Since the industrial revolution, seawater has increased in temperature and acidity due to the increase in anthropogenic carbon dioxide in the atmosphere. To understand the effect of elevated CO2 on the early life stages of fish, we reared fertilized eggs of black sea bream
해양산성화는 대기중 이산화탄소가 해수로 녹아들어가면서 화학적 반응에 의해 pH가 낮아지는 현상으로, 지구온난화와 더불어 대기 중 이산화탄소의 농도 증가로 나타나는 대표적인 지구환경변화이다. 해수의 산성화 경향에 기인한 해양생태계의 변화 연구는 2000년대 들어와서 세계적으로 활발하게 진행되고 있으나(Feely et al., 2004; Orr et al., 2005), 아직까지 국내에서 연구가 많이 이루어지지 않은 실정이다. 현재 학계에 보고된 연구의 대부분은 산성화된 환경에서 무척추동물의 생존이나 성장에 관한 것이었다. 특히 외골격이 CaCO3로 구성된 동물 플랑크톤이나 산호, 패류의 경우, 산성화된 해수는 패각에 직접 작용하여 패각을 녹여내거나 형성을 지연시켜 이들의 생명현상에 큰 영향을 미치고 있다(Delille et al., 2005; Talmage and Gobler, 2010). 반면, 척추동물인 어류에 대한 연구는 아직 활발하지 않은데(Fabry et al., 2008), 이는 활발히 움직이는 어류를 대상으로 하는 실험환경이 패류 등에 비해 상대적으로 어려운 점도 있으며, 무척추동물에 비해 내골격을 가지는 척추동물은 해수의 pH 변화를 간접적 경로로 영향받거나 환경변화에 능동적으로 대처할 수 있기 때문에(Ishimatsu et al., 2005) 상대적으로 해양산성화로 인한 영향을 적게 받을 것으로 예상되기 때문이다. 그러나 해양산성화로 인한 생물들의 반응이 종별, 생장 단계별, 서식지별 등으로 달리 나타나므로(Doney et al., 2009; Kroeker et al., 2013), 해양생태계의 상위단계 포식자이며 인류의 단백질 공급에 큰 기여를 하는 어류를 대상으로도 적극적인 연구가 필요할 것으로 생각된다. 본 논문에서는 이러한 중요성에 인식하여, 어류 중 현재 상업적 어업과 양식종으로 이용되고 있는 감성돔(
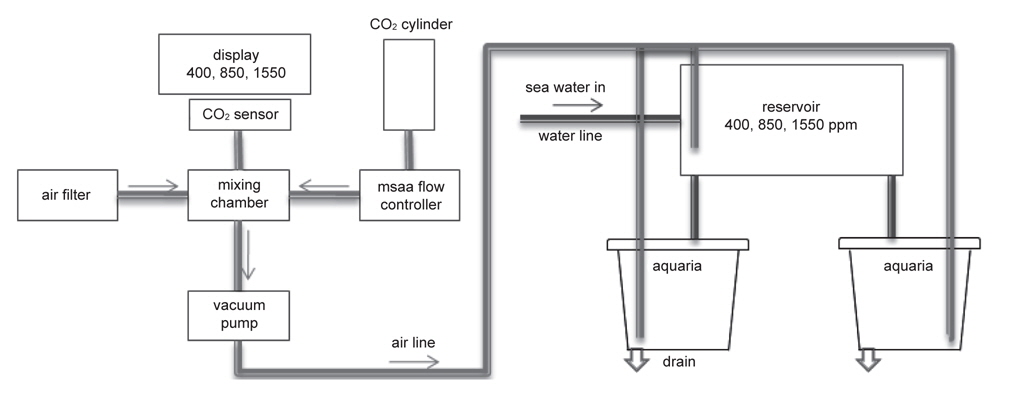
어류를 대상으로 해양산성화에 의한 영향을 파악하는 데 적합한 실험장치를 설계하고 제작하였다(Fig. 1). 이 장치는 원하는 농도의 이산화탄소를 만드는 공기 혼합조와 생물을 사육하는 사육조로 크게 2단계로 구분된다. 외부공기는 이산화탄소 흡습제(soda lime)가 충진 된 공기필터(air filter)를 통과하면서 이산화탄소가 완전히 제거된 상태로 혼합조(mixing chamber)로 유입되고, 이산화탄소가스는 정밀 유량계(mass flow controller, MFC, KOFLOC Co., Model 3660 Series, Japan)로 일정하게 혼합조로 유입되어, 결국 혼합조 내부는 이산화탄소가 제거된 공기와 순수 이산화탄소 기체가 섞이게 된다. 이처럼 형성된 혼합조 내부의 공기는 이산화탄소 농도 측정기(CO2 sensor, SOHA Tech., SH-VT250, Korea)를 사용하여 연속적으로 측정 하면서 농도가 일정하게 유지되도록 하였다. 혼합조의 공기는 다시 진공펌프(vacuum pump, KnF co., N86KT.18, France)를 통하여 사육수 저장고(reservoir)와 사육 수조(aquaria)로 각각 지속적으로 공급되며(Fig. 1), 이를 통해 저장고와 사육수조에서 원하는 수준의 이산화탄소 농도를 유지할 수 있도록 하였다. 사육수조는 용량 60 L의 원형 수조를 사용하였으며, 뚜껑을 설치하여 대기와 사육수간의 기체 교환으로 인한 사육수 내의 이산화탄소 농도 증감을 최소화하였다. 또한 사육수조의 물을 교환할 때, 저장고에서 물이 보충되게 하여 환수로 인한 이산화탄소 농도의 급격한 변화를 방지하였다. 수조는 한 개의 저장고와 두 개의 사육수조가 한 세트로 총 3 세트(대조구, 저농도구, 고농도구)를 운용하며, 각각 다른 이산화탄소 농도로 유지하였다.
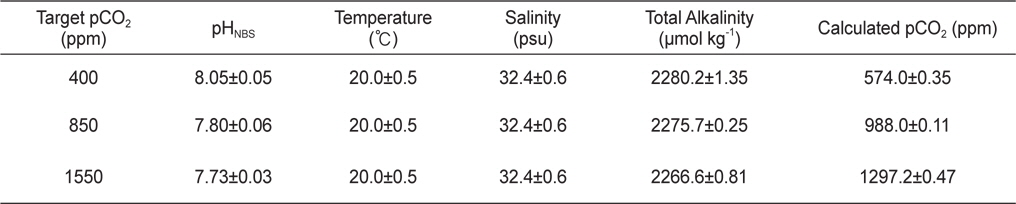
사육수의 이산화탄소는 대조구가 400 ppm, 저농도구와 고농도구는 각각 850 ppm와 1550 ppm로 설정하였으며, 대조구 농도는 현재 대기 이산화탄소 농도 수준이며, 850 ppm과 1550 ppm은 IPCC의 배출시나리오(SRES) 중 중간(A1B)과 최악(A1F1)의 시나리오에서 각각 예상하는 2100년 대기 이산화탄소 농도에 해당한다(IPCC, 2007). 이때의 pH는 대략적으로 대조구 8.1, 저농도구 7.9, 고농도구 7.7의 수준으로, 각 실험구별로 pH 미터(5 Star-pH meter with DO sensor, Thermo Scientific Orion, USA)를 이용하여 매일 3회 측정하여 그 수치를 확인하였다. 한편 공기 혼합조에서 측정되는 이산화탄소농도와 사육수의 pH를 검정하고 그 외 탄소계 인자 분포를 파악하기 위해, 사육수의 총알칼리도를 측정하였다(Dickson et al., 2007). 이때 총알칼리도는 개방용기 적정법을 이용하여, Dr. Andrew G. Dickson(Scripps Institute of Oceanography, University of California, San Diego, U.S.A) 실험실에서 제작한 표준물질로 (reference materials) 검정하였다. 해수 탄소계 인자 중 총용존무기탄소와 이산화탄소는 실험실에서 측정한 pH와 총알칼리도를 이용하여 CO2SYS프로그램(http://cdiac.ornl.gov/ftp/co-2sys, Pierrot and Wallace, 2006)으로 계산하였다(Table 1). 그 결과, 원하던 수준의 이산화탄소농도와 실제 사육수조의 이산화탄소 농도 사이에 다소 차이가 있었다: 대조구(약 570 ppm)와 저농도구(약 1000 ppm)는 다소 높았으며, 고농도구(약 1300 ppm)는 다소 낮았음. 그러나 대조구에 비해 두 실험구의 이산화탄소 농도가 월등히 높으므로, 해양산성화의 영향을 평가하기에 큰 문제가 없는 것으로 생각된다. 또한 대조구의 이산화탄소농도는 외부공기와 교환이 원활하지 않는 밀폐된 실내에서 흔히 경험할 수 있는 농도범위로, 감성돔의 양식환경을 고려한다면 현실적인 농도범위에 속한다고 할 수 있다.
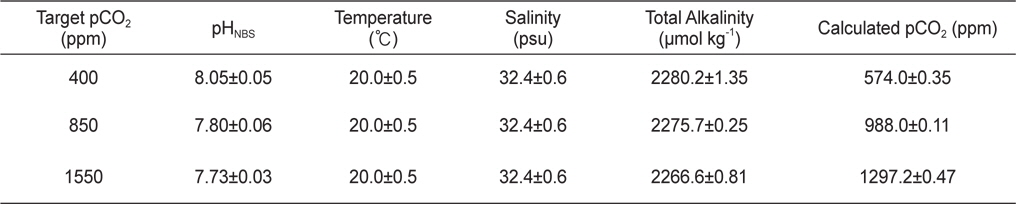
Chemical properties for seawater in three experimental tanks in July. Calculated pCO2 was computed by CO2SYS software (Pierrot and Wallace, 2006) using measurements of pHNBS, total alkalinity, temperature, and salinity
감성돔을 대상으로 하는 해양산성화 사육실험은 2011년 3월에 약 3주간 수행되었으며, 이때 사용한 감성돔 수정란은 2011년 2월 25일 전남 여수에 위치한 경양수산에서 공급받았다. 감성돔 수정란은 실험실에서 약 8시간의 순치를 거친 후 6개의 사육 수조(이산화탄소 농도 3수준×2 반복)에 수조당 2000립씩 수용하였다. 사육 수온은 20±0.5℃로 유지되었으며, 사육수의 용존산소 농도는 모든 수조에서 약 10 mg L-1로 유지되게 하였다. 사육을 위한 광주기는 14L:10D로 설정하였으며, 사육수는 수정란 부화 후 7일까지는 교환하지 않았으며 7일 이후부터 교환을 시작하여, 점점 교환율을 증가시켰다. 감성돔의 먹이는 부화 후 2일째부터 로티퍼(
해양산성화 실험장치에서 감성돔 수정란을 3주간 사육 후 생존개체에 대하여 체장과 체중을 측정하였다. 체장 측정은 각 개체별로 실체현미경(Carl Zeiss co., Discovery V8, Germany) 하에서 카메라(Carl Zeiss co., ICC1, Germany)를 이용하여 사진을 찍어, 분석 프로그램(Carl Zeiss co., Axiovision4.7, Germany) 을 통해 개체의 체장을 정밀하게 측정하였다. 체중 분석은 일반적인 저울을 이용하여 측정하기에는 시료 각 개체의 중량이 매우 작으므로 다음과 같은 방법을 이용하였다. 시판되는 micro tube (AXYGEN INC., PCR-02-C, USA)에 일련 번호를 적은 후, 각 번호에 해당하는 micro tube의 무게를 정밀저울(Mettler Toledo co., AB204-S, Switzerland)을 이용하여 측정하였다. 이후 각 micro tube에 감성돔 치어를 한 마리씩 넣어서 무게를 다시 측정하여, 그 차이로 치어의 무게를 계산하였다.
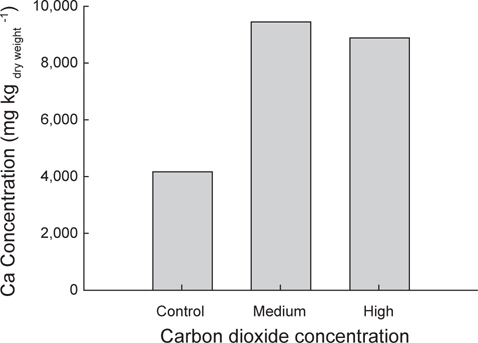
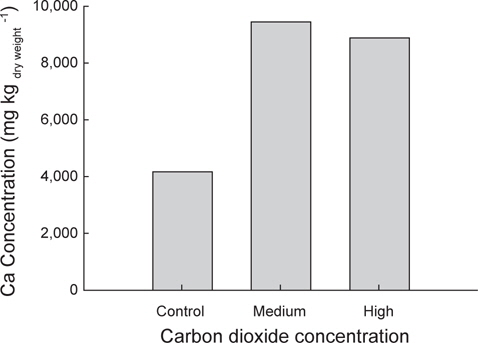
또한 시료(치어) 내에 존재하는 칼슘의 농도를 측정하기 위하여, 각 실험구의 시료 대부분을 동결건조(SAMWON ENG.,SFDSF06, KOREA)하고 곱게 갈아서 분말로 만들었다. 이후 한국기초과학지원연구센터(부산센터)에서 ICP-OES (ACTIVA, JY HORIVA, JAPAN)로 분석하였다. 이상의 시료 분석 결과는 Minitab 16 (Version 16.2.1, USA) 프로그램으로 One-way ANOVA 통계 분석을 하였다.
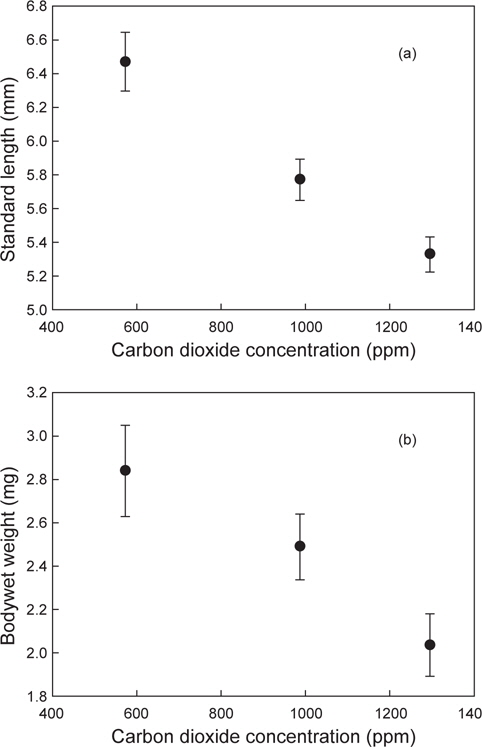
현재와 미래에 예상되는 이산화탄소 농도에서 감성돔 치어를 3주일 동안 사육한 결과, 사육수 내의 이산화탄소 농도가 증가함에 따라 감성돔 자치어의 성장이 저해되는 것을 확인할 수 있었다(
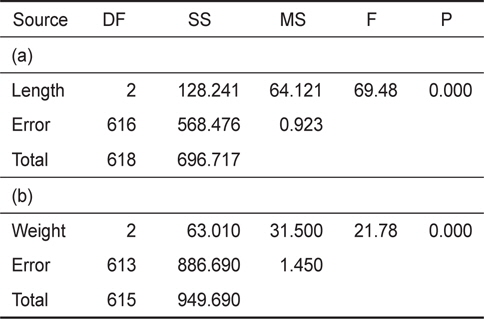
[Table 2.] Results of statistical analysis (one-way ANOVA). (a) is length, (b) is weight result
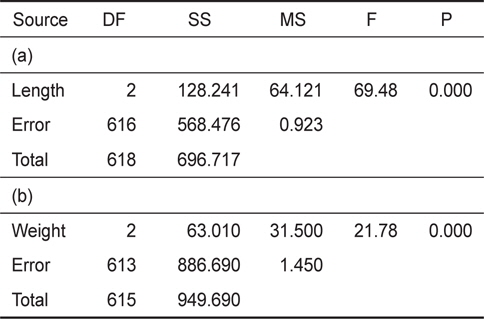
Results of statistical analysis (one-way ANOVA). (a) is length, (b) is weight result
한편, 일부 연구자들은 해양산성화의 일반적인 결과와 반대로, 이산화탄소 농도가 증가할수록 어류 성장에 좋은 영향을 미친다는 결과를 제시하였다(Morgan et al., 2001; Munday et al., 2009; Checkley et al., 2009). Munday et al. (2009)는 크라운피쉬(orange clownfish,
한편 Checkley et al. (2009)는 농어의 일종인 white sea bass(
이상에서 해양산성화 환경에서 감성돔을 수정부터 자치어 단계까지 3주간 배양한 결과 산성화가 진행될수록 개체의 체장과 체중이 감소하는 것으로 나타났으나, 어체의 칼슘농도는 오히려 증가하는 것으로 나타났다. 이로서 이산화탄소가 높은 환경은 감성돔의 초기발달단계에서 체장과 체중 성장에는 다소 나쁜 영향을 미칠 것으로 생각된다. 반면, 산성화환경이 감성돔 체내 칼슘축적이나 이석을 포함한 골격형성에 미치는 영향은 추후 연구를 통해 감성돔의 초기발달단계에서 산-염기 평형 유지능력과 연계하여 검정할 필요가 있을 것으로 생각된다.
이처럼 해양산성화는 해양생물의 성장, 발달, 체성분 등에 영향을 미칠 뿐만 아니라, 동일 분류군(또는 동일 종) 중에도 그 반응의 정도 및 양상이 다르게 나타나며, 더욱이 한 개체의 생체영역에 따라서도 영향이 달리 나타났다. 따라서 여러 연구자에 의한 것처럼(Doney et al., 2009; Kroeker et al., 2013) 해양산성화로 인한 환경변화에 해양생물의 반응을 종합하기에는 그 반응 형태가 매우 다양하고 특이성이 있으므로 앞으로의 연구는 이를 반영하여 다각도로 진행되어야 할 것으로 생각된다. 특히, 미래환경에서 수산자원의 안정적인 공급을 위해, 다양한 (양식)어류를 대상으로 해양산성화의 영향을 정밀하게 평가하고 이를 토대로 정책수립에 반영할 수 있는 기초자료를 제시해야 할 것으로 생각된다.