In order to understand the co-culture impact of the sea squirt
경남연안해역은 1990년대 이후 연안해역의 생산성 향상을 위해 대대적인 정비사업을 실시한 이래 비교적 안정적 생산량을 유지하여 왔다. 그러나 최근 주변 연안지역의 지리적 변화와 양식환경의 변화로 인하여 굴을 비롯한 여과섭식성 양식생물의 생산성이 저하되고 있어 어장의 이용실태가 기존과는 많이 변화하고 있는 실정이다. 특히 연안개발로 인한 양식해역을 폐쇄할 경우에도 어장을 소멸하지 않고 다른 해역으로 이설하여 국지적 밀식현상을 초래하고 있고, 보다 수익성이 높은 품종으로 품종변화를 무차별적으로 시도함으로써 이종간의 먹이경쟁을 초래하고 있다(Jeong, 2011).
멍게와 굴같은 여과섭식성 생물은 해수 중에 부유하고 있는 부유성 유기입자를 여과 섭취하여 에너지원으로 이용하므로 동일 해역에서 이들 생물군의 생체량은 해역의 먹이량에 의하여 한정된다고 볼 수 있지만(Jeong et al., 2009), 두 종의 먹이 선택성(Kang et al., 2009) 및 먹이섭취량(Kim, 2003)등의 차이가 있는 것으로 보고되고 있어 두 종간의 혼서가 서로에 미치는 영향에 대한 정확한 평가는 매우 어렵다, 그러나 이론적으로 유지 가능한 집단 생체량의 범위는 양식생물의 요구에너지량이 양식해역에서 섭취 가능한 에너지량의 한계를 초과하지 않는 범위로 한정된다고 할 수 있으므로(Fiala-Mèdioni, 1978), 주변 환경수로부터 섭취하는 먹이의 양을 정량적으로 평가하는 것은 두 종간의 먹이경쟁을 이해하는 기초자료가 될 수 있을 것이다.
최근 통영연안에는 수하식 양식생물이 다양화 되고 있고, 이들 종간의 직간접적인 먹이경쟁은 불가피하다. 또한 수하식 양식생물의 종에 따른 생리생태적 특성의 차이는 배설물의 침강 등에 의한 오염부하량의 가중으로 인하여 보다 빠른 양식환경 변화를 초래할 수 있다(Kingsley and Watabe, 1987; Jeong and Cho, 2003). 우리나라 양식시설기준은 이러한 생물학적 특성을 고려하지 않고 허가면적대비 시설률의 획일적인 시설기준을 적용하고 있다. 생물학적 특성이 서로 다른 다양한 생물을 같은 해역에 수용하여 양식할 경우, 생태학적으로 우위를 점하는 생물의 생존 및 성장의 주도권을 가지게 되므로 그렇지 않은 생물의 생산성을 크게 저하시킬 우려가 있다. 또한, 성장이 빠른 생물일수록 다량의 배설물을 분비하므로 양식어장 노화를 촉진시키는 등 이차적인 피해가 발생할 수 있다(Jeong and Cho, 2003.)
여과섭식성 수산생물 양식어장의 생산성을 안정적으로 유지하기 위해서는 이들 생물의 계획적 생산이 필수적이고, 이는 어장 수용력 내지는 환경용량의 범위로 제한되어야 할 것이다. 그러나 굴(
실험에 사용한 멍게는 경남 통영시 인근 북만의 멍게양식장에서 채집하여 군산대학교 천해양식실험실로 옮겨 100 L들이 순환여과식수조에 연안 수온과 근접한 수온 5℃에서 3일간 절식후 시판중인 혼합 미세조류(Shellfish Diet 1800TM, ReedMariculture Inc., USA:
순치된 멍게는 수온 5-25℃의 범위에서 5℃간격으로 5개의 실험구를 설정하여 여수율을 측정하였다. 각 수온구간별 측정시에는 1℃/day의 간격으로 점차적으로 상승시켰고, 실험수온에서 2일간 순치시켰다. 여수율 측정 시에 시료채취에 의한 외부영향을 최소화하기 위해 유수식 수조에 별도의 1 L들이 아크릴수조를 설치하여 각각 1개체씩 수용하였고(n=10), 이후 수위를 낮춰 각 수조별 먹이생물의 농도변화를 측정하였다. 여수율은 동일개체를 2회 반복 측정하였고 각 측정치는 이들의 평균치를 사용하였다. 먹이생물농도는 각 실험수조에서 해수를 2 mL씩 채수하여 Kim (1995)에 따라 분광광도계(Optizen 3220UV bio, Mecasys Co. LTD, Korea)로 675 nm에서 chlorophyll-a의 농도변화를 측정하였다. 실험용 먹이생물은 순치시 사용하였던 혼합 미세조류와 동일하였고, 미세조류의 초기농도는 0.20±0.10으로 조절하였다. 여수율(Fr)은 Kim (1995)에 따라 아래 식과 같이 산정하였다.
Fr, L/h/animal = V·(1-e-Z)
이 때, V는 폐쇄순환계의 수량(L)에 해당하고, Z는 먹이농도의 감소계수를 나타낸다. 여수율을 측정한 개체는 전중(g), 육중 (g) 및 건조중량(g)을 0.1 g단위까지 측정하였다. 생체 건조중량은 껍질을 제거한 후 80℃에서 3일간 건조시킨 후 데시케이터에서 상온까지 방냉시킨 후 중량을 측정하였다.
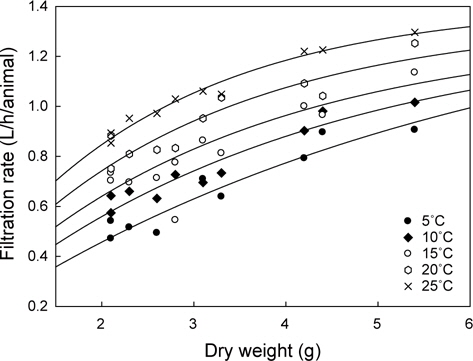
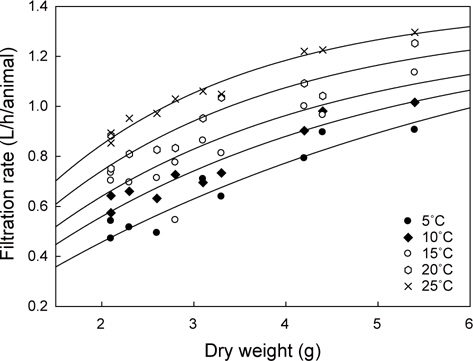
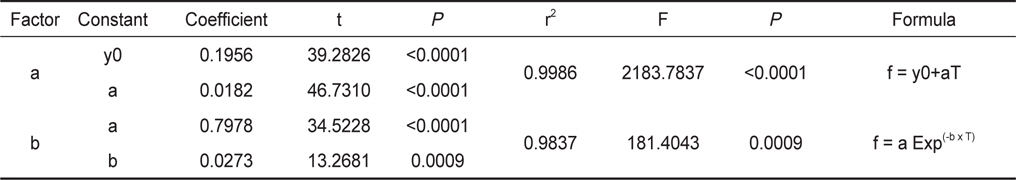
각 수온별 체중에 대한 여수율(Fr)의 변화를 예측하기 위해 최소자승법(FR=aXb)으로 회귀분석을 실시하였고(
멍게의 여수율을 조사한 결과는 Fig. 1과 같다. 수온과 체중에 따라 멍게의 여수율은 0.47-1.30 L/h/animal의 범위였고, 수온 및 체중에 비례하였다. 모든 수온에서 여수율은 체중이 증가함에 따라 증가하였다.
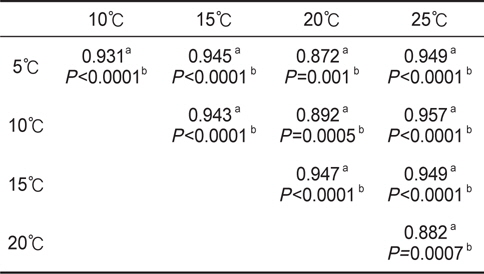
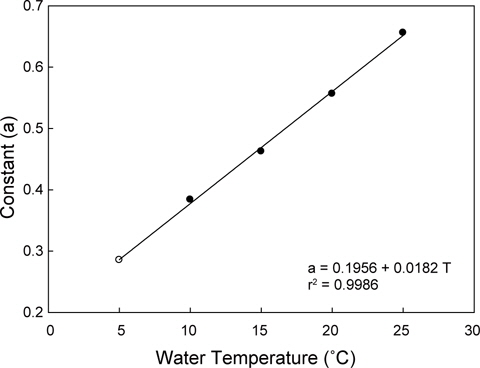
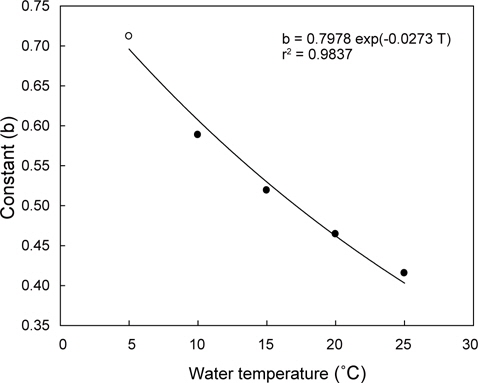
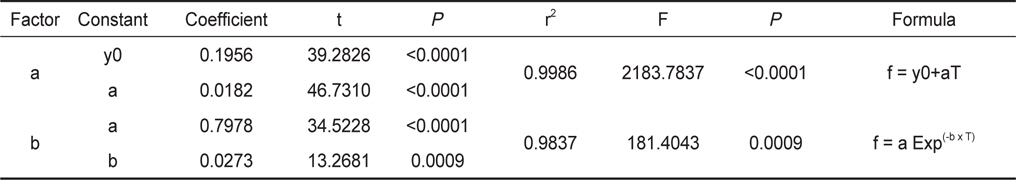
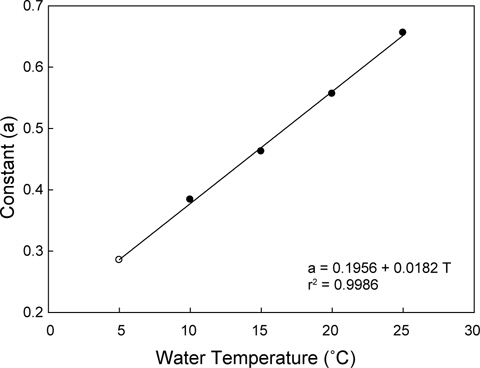
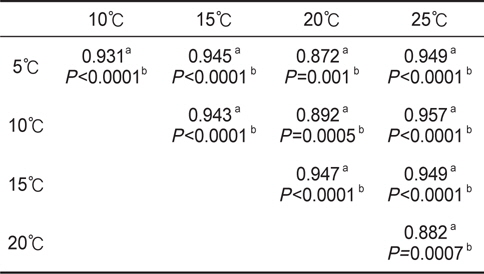
수온구별 여수율의 회귀분석결과의 공선성은 수온구별 여수율간의 상관관계를 분석을 통해 검증하였다. 각 수온구별 여수율의 Pearson 상관계수는 0.872-0.957의 범위로 매우 높은 정상관계를 나타내어 공선성이 인정되었다(Table 1). 따라서 이들 회귀선의 공통상수를 찾고자 회귀분석을 실시하였다. 상수(a)는 일차 선형회귀하여 a=0.1956+0.0182T로 추정되었다(Fig. 2, r2=0.9986,
Analysis of Pearson product moment correlation for filtration rate of Halocynthia roretzi in the experimental water temperatures (5-25℃) (n=10)
Statistical results for regression of constant (a) and (b) in the experimental water temperatures (5-25℃).
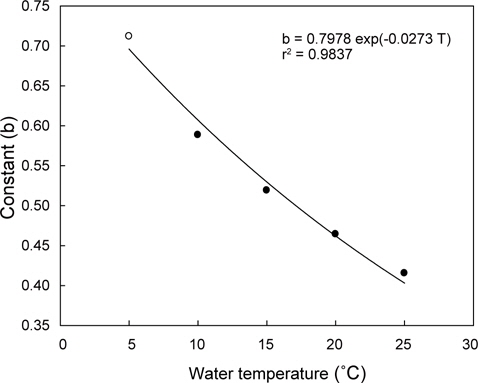
Fr(L/h/animal) = (0.1956+0.0182T)×DW0.7978EXP(-0.0273T) (5 ≤ T ≤ 25) 이때 T는 수온(℃)이고, DW는 육질부의 건조중량(g)에 해당한다.
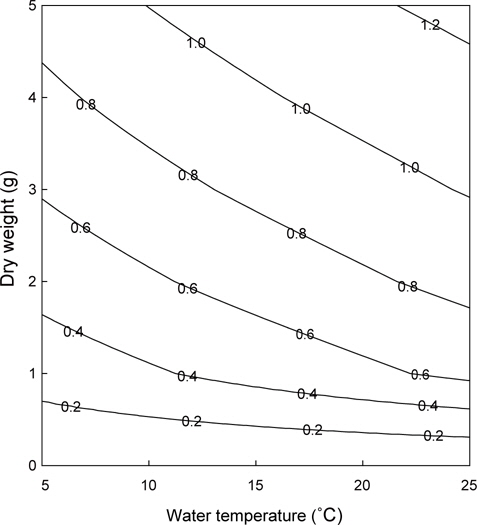
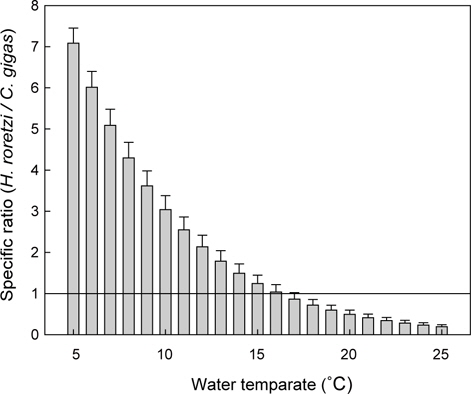
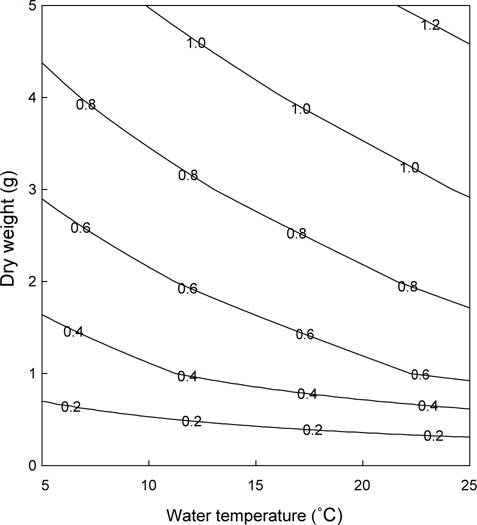
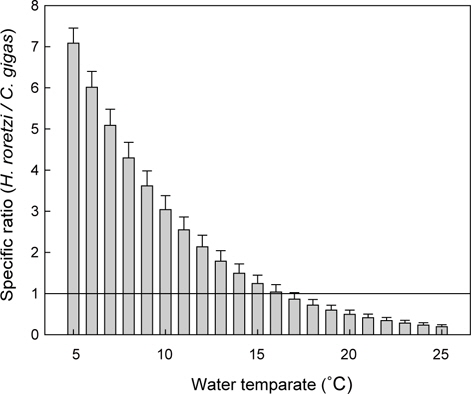
이상의 여수율 모델식을 이용하여 수온과 체중에 따른 등여수율분포도를 그려보면 Fig. 4와 같다. 동일체중인 멍게의 여수율은 저수온에서 보다 고수온에서 높았지만, 고수온일수록 등여 수율선의 간격이 넓어져 증가속도가 감소하였다. 특히 이러한 경향은 고체중의 개체에서 보다 확연하게 나타났다. 우리의 모델식과 굴의 여수율 모델(Kim, 1995)을 이용하여 동일 체중 및 수온조건에서 여수율을 비교면 Fig. 5와 같다. 멍게의 여수율은 수온 5℃에서 최대 7배 이상으로 높았지만, 수온 16℃를 기점으로 굴에 비해 낮은 것으로 나타났다.
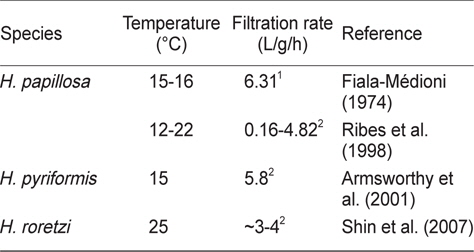
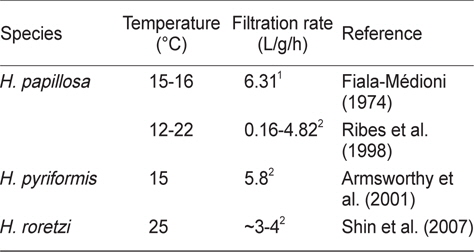
우리 연구결과 멍게의 여수율은 0.47-1.30 L/h/animal의 범위였다. 이는 기존 연구결과보다는 다소 낮은 값을 나타내었지만 (Shin et al., 2007), 이는 측정방법의 차이에 기인한 것이라고 할 수 있다. 즉, 이 연구에서는 먹이생물을 지표물질로 이용하였지만, Shine et al. (2007)은 Neutral red를 지표물질로 이용하였다(Cole and Hepper, 1954). 또한 우리의 연구에 활용된 멍게들은 실내 순환수조에서 순치시킨 점등을 고려하면 Shine et al. (2007)에 비해 다소간 낮은 측정치를 나타내고 있다. 그러나 다른 종이긴 하지만 Ribes et al. (1998)의 측정치의 범위 안에 포함되어(Table 3), 우리의 측정치를 이용하여 멍게의 여수율 모델식을 만들었다.
[Table 3.] Filtration rate (L/gDW/h) reported for different species in genus Halocynthia
Filtration rate (L/gDW/h) reported for different species in genus Halocynthia
멍게의 여수율에 대한 수온과 체중에 의한 영향을 평가한 결과 수온과 체중의 독립적 영향은 유의하였지만(
회귀선의 공선성은 일반적으로 상관계수가 0.7이상일 때 일반적으로 공선성을 고려하여야 하며, 특히 0.8-0.9이상일 때 공선성을 반드시 고려하여야 한다. 우리의 상관계수는 0.872- 0.957의 범위로(Table 1) 공선성을 인정할 수준에 해당하였다 (Hair et al., 1998; Wikiversity, 2010). 따라서 각 수온별 회귀선이 평행성이 인정된다고 가정하고, 이 회귀선들의 상수들에 대해 회귀분석을 실시한 하였다. 상수(a)는 체중 1 g일 때 각 수온별 여수율에 해당하고, 이들 상수는 Fig. 1과 같이 직선회귀하였다(r2=0.9986,
위의 결과를 통해 등여수율 분포도를 그리면 Fig. 5와 같다. 체중이 낮은 개체보다 체중이 높은 개체가 수온에 대한 여수율 변화가 컸고, 동일수온에서는 저체중에서 체중에 따른 여수율 변화가 크게 나타났다. 이는 냉수성 종인 멍게의 생리적 특징으로 판단된다. 그러나 수온이 높을수록 여수율은 증가하였고, 이는 곧 높은 먹이요구량을 의미하므로, 일반적인 수온에 따른 대사율의 반응을 고려한다면 멍게양식시 봄철 수온상승과 대형개체의 폐사량이 증가하는 것은 이러한 생리적 요인과 무관하지 않다고 할 수 있을 것이다. 그러나 봄철 대량폐사와 먹이공급능력에 관해서는 보다 많은 연구가 요구된다.
멍게와 굴의 식성에 대해서는 논란의 여지가 있다. 멍게와 같은 미색류인 미더덕의 육과 변의 지방산 조성을 조사한 결과, 굴과 미더덕의 식성이 다르다는 보고(Maf, 1997) 이후 굴과 멍게의 식성에 관한 논쟁은 오랜 기간 지속되어 왔다. 또한 멍게와 굴의 복합양식시 멍게는 주로 20 μm이하의 미립자유기물을 섭취한다고 보고됨으로써(Kang et al., 2009) 멍게와 굴의 복합양식이 폐사를 줄일 수 있는 하나의 방법으로 알려지고 있다(Hur, 2009). 그러나 우리의 연구에서 공급한 혼합 미세조류는 크기 5-20 μm의 미세조류를 공급하였고, 위 내용물을 분석한 결과 주로 소형미세조류가 주로 검경되었지만, Cho (2011)의 연구에서는 멍게 분(feces)에서 주로 대형미세조류가 다량 검출되었다. 이는 멍게가 주변해수 중에 부유하는 유기입자성 물질을 먹이선택적으로 섭식한다기 보다는 생리적 특성에 의한 소화 효율의 차이로 굴과 다른 소화체계를 지니고 있음을 의미할 수 있다. 실제 Ribes et al. (1998)에 따르면, 멍게는 먹이선택성이 없고, 0.5-100 μm 크기의 먹이를 섭취하고 특히 0.6 μm 이상의 크기에 대해 높은 효율을 지니며 (Studart and Klumpp, 1984) 규조류, 와편모조류 및 섬모충류 및 입자성유기탄소까지 섭취를 하는 것으로 보고하였다. 따라서 멍게와 굴의 혼서양식시 주변해수에 부유하는 입자성유기물에 대한 먹이경쟁은 불가피한 것으로 판단된다.
남해안에서 멍게양식은 저수온기에는 성장 등의 목적으로 연안해역으로 이식하게 된다(Seo, 1982; Kim and Moon, 1998; Kim, 2003; Jeong, 2011). 굴이나 멍게는 해수중에 부유성 유기물을 여과섭식하는 생물이므로 이들 생물의 생산력은 환경의 먹이가용성, 즉 수용력에 의해 결정된다(Jeong, 2002; Jeong et al., 2009). 따라서 멍게와 굴의 먹이경쟁을 밝히기 위해서는 두 생물의 포식압을 평가할 필요가 있다. 멍게와 굴과 같은 여과섭식성생물의 포식압은 여수율로 평가할 수 있어, 우리는 기존굴의 여수율(Kim, 1995) 을 동일한 조건에서 적용하여 여수율을 비교하였다. 그 결과 16℃이상의 수온에서는 동일한 체중조건에서 굴의 여수율이 멍게보다 높게 나타났고(Fig. 5), 이는 봄철 수온상승기 이후 연안에 이식된 멍게는 굴과의 먹이경쟁에서 다소 불리한 조건에 노출될 수 있음을 의미한다. 따라서 최근멍게의 대량폐사 해결방안으로 제시된 혼서양식을(Hur, 2009) 위해서는 두 종간의 먹이경쟁관계에 대한 보다 심층적인 연구가 필요할 것으로 판단되고, 이는 멍게의 생리적 특징에 대한 연구가 선행되어야 할 것으로 판단된다.