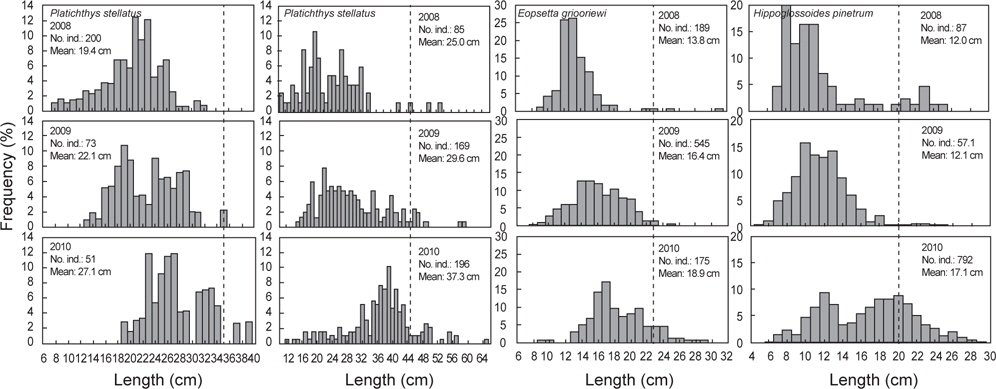
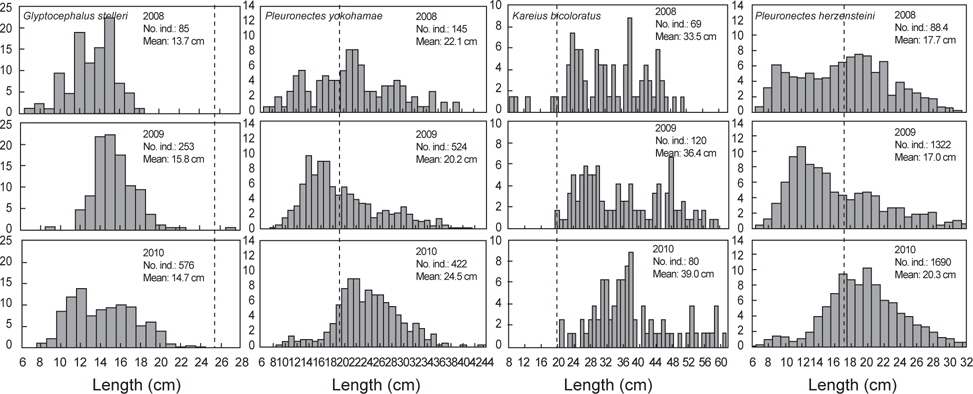
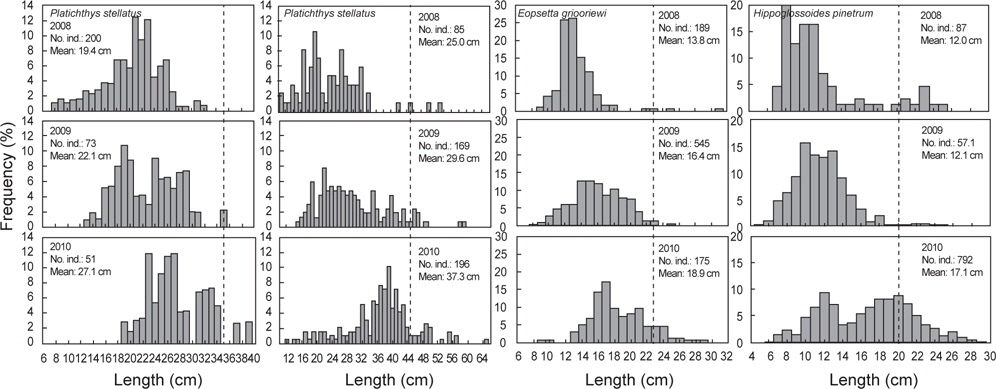
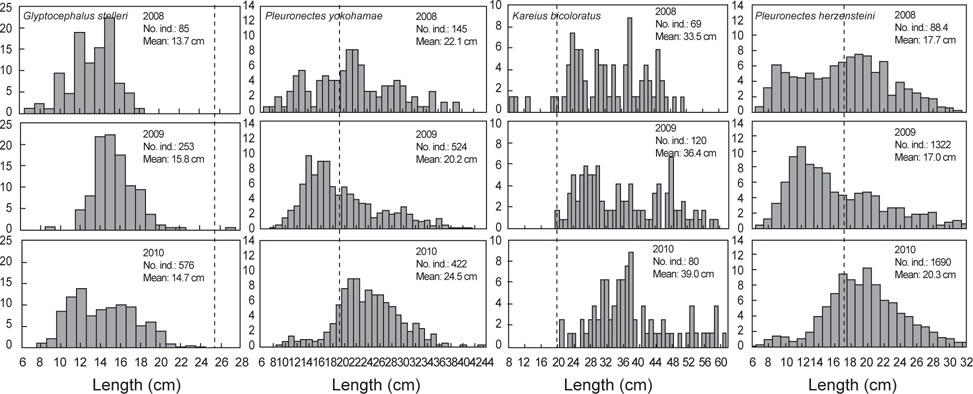
2008년 2월에서 2010년 12월까지 출현한 가자미목의 평균 체장분포의 변화를 분석한 결과 강도다리(
2008년 2월부터 2010년 12월까지 울진바다목장 해역에서 소형저인망을 이용하여 가자미목의 현존량과 분포특성을 조사한 결과 출현 종은 총 4과 17종으로 서식환경 및 조사방법에 따라 약간의 차이가 있지만 광양만에서는 8종(Cha and Park, 1997), 거제도 남쪽 해역에서는 10종(Youn and Shim, 2000), 부산의 수영만에서는 11종(Kim et al., 2000), 가덕도 주변의 해역에서는 13종(Huh and An, 2000)이 출현하여 본 해역이 다른 해역에 비해 많은 종들이 출현했다. 광양만과 수영만의 경우 본 조사에 비해 수심이 낮고 니질의 함량이 높으며 폐쇄적인 내만의 특성 때문에 본 조사에 비해 출현 종수가 적었던 것으로 생각된다. 가덕도의 경우 13종으로 가장 많은 종이 출현하였는데 가덕도는 낙동강에서 유입되는 많은 영양염으로 일차생산성이 높아 어류의 먹이원이 되는 새우류와 요각류 등 많은 무척추동물이 번성하여 다양한 가자미목 어류들이 출현한 것으로 판단된다. 서해안에서 저인망을 이용한 조사에서는 학암포에서 7종, 연포에서 7종(Noh et al., 2009), 서해 중부연안의 조사에서는 8종(Lee et al., 2010)이 출현하여 남해와 본 조사에 비해 종수가 적은 것으로 나타났는데, 서해 조사에서는 퇴적물 중에 사질의 함량이 높은 곳에서 서식하는 특성을 가진 가자미목 어류들만 제한적으로 출현하여 종수가 적었던 것으로 판단된다. 동해의 영일만에서 자망에 어획된 가자미목은 총 14종으로 다른 지역에 비해 많은 종이 출현하는 경향을 보였는데(Hong et al., 2008), 영일만의 경우 10~30 m 수심으로 본 조사와 수심이 유사하고 폐쇄적인 특성이 적으며 다양한 퇴적환경을 가지고 있어 많은 종의 가자미목 어류가 채집된 것으로 보인다. 자망은 저서성 어류인 가자미목에 대한 어획강도가 높고(Hwang et al., 1997), 저인망도 저서성과 반저서성 어류를 주로 어획하는 어구로 본 조사와 영일만의 조사에서 채집된 종수의 차이는 어구에 의한 차이보다는 본 조사지역은 다양한 퇴적환경 조성되어 있고, 조사 횟수가 많았던 것 때문으로 생각된다. Lee (2011)는 2002년 후포연안에서 오터트롤을 이용한 조사에서 가자미목 어류가 4종이 출현하였다고 보고하였는데, 이는 본 조사에 비해 조사지역과 조사 수심이 한정되어 있었고, 총 4회 조사로 조사횟수 또한 적었기 때문으로 판단된다.
본 조사결과 채집된 가자미목 어류 중에서 전체 개체수의 10% 이상을 차지하는 우점종은 참가자미, 용가자미, 기름가자미였고, 전체 생체량의 10% 이상을 차지하는 우점종은 참가자미, 문치가자미, 용가자미, 돌가자미로 나타났다. 남해의 수영만의 조사에서는 문치가자미와 개서대(
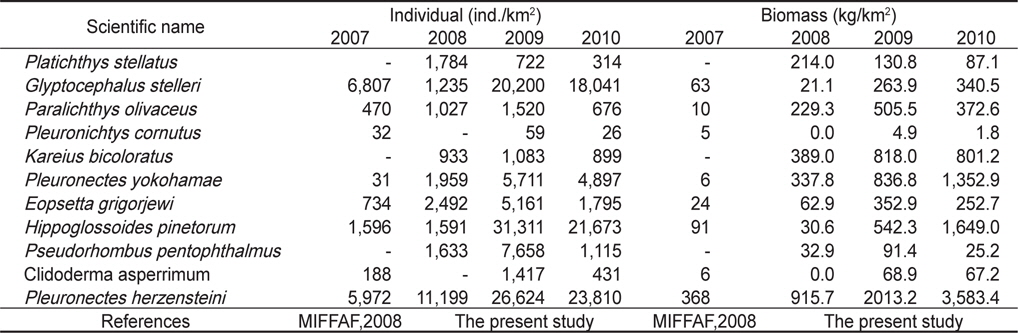
Individual and biomass of dominant Pleuronectiformes in the Uljin marine ranching area from 2007 to 2010
생물군집의 월별 유사성을 분석한 결과에서 2008년과 2009년 1-4월과 2010년 1-4월에 출현한 군집(Group A)과 2009년 5-12월과 2010년 5-12월에 출현한 군집(Group B)으로 구분되었는데, Group A에서 평균 개체수는 24,700 ind./km2이었고, Group B에서는 평균 121,600 ind./km2로 더 많은 개체가 출현하였다(
2009년과 2010년 1-4월에 채집된 가자미목 어류들에 비해 2009년과 2010년 5-12월에 채집된 가자미목 어류들은 종수와 개체수가 모두 높은 것으로 나타났는데 (
가자미목 어류들의 공간적 분포 특성을 분석한 결과 fine sand (2.7phi)의 퇴적물 입도조성을 보이고 수심이 10 m 내외로 낮으며 암반과 인공어초의 분포가 낮은 곳(MOMAF, 2005; MFAFF, 2008)에 위치한 정점(St. 1과 St. 3)들이 Group 1로 구분되었고, 다양한 인공어초가 투하되어 있고 곳곳에 암반이 넓게 분포하며 very fine sand와 clay의 함량이 높은 퇴적물 특성(3.2phi)을 가지며 평균 30~40 m의 상대적으로 수심이 깊은 곳(MOMAF, 2005; MFAFF, 2008)에 위치한 정점(St. 2, St. 4~St. 6)들이 Group 2로 구분되었다. Group 1에서는 점넙치가 평균 9,159 ind./km2, 참가자미가 4,864 ind./km2, 강도다리가 2,228 ind./km2, 넙치가 2,203 ind./km2로 우점하는 것으로 나타났는데 이들은 퇴적물의 입도조성에서 사질의 함량이 높은 곳에서 높은 서식밀도를 보이는 종들로 Group 2 지역에 비해 많은 개체수를 보였다 (
가자미목의 체장분포를 살펴본 결과 강도다리, 넙치, 물가자미로 평균 체장이 시간이 지남에 따라 점차 증가하는 경향을 보였지만 강도다리는 성숙체장이 35 cm (Ralston, 2005)로 개체군중에서 성숙개체들의 비율은 2008년 0%, 2009년 2.2%, 2010년 5.6%, 넙치의 성숙체장은 45 cm로 개체군중에서 성숙개체의 비율은 2008년 5.6%, 2009년 4.7%, 2010년 6.5%, 물가자미의 성숙체장은 23 cm (Kim et al., 2011)로 개체군 중에서 성숙개체의 비율은 2008년 1.1%, 2009년 0.7%, 2010년 8.6%로 대부분의 개체들이 미성숙 개체들로 나타나 울진바다목장 해역이 어린 개체들의 성육장으로 이용되긴 하지만 산란장으로 이용하지 않는 것으로 판단된다. Hwang 등 (2012)이 2009년 11월-2010년 10월까지 울진바다목장에서 강도다리의 번식능력에 대한 연구를 수행한 결과에 따르면, 본 해역에서 채집된 강도다리는 대부분 성숙체장에 이르지 못하여 바다목장 내에서 재생산은 미비한 것으로 나타났지만 강도다리와 넙치같이 방류했던 개체들의 평균 체장이 시간이 지남에 따라 지속적으로 증가하는 것으로 보아 바다목장 해역에서 정착하여 서식하는 것을 알 수 있었다. 강도다리와 넙치의 경우 미성숙 개체들이 점차 성숙체장에 이름에 따라 차후 연구를 통해 바다목장 해역 내에서의 번식특성에 대한 연구와 다른 가자미들과의 먹이경쟁과 서식밀도에 따른 개체군의 변동에 대한 연구가 지속적으로 수행되어야 할 것으로 판단된다. 기름가자미 또한 유사한 경향을 보였는데 기름가자미의 성숙체장은 25.6 cm (Cha et al., 2008)로 조사기간 동안 채집된 개체들의 99% 이상이 미성숙 개체를 보였다. 일반적으로 기름가자미는 40~700 m의 해역에서 광범위하게 서식하며 산란기에 연안으로 이동하여 산란하고 치어일 때는 연안에서 서식하다 성장하면서 외해로 나가는 생태적 습성을 지니고 있어 (Cha et al., 2008), 본 바다목장 해역을 성육장으로 이용하지만 바다목장 해역에서는 산란하지 않는 것으로 판단된다. Lee (2011)의 후포연안에서 수행한 연구에서도 14~18 cm 크기의 기름가자미가 가장 많이 분포했으며, 성숙체장 이상의 기름가자미는 어획되지 않아 본 조사와 유사한 경향을 보였다.
반면 돌가자미, 문치가자미, 용가자미, 참가자미는 성숙개체들이 어획비율이 상대적으로 높은 특성을 보이고 있었는데, 돌가자미의 성숙체장은 25.0 cm (Jun et al., 2003)로 바다목장 해역에서 어획된 개체의 85% 이상이 성숙개체들로 나타났고 문치가자미의 성숙체장은 19.8 cm (Seo et al., 2010)로 성숙개체의 비율이 2008년 56.6%, 2008년 36.6%, 2010년 75.4%, 용가자미는 성숙체장이 20.0 cm로 성숙개체의 비율이 2008년 10.5%, 2009년 1.2%, 2010년에 33.0%, 참가자미의 성숙체장은 17.8 cm (Cha et al., 2006)로 2008년 성숙개체의 비율은 42.8%, 2009년 32.5%, 2010년 58.3%를 나타냈다. 이들 개체군들은 바다목장 해역에서 산란하고 어린 개체들이 바다목장을 보육장과 성육장으로 이용하며 성숙개체가 되면 다시 바다목장 내에서 재생산을 하는 것으로 판단된다. 이들 개체군들은 다른 가자미목 어류들에 비해서 상대적으로 개체수와 생체량이 높은 특징을 보여 다른 가자미목 어류들에 비해 먹이와 서식지 경쟁에서 우위에 있는 것으로 판단되며 바다목장 해역의 높은 일차생산성을 고려할 때 점차적으로 개체수가 증가할 것으로 보인다.
울진바다목장 해역에서 가자미목의 군집구조의 변화를 분석한 결과 가자미목 어류들 중에서 강도다리, 넙치, 물가자미는 바다목장에서 산란을 하지는 않지만 어린 개체들을 성육하는 성육장으로 이용하는 것으로 판단되고, 돌가자미, 문치가자미, 용가자미와 참가자미는 울진바다목장에서 산란하고 어린 치어들의 보육장과 성육장으로 적절하게 이용하는 것으로 판단된다. 울진바다목장 해역의 방류 대상종인 강도다리와 넙치의 경우 지속적인 방류가 이루어지고 있으며 방류된 개체들은 바다목장 내에서 안정적으로 정착하여 성장하고 있으며, 이들 개체군들이 성장함에 따라 성숙개체 이상이 되면 바다목장 내에서 재생산이 이루어질 수도 있기 때문에 지속적인 모니터링이 필요할 것으로 판단된다. 또한 울진바다목장의 환경수용능력과 퇴적물 특성을 고려하여 강도다리와 넙치와 같은 가자미류 어류들의 방류량과 방류 장소를 선정하여야 하며, 2차 방류대상종의 선정에 있어서 상대적으로 서식밀도가 낮은 정점 1과 3에서 정착하여 서식할 수 있는 종을 고려하여야 할 것으로 판단된다.