The growth, respiration, ingestion, and ecological efficiencies of
최근 새로운 증·양식 대상어종 중 부가가치와 선호도가 높은 어종으로 부각되고 있는 쥐노래미(
쥐노래미는 최근 연안 서식장의 환경 악화와 무분별한 남획으로 인해 급격히 감소되고 있다. 따라서 본 종을 포함한 연안어족자원의 증가를 도모하기 위해서는 적절한 어획 및 자원관리가 요구되며 산업화를 위한 인공종묘의 대량생산기술개발이 필요하다.
지금까지 쥐노래미에 관한 연구는 난·치자어에 있어서 형태변이와 사망에 미치는 온도의 영향(Hamai and Kyushin, 1966), 초기생활사(Fukugara and Fushimi, 1984) 등이 있다. 우리나라의 경우는 체장과 체중의 상관관계(Kim, 1977), 치어의 적정사육밀도(Kim and Chang, 1998), 난발생과 자어 형태(Kim et al., 1993), 종묘생산을 위한 생물학적 연구(Lee, 1999), 번식상태 및 자치어의 성장(Kang, 2002)등 많은 연구가 되어 있지만 자치어기 동안의 생태생리학적 연구는 보고 된 바 없다.
본 연구에 있어서 동물의 성장, 호흡 및 동화효율 등의 생리, 생태적 연구는 에너지흐름의 역학적 구조를 이해하고 생태계의 기능을 파악하는데 있어 유용한 수단이 된다. 생태계는 복잡한 먹이사슬과 같은 영양학적 구조와 집단 간 에너지흐름에 밀접하게 관련되어 있으며, 이러한 복잡한 먹이망 속에서 그 물질의 전환에 따른 에너지 흐름을 파악하기 위해서 에너지수지를 분석하는 것이 매우 중요하다 (Klekowski and Ducan, 1975).
이와 같이 어류가 먹이를 통하여 획득한 모든 에너지는 체내에 축적되거나 대사과정을 통한 상실 및 배설질소와 분의 형태로 배출됨으로써 에너지 수지의 균형을 나타낸다. 특히 에너지 대사에 관한 연구는 수온, 염분 및 먹이 등의 환경에 대한 동물의 대사적 유형과 다른 영양 단계간의 에너지 적응유형을 이해하는 총체적인 지표가 될 수 있으며, 그 동물의 생산성을 추측하는데 유용하다.
어류의 생리, 생태적 연구의 중요한 자료인 에너지 수지에 관한 연구는 타 어종에 있어서 많이 보고되었으며(Kitchell et al., 1977 ; Eldridge et al., 1982 ; Dabrowski et al., 1988), 본 종과 관련된 연구로는 자연에서 채집된 노래미의 섭식량과 성장량으로부터 에너지 대사의 추정(Kim and Zhang, 1998)에 관한 연구가 유일하다. 또한 환경요인과 관련한 양식어류의 생리적 변화 및 에너지수지에 관한 연구는 주로 틸라피아(Jauncey, 1982)와 무지개송어(Elliott, 1976; Staples and Nomura, 1976) 등에서 볼 수 있다.
위와 같은 수산동물의 에너지흐름에 대한 생리, 생태학적 연구는 동물의 생활사에 관한 전반적인 양상을 설명해주며 자원증식과 종묘생산기술을 체계화 할 수 있는 중요한 요인이 된다. 따라서 본 연구에서는 몇몇 종에 한정되어 있는 우리나라 해산양식 현황에 있어 최근 산업적으로 가치가 높은 쥐노래미를 대상으로 부화 후의 자치어 기간 동안 성장에 가장 큰 요인으로 작용하는 수온과 먹이를 변수로 하여 이에 대한 성장, 호흡 및 생태효율을 비교, 조사하여 에너지수지를 규명함으로서 에너지흐름에 대한 생리, 생태적 기초자료를 얻는데 그 목적이 있다.
실험어는 2005년 11월 23일 충청남도 태안반도 부근에서 자망으로 어획된 성숙한 어미를 강원도 도립수산자원연구원 어류 사육동으로 즉시 운반하여 실내 사육수조에 수용하였으며 세균의 감염을 막기 위하여 Oxytetracycline HCL 성분인 Ultramysin 150 ppm에서 1시간 약욕시킨 후 1일간 안정시켰다. 성숙한 수컷은 몸빛깔이 등황색으로 뚜렷한 혼인색을 띄고 있었고, 암컷은 산란기에도 혼인색을 나타내지 않으나 복부가 팽만하여 있었다. 채란은 외상이 없고 안구가 선명하며 상태가 양호한 개체를 선별하여 암수의 비율을 1:1로 맞춘 후 복부압박법에 의해 2005년 11월 24일-28일(2회)에 걸쳐 실시하였다.
채란된 성수란과 정자는 건식법으로 수정하였으며 수정란 관리는 유수식의 사각틀망이 설치되어 있는 콘크리트 수조에 수용하였다. 수온은 자동온도 조절장치를 이용하여 12 ± 0.5℃로 일정하게 유지해 주었다. 수정 후 부화소요시간은 29일이 소요(약 696시간)되었다. 부화된 자어는 자연해수(5.1-8.5℃, natural seawater temperature, NT)와 가온해수(13 ± 0.5℃, heated seawater temperature, HT)로 구분하여 250 L (수용적: 200 L) FRP 수조에 각 먹이별 2반복으로 1,000마리씩 수용하였다. 사육수는 여과해수를 2 L/min 유수시키며 조절하였다. 부화 자어의 먹이공급은 난황흡수 후 먹이를 섭취하기 시작한 날로부터 2주간 rotifer,
자치어의 성장을 알아보기 위하여 실험 개시 일로부터 1주일 간격으로 각 수조 당 10마리씩 무작위로 선택하여 만능투영기 (Mitutoyo, PJ-A3000, Japan)로 체장을 0.001 mm 단위까지 측정하였다. 동일개체의 건중량 측정을 위해 열풍건조기(SANYO, MOV-212F, Japan)를 이용하여 60℃에서 24시간 동안 건조하여 0.1 mg 단위까지 측정하였다. 각 실험구의 성장은 실험 개체 모두의 평균값으로 나타내었고 6주 동안 각 실험구의 개체의 성장에너지는 실험 종료 시 에너지 값으로부터 실험 시작시 에너지 값을 공제해주었다. 생존율은 매일 사망한 개체를 파악하여 일주일 간격으로 합산 후, 전체 사육 개체수에 대한 누적 사망 개체수의 백분율로 나타내었다.
산소소비량을 측정하기 위하여 1주일 간격으로 각 수조 당 먹이를 섭취한 실험어 10마리씩 무작위로 추출하여 여과해수가 담긴 밀폐용기(500 mL)에 수용한 후, 12시간 동안의 산소소비량을 자동산소검량기(YSI 95)로 측정하고, 실험 전후의 용존산소차로서 정량화하였으며, 개체 1마리당 일간 산소소비량 (mg O2/ind./day)으로 나타내었다. 실험 전체기간 동안 산소소비에 이용된 에너지 값을 구하기 위하여 산소소비량은 대사열생산에 의해 소비된 에너지로 전환시켰다(1 mg O2 = 14.06 J) (Gnaiger, 1983).
개체의 산소소비량은 환경수의 산소분압에 영향을 받기 때문에(Prosser, 1973) 산소소비 측정이 끝날 때의 실험용기 내 용존산소량이 처음 용존산소량의 80% 이하로 떨어지지 않게 조절하여 주었다.
자치어의 섭취량을 알아보기 위하여 적정공급밀도(inds./mL)를 파악한 후 1주일 간격으로 각 수조 당 실험어 10마리씩 무작위로 추출하여 여과해수가 담긴 500 mL 용기에 수용한 후 NEA구과 EA구의
실험 기간 동안 분배출량은 극히 소량으로 먹이 급여 후 24시간 동안 방치하여도 siphon방식을 사용하여 수동적으로 분을 수거하기에는 불가능하였다. 따라서 분배출 에너지 함량은 에너지수지 방정식에 의거하여 섭취에 따른 성장, 호흡 및 뇨배설의 에너지함량 차로 구하였다.
뇨배설 양은 먹이를 충분히 먹고 난 실험어를 산소병에 각각 10마리씩 수용하고 24시간 후 phenol-hypochlorite method (Solorzano, 1969)으로 UV-Visible spectro-photometer (Varian DMS-1005)를 이용하여 흡광도를 측정하여 암모니아 질소를 정량한 후 개체 한 마리당 일간 질소배설량(ug atoms-N/ind./day)으로 표시하였다. 배설된 뇨의 에너지함량은 4.06 cal/mg (Brafield and Solomon, 1972)으로 계산하였다.
쥐노래미 자치어의 에너지 수지는 Standard IBP 방정식(Klekowski and Duncan, 1975)에 의거하여 다음과 같이 나타내었다.
I = P + R + F + U
여기서 I는 섭취 에너지, P는 체성장에 쓰인 에너지, R는 유지 에너지로써 호흡에 쓰인 에너지, F는 동화되지 않고 분으로 배출된 에너지, U는 뇨로 배설된 에너지를 나타내었다(Dawirs, 1983). 단, 분배출 에너지 함량은 에너지 수지 방정식에 의거하여 섭취에 따른 성장, 호흡 및 뇨배설의 에너지함량 차로 구하여 주었다.
쥐노래미 자치어의 섭취, 성장, 호흡 및 뇨배설 에너지를 측정한 자료로부터 아래의 생태효율을 계산하였다.
동화효율(Assimilation effi ciency, %) A = (P + R + U) / I ×100 총 성장효율(Gross growth effi ciency, %) K1 = P / I ×100 순 성장효율(Net growth effi ciency, %) K2 = P / A ×100
여기서 A는 총 섭취한 에너지에서 분배출로 사용된 에너지를 제외한 동화된 에너지를 백분율로 나타낸다. K1는 총 섭취 에너지에서 성장에 이용된 에너지를 백분율로 나타낸 것이며, K2는 동화된 에너지에서 성장에 이용된 에너지를 백분율로 나타내었다.
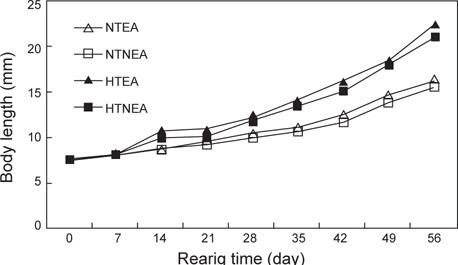
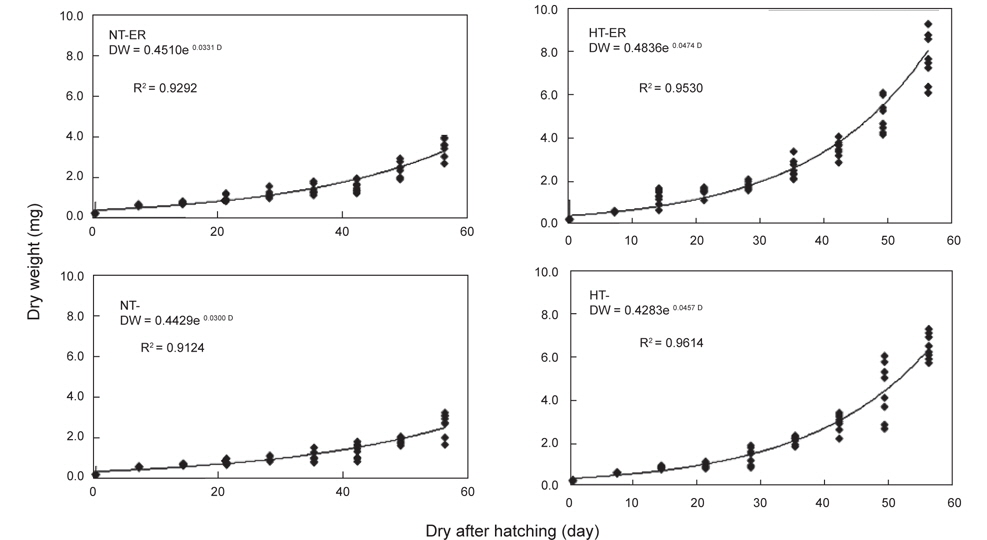
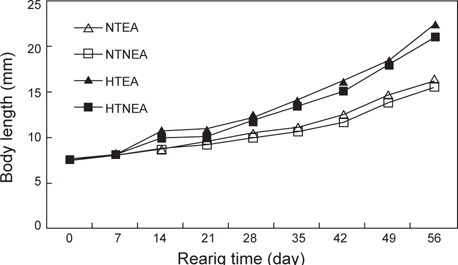
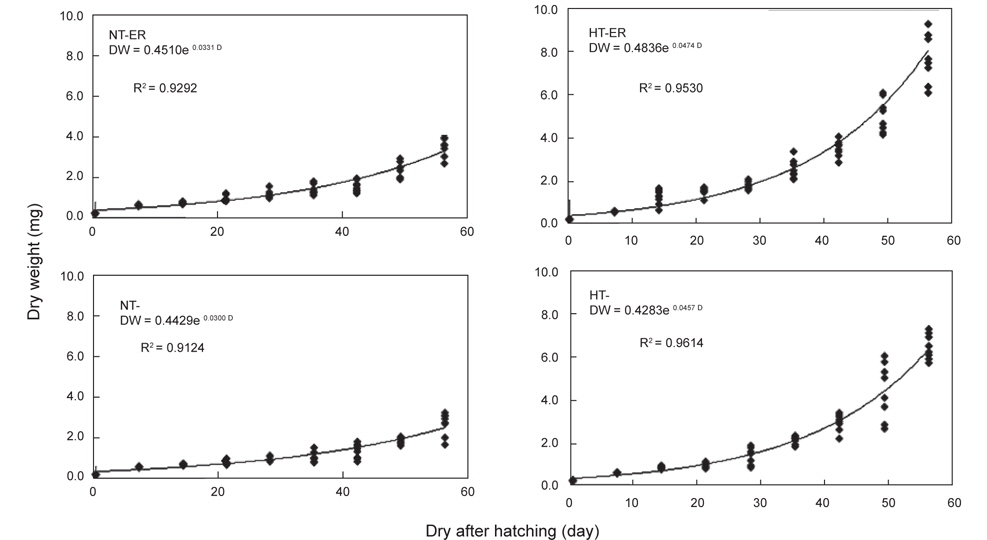
쥐노래미 자치어의 체장성장은 전체적으로 부화 후 21일까지는 점진적으로 증가하다가 28일 이후에는 빠른 성장을 보였다. 실험 시작 시 평균체장은 7.619 mm이였으며, 실험 56일 후 NT구의 EA구는 16.40 mm, NEA구는 15.54 mm이였고, HT구의 EA구는 22.50 mm, NEA구는 21.09 mm로 나타났다. 위의 결과로부터 먹이별 성장은 큰 차이를 보이지 않았으나 수온에 따른 자치어 성장은 뚜렷하게 차이를 보였다(Fig. 1). 사육기간 동안의 체중성장을 살펴보면, NT구의 NEA를 공급한 실험구에서 사육기간 동안에 지수적인 증가{DW=0.4429e0.03t (R2=0.9124)}를 보였다. EA구에서도 사육기간 동안 체성장에 지수적인 증가{DW=0.451e0.0331t (R2=0.9292)}를 보였다 (Fig. 2). HT구의 체중성장을 살펴보면, NEA구에서 사육기간 동안에 지수적인 증가{DW=0.4283e0.0467t (R2=0.9614)}를 보였다. EA구에서도 사육기간(56일) 동안 지수적인 증가{DW=0.4836e0.0474t (R2=0.953)}를 보였다.
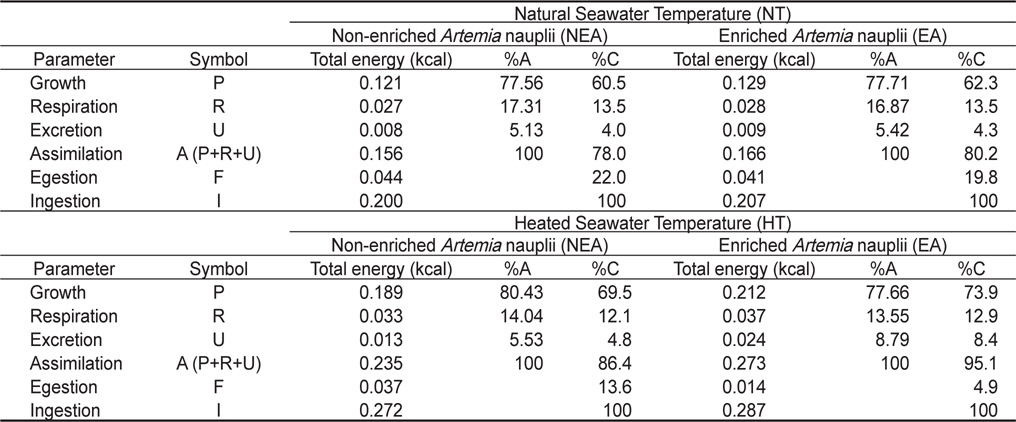
쥐노래미 자치어의 체중성장에 사용된 에너지는 Table 1에 나타낸 생체에너지 함량(cal/g dry weight)으로 부터 산출하였다. NT구의 NEA구에서 사육기간 동안 실험종료 시의 생체에너지 (5,593.30 cal)에서 실험시작 시의 생체에너지(5,471.64 cal)를 뺀 값 즉, 121.66 cal의 에너지를 체중성장에 사용하였고, EA구에서는 실험종료시의 생체에너지(5,600.45 cal)에서 실험시작시의 생체에너지(5,471.64 cal)를 뺀 값 즉, 128.81 cal의 에너지를 체중성장에 사용하였다. 또한 HT구의 NEA구에서 사육기간 동안 실험종료 시의 생체에너지(5,660.98 cal)에서 실험시작 시의 생체에너지(5471.64 cal)를 뺀 값 즉, 189.34 cal의 에너지를 체중성장에 사용하였고, EA구에서는 실험종료시의 생체에너지(5,683.96 cal)에서 실험시작시의 생체에너지(5,471.64 cal)를 뺀 값 즉, 222.32 cal의 에너지를 체중성장에 사용하였다.

Energy contents of a Hexagrammos otakii larvae and juveniles fed the different diets {non-enriched Artemia nauplii (NEA) and enriched Artemia nauplii (EA)} and temperatures {natural seawater temperature (NT) (5.1-8.5℃) heated seawater temperature (HT) (13.0±0.5℃)}
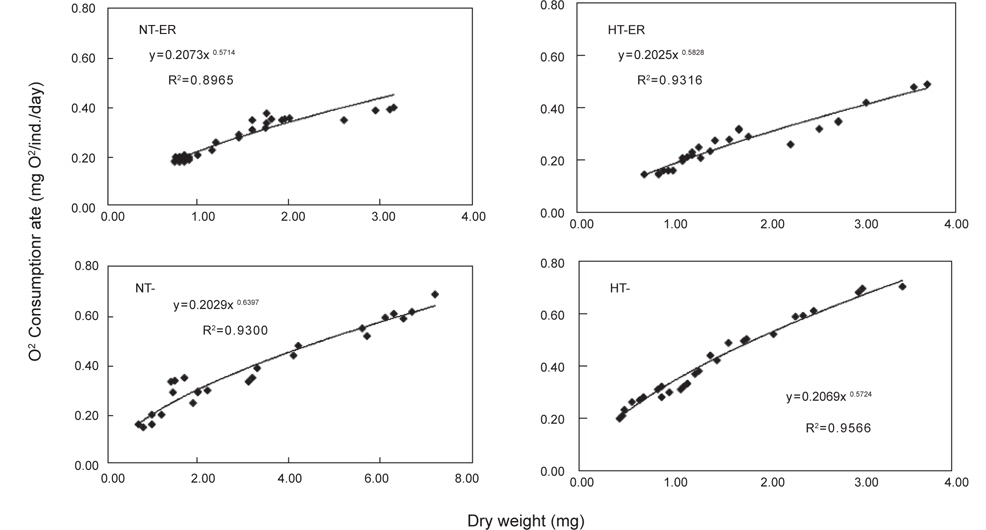
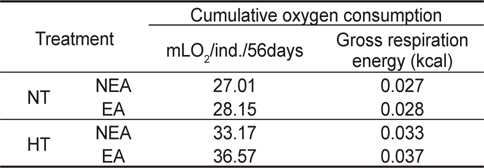
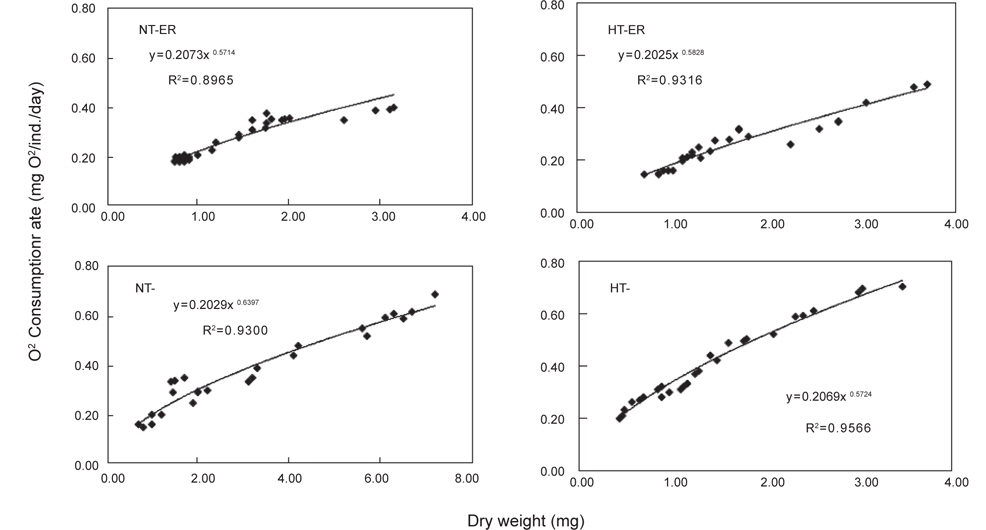
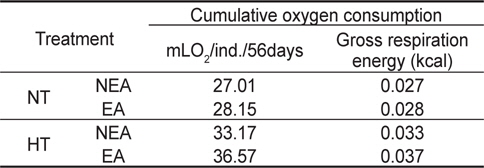
쥐노래미 자치어에 있어 온도별, 먹이별 산소소비율(mL O2/ ind./day)경향을 살펴보면, 사육기간 동안에 성장과 더불어 계속 증가하였다. NT구에서 NEA구와 EA구의 자치어 산소소비 경향을 살펴보면, 건조중량이 증가함에 따라 누승적으로 증가 하였다(Fig. 3). NT구에서 NEA구의 산소소비율(mg O2/ind./day)은 Y=0.2073x0.6714 (R2=0.8965)로 나타낼 수가 있었으며, EA구의 산소소비율은 Y=0.2029x0.6379 (R2=0.93)로 나타났다 (Fig. 3). 따라서 성장곡선과 산소소비율을 적분구간(56일)동안의 적분에 의해 먹이별로 각각 27.01 mg O2/ind. 및 28.15 mg O2/ind.를 소비하였으며, 호흡에 사용된 대사에너지는 각각 0.027 kcal/ind.와 0.028 kcal/ind.였다(Table 2). HT구에서 NEA구의 산소소비율(mg O2/ind./day)은 Y=0.2025x0.5828 (R2=0.9316)로 나타났고, EA구의 산소소비율은 Y=0.2069x0.5724 (R2=0.9566)로 나타났으며, 건조중량이 증가함에 따라 누승적으로 증가하였다(Fig. 3). 따라서 성장곡선과 산소소비율를 적분구간(56일)동안의 적분에 의해 먹이별로 각각 33.17 mg O2/ind. 및 36.57 mg O2/ind.를 소비하였으며, 산소소비에 사용된 에너지는 각각 0.033 kcal/ind.와 0.037 kcal/ind.였다(Table 2).
Cumulative oxygen consumption rate and maintenance energy of H. otakii larvae and juveniles fed the different diets {non-enriched Artemia nauplii (NEA) and enriched Artemia nauplii (EA)} and temperatures {natural seawater temperature (NT) (5.1-8.5℃) and heated seawater temperature (HT) (13.0±0.5℃)}
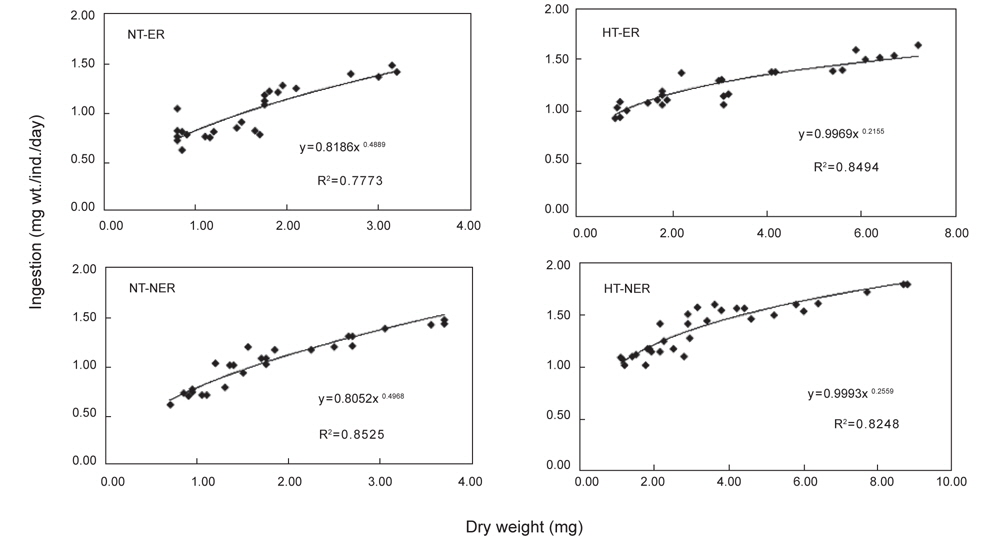
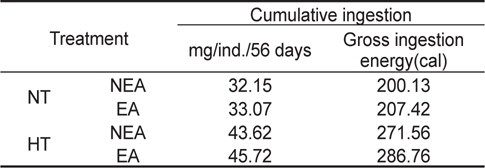
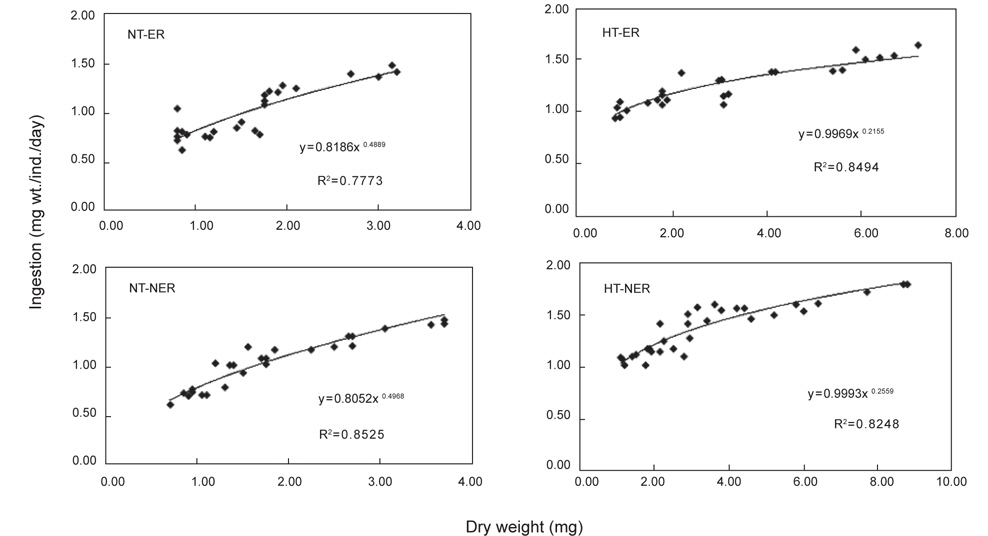
NT구에서 섭취경향을 살펴보면 일간 섭취량은 누승적으로 증가하였으며, 자치어의 건조중량과 일간 섭취량간의 관계는 NEA구와 EA구에서 각각 Y=0.8186x0.4889 (R2=0.7773), Y=0.8052x0.4968 (R2=0.8525)로 나타났다(Fig. 4). 성장곡선과 섭취율을 적분구간(56일)동안의 적분에 의해 NEA구와 EA구에서 각각 32.15 mg, 33.07 mg을 섭취하였으며, 섭취에 사용된 에너지는 각각 200.13 cal, 207.42 cal 였다(Table 3). HT구에서 자치어의 건조중량과 일간 섭취량 관계는 먹이별로 NEA구와 EA구에서 각각 Y=0.9969x0.2155 (R2=0.4894), Y=0.9993x0.2559 (R2=0.8248)로 나타났으며 건조중량이 증가함에 따라 누승적으로 증가하였다(Fig. 4). 성장곡선과 섭취율을 적분구간(56일)동안의 적분에 의해 NEA구와 EA구에서 각각 43.62 mg/ind 45.72 mg/ind.을 섭취하였으며, 섭취에 사용된 에너지는 NEA구와 EA구에서 각각 271.56 cal/ind., 286.76 cal/ind.이였다 (Table 4).
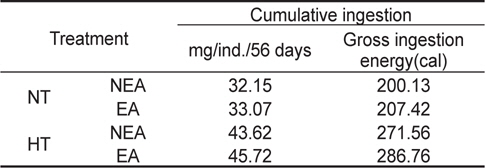
Cumulative ingestion rate and ingestion energy of H. otakii larvae and juveniles fed the different diets {non-enriched Artemia nauplii (NEA) and enriched Artemia nauplii (EA)} and temperatures {natural seawater temperature (NT) (5.1-8.5℃) and heated seawater temperature (HT) (13.0±0.5℃)}
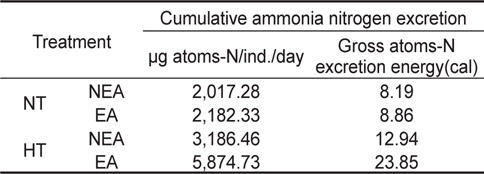
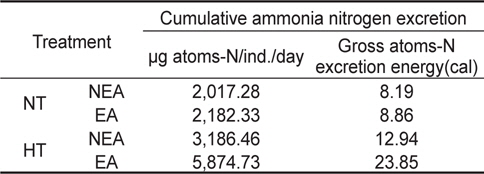
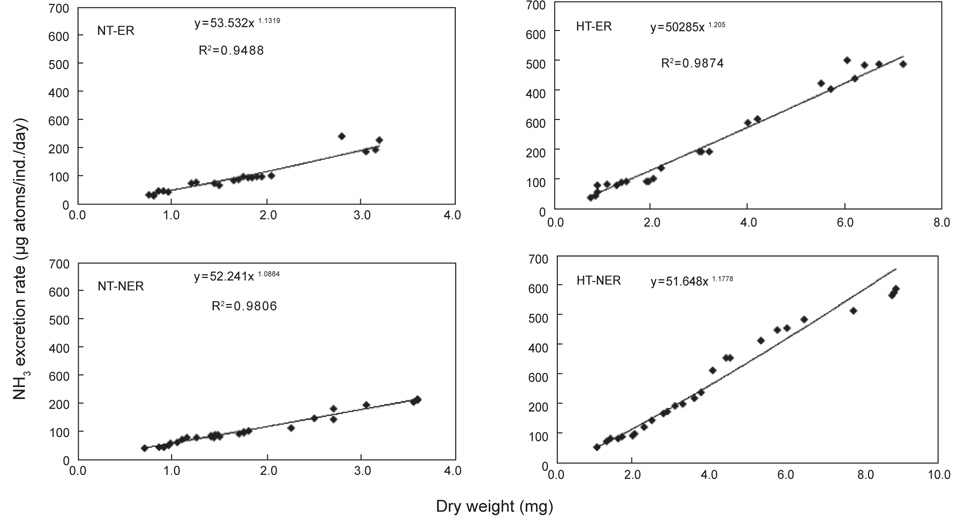
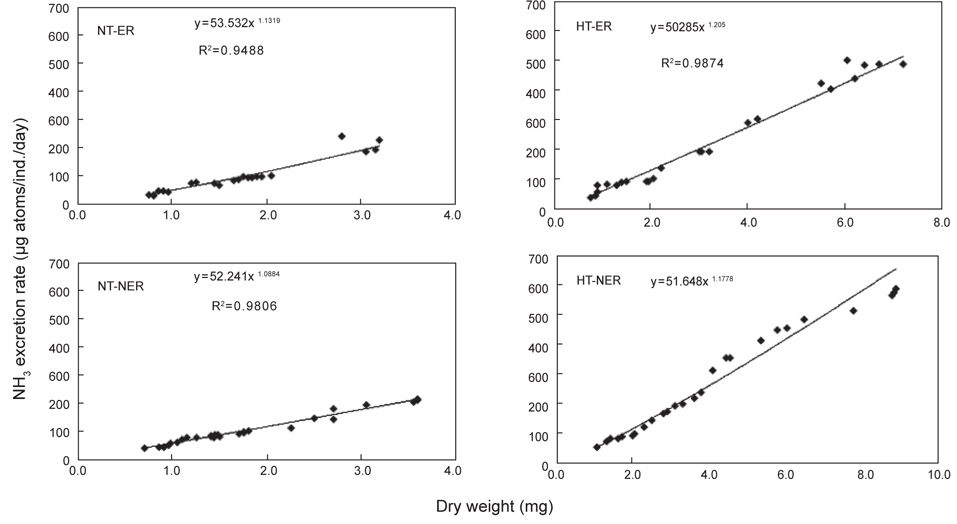
Cumulative ammonia nitrogen excretion rate and excretion energy of H. otakii larvae and juveniles fed the different diets {non-enriched Artemia nauplii (NEA) and enriched Artemia nauplii (EA)} and temperatures {natural seawater temperature (NT) (5.1-8.5℃) and heated seawater temperature (HT) (13.0±0.5℃)}
쥐노래미 자치어의 뇨배설을 보면 서로 다른 수온구와 먹이구 간에 모두 체중이 증가함에 따라 비례적으로 증가하였다. NT구에서 뇨배설 경향을 살펴보면, 일간 뇨배설량은 누승적으로 증가하였으며, 자치어의 건조중량과 일간 뇨배설량과의 관계는 먹이별로 NEA구와 EA구에서 각각 Y=53.532x1.1319 (R2=0.9488) 및 Y=52.241x1.0884 (R2=0.9806)로 나타났다(Fig. 5). 성장곡선과 뇨배설율을 적분구간(56일)동안의 적분에 의해 NEA구와 EA구에서 각각 2,017.28 μg/ind. 및 2,182.33 μg/ind.을 배설하였으며, 뇨배설에 사용된 에너지는 각각 8.19 cal/ind. 및 8.86 cal/ind.였다(Table 4). HT구에서 뇨배설 경향을 살펴보면, 일간 뇨배설량은 누승적으로 증가하였으며, 자치어의 건조중량과 일간 뇨배설량과의 관계는 먹이별로 NEA구와 EA구에서 각각 Y=50.285x1.206 (R2=0.9874) 및 Y=51.648x1.1778 (R2=0.9831)로 나타났다(Fig. 5). 성장곡선과 뇨배설율을 적분구간(56일)동안의 적분에 의해 NEA구와 EA구에서 각각 3,186.46 μg/ind. 및 5,874.73 μg/ind.을 배설하였으며, 뇨배설에 사용된 에너지는 각각 12.94 cal/ind. 및 23.85 cal/ind.이였다(Table 4).
쥐노래미 자치어의 실험사육기간 56일 동안 수온별(NT구와 HT구), 먹이별(NEA구와 EA구)의 누적 에너지 수지를 Table 5에 나타내었다. NT구에서 NEA구의 경우 체성장에 사용된 에너지가 0.121 kcal, 호흡에 사용된 누적 에너지는 0.027 kcal, 총 섭취한 누적에너지는 0.200 kcal로서 총 섭취에너지에 대해 성장은 60.5%, 호흡은 13.5%를 차지하였다. 또한 분배출 및 뇨배설에 사용된 에너지는 각각 0.044 kcal와 0.008 kcal로 총 섭취 에너지에 대해 각각 22% 및 4%를 차지하였다. NT구에서 EA구의 경우 체성장에 사용된 에너지가 0.129 kcal, 호흡에 사용된 누적 에너지는 0.028 kcal, 총 섭취한 누적에너지는 0.207 kcal로서 총 섭취에너지에 대해 성장은 62.3%, 호흡은 13.5% 를 차지하였다. 또한 분배출 및 뇨배설에 사용된 에너지는 각각 0.041 kcal, 0.009 kcal로서 총 섭취에너지에 대해 각각 19.8%, 4.3%를 차지하였다. HT구의 경우 NEA구에서 체성장에 사용된 에너지가 0.189 kcal, 호흡에 사용된 누적 에너지는 0.033 kcal, 총 섭취한 누적에너지는 0.272 kcal로서 총 섭취에너지에 대해 성장은 69.5%, 호흡은 12.1%를 차지하였다. 또한 분배출 및 뇨배설에 사용된 에너지는 각각 0.037 kcal, 0.013 kcal로서 총 섭취에너지에 대해 각각 13.6%, 4.8%를 차지하였다. HT구의 경우 EA구에서 체성장에 사용된 에너지가 0.212 kcal, 호흡에 사용된 누적 에너지는 0.037 kcal, 총섭취한 누적에너지는 0.287 kcal로서 총 섭취에너지에 대해 성장은 73.9%, 호흡은 12.9%를 차지하였다. 또한 분배출 및 뇨배설에 사용된 에너지는 각각 0.014 kcal와 0.024 kcal로서 총 섭취에너지에 대해 각각 4.9% 및 8.4%를 차지하였다.
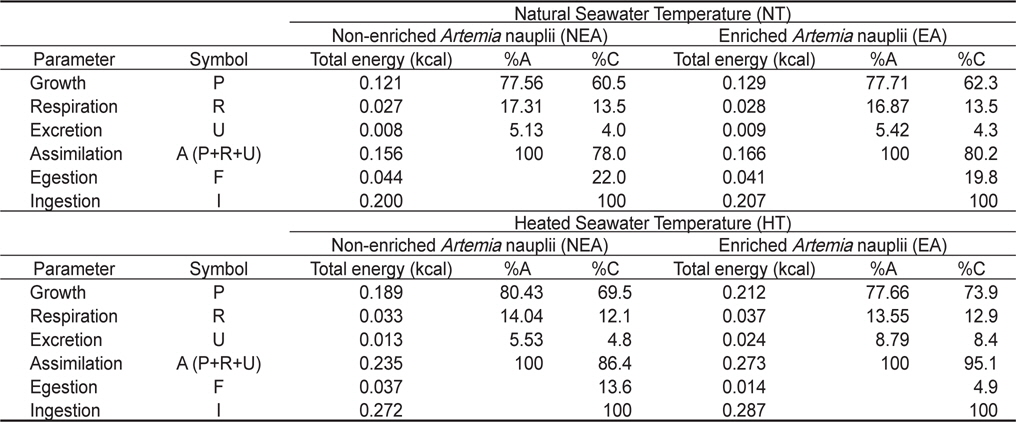
Cumulative energy budget of the H. otakii larvae and juveniles fed the different diets {non-enriched Artemia nauplii (NEA) and enriched Artemia nauplii (EA)} and temperatures {natural seawater temperature (NT) (5.1-8.5℃) and heated seawater temperature (HT) (13.0±0.5℃)}
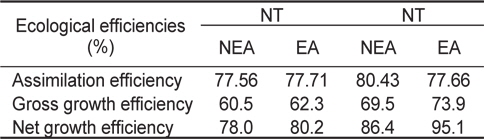
쥐노래미 자치어의 섭취, 성장, 호흡, 뇨배설 및 분배출을 측정한 자료로부터 생태효율을 구하였다(Table 6). NT구에서 자치어의 동화효율은 먹이별로 각각 77.56%, 77.71%로 EA구가 NEA구에 비해 높은 값을 나타냈으며, 총 성장효율(K1)은 각각 60.5%, 62.3%, 순 성장효율(K2)은 78%, 80.2%로 나타났다. HT구에서 자치어의 동화효율은 먹이별로 각각 80.43%, 77.66%로 NEA구가 EA구에 비해 높은 값을 나타냈으며, 총 성장효율(K1)은 각각 69.5%, 73.9%, 순 성장효율(K2)은 86.4%, 95.1%로 나타났다.
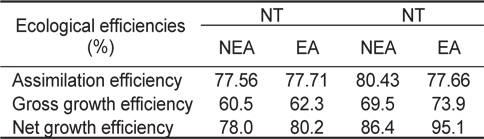
Ecological efficiencies of H. otakii larvae and juveniles fed the different diets {non-enriched Artemia nauplii (NEA) and enriched Artemia nauplii (EA)} and temperatures {natural seawater temperature (NT) (5.1-8.5℃) and heated seawater temperature (HT) (13.0±0.5℃)}
해산동물의 에너지흐름을 파악하기 위해서 에너지수지를 규명하기 전에 그 생물의 성장과 생활사를 정확하게 조사하는 것이 필요하다. 또한 일반적으로 에너지는 생명체의 생존에 가장 기본적으로 요구되므로 먹이를 선택함에 있어 우선적으로 고려해야 할 것이 먹이의 에너지 함량이다. 동일한 어종에 있어서도 먹이, 실험 디자인, 사육방법 및 환경적 요인에 의해 달라질 수 있다고 보고되었다(Keembiyehetty and Wilsom, 1997).
본 실험에 사용된 먹이
해산어류의 성장은 일반적으로 지수적인 성장을 나타낸다. 쥐노래미 자치어의 수온별, 먹이별 성장 역시 전체적으로 지수적인 성장 경향을 나타내었다. 자치어의 부화 직후 평균체장은 7.619 mm이었으며 실험 56일 후 자연수온구(NT구)의 영양강화한
쥐노래미 자치어의 에너지수준에 따른 개체 중량과 산소소비량은 자연수온(NT구)에서 갓 부화한
위의 결과로 미루어 볼 때 자치어의 산소소비량은 먹이에 대한 영향보다는 수온에 따른 대사활동에 의한 차이로 사료된다. 동물의 대사작용과 온도는 밀접한 관계가 있다. 효소 반응은 온도에 따라 변하므로 체온이 낮으면 대사율이 저하되기 때문이다. 또한 산소소비량은 어종 및 크기에 따라 다르게 나타나거나 어류의 생태적인 특징(생활사)에 따라 다소 차이가 있는 것으로 사료되며, 아울러 자치어의 대사율은 수온, 염분 및 먹이의 밀도 등의 환경 요인에 따라 영향을 받는다고 보고되었다 (Almatar, 1984).
쥐노래미 자치어의 수온별, 먹이별 섭취량은 전체적으로 누승적인 증가 경향을 보였으며 건조중량이 증가함에 따라 일간 섭취량은 계속적인 증가를 나타내었다. 실험 기간 동안의 자치어의 섭취량에 대한 결과 또한 수온에 대한 영향이 컸으며 대사활동에 의한 소화와 흡수의 관계에서 기인된 것이라 사료된다.
해양생물에 있어 암모니아 분비는 단백질 이화작용(소화)에 의한 배설의 마지막 산물이며, 생물의 영양학적인 상태, 재생산적인 상태와 함께 다양한 단백질 이화작용의 지시자로 간주되고 있다(Griffiths and Griffiths, 1987). 그러나 쥐노래미 자치어의 수온별, 먹이별 뇨와 분으로 소비된 에너지 비율은 다른 생리작용의 에너지보다 낮은 비중을 차지하기 때문에 에너지수지에 미치는 영향은 크지 않을 것으로 사료된다. Calow (1985)은 먹이의 동화효율로 먹이의 밀도와 성분 및 크기가 치어기 어류의 에너지 이용에 영향을 끼친다고 보고하였고, 또한 일반적으로 초식성 어류는 섭취한 먹이의 동화효율이 낮아 분배출을 통한 에너지의 상실이 많으므로 총 성장효율이 낮게 나타나지만 (Bighead carp의 경우 37-49%, Opuszynski et al., 1991), 는 반면에 육식성 어류는 단백질 함량이 높은 먹이를 섭취하므로 질소배설을 통한 에너지 상실량이 초식성 어류에 비해 많으므로 동화효율이 높게 나타난다(틸라피아의 경우 61.4-80.0%, Lu et al., 2004)고 보고되었다. 본 실험에서는 가온해수에서 갓 부화한
이상과 같은 에너지수지에 대한 성장률, 섭취량, 소화효율 등의 자료는 종묘생산 관점에서 매우 중요하다. 에너지흐름에 대한 수치는 각각의 종마다 다르지만 수온과 먹이 및 그 밖의 사육환경에 따른 발육단계의 수치변화를 파악할 수 있으며 양식대상종의 생산성 향상 및 안정적인 종묘생산 라인이 구축 될 수 있을 것이다.