Flathead flounder
홍가자미(
홍가자미에 대한 연구로는 외국의 경우 분포와 이동, 계절적 분포(Yokohama et al., 1990; Vdovin, 2004), 연령·성장 및 성장양식(Yagishita et al., 2006; Iwakawa et al., 2013), 푼카만의 연급군 연변동 원인(Nakatani et al., 2002), 산란장 특성, 성숙상태와 산란, 성어의 생활사 및 성숙체장의 소형화(Ishida and Kitakata, 1982; Yokohama et al., 1991; Hirose and Minami, 2007; Yamada et al., 2008; Fujiwara et al., 2009), 난자치어의 분포 및 식성(Miyamoto, 1993) 등 많은 연구가 있었지만, 우리나라의 경우 동해안 가자미 연구는 용가자미의 성숙과 산란(Choe et al., 1999), 참가자미의 연령과 성장 및 성숙과 산란(Lee et al., 2006; Cha et al., 2006), 기름가자미의 성숙과 산란 (Cha et al., 2008) 등이 있을 뿐 홍가자미에 관한 연구는 아직 이루어 지지 않았다.
최근 기후변화 등에 의한 해양환경의 변화로 우리나라 동해안에 분포하는 가자미류는 종수가 현저히 줄어들었고 자원수준도 많이 감소하였으며, 특히 홍가자미는 동해안 일원에서만 한정적으로 서식하고 있어 자원의 보호와 회복이 시급한 실정에 있다. 본 연구는 우리나라 동해안 홍가자미의 성숙 및 산란과정을 관찰하고 산란시기, 포란수 및 성숙체장 등의 자원생태학적 특성치를 분석하여 어획크기 제한과 어획기준 등 자원관리를 위한 기초자료를 얻고자 시도하였다.
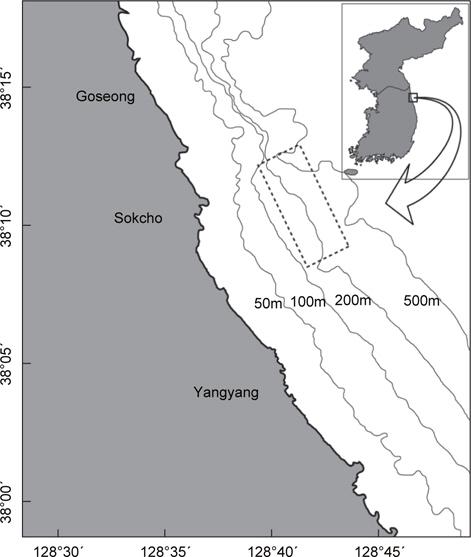
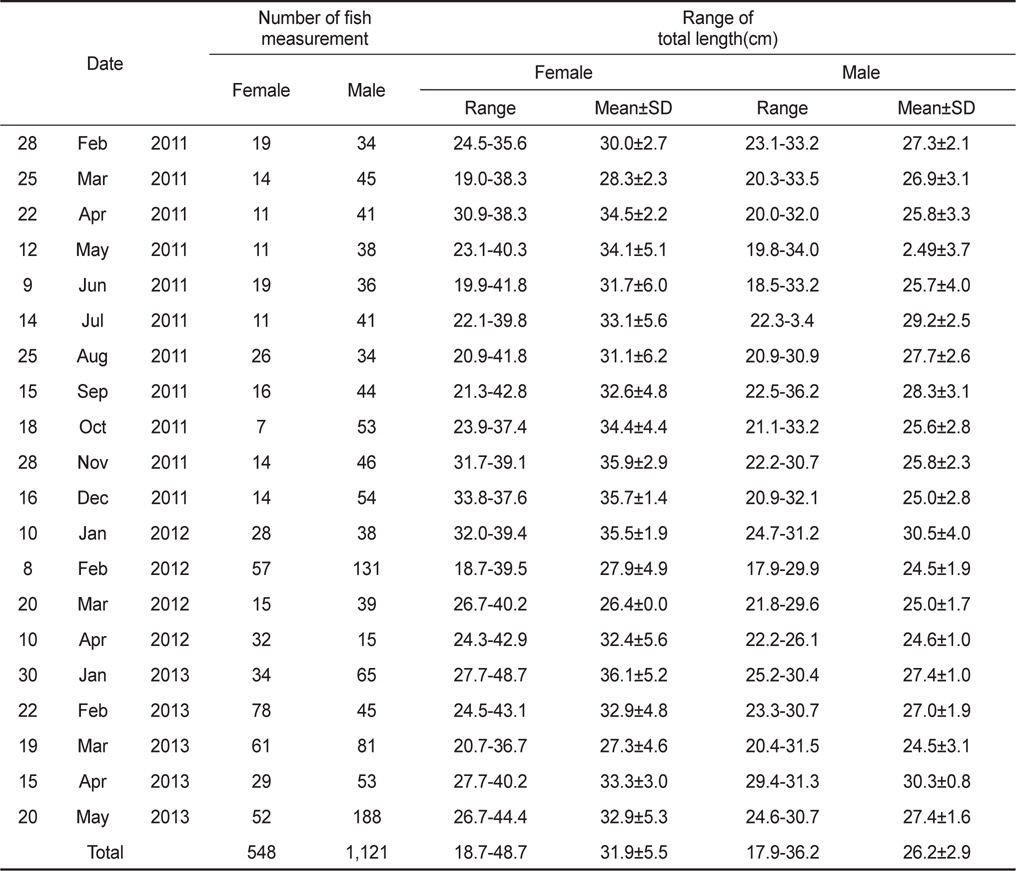
실험에 이용된 홍가자미는 2011년 2월부터 2013년 4월까지 강원연안 수심 150-400 m부근에서 상업적인 자망어선으로 조업한 어획물을 위판장에서 구득한 것이다(Fig. 1, Table 1).
홍가자미에 대한 생물학적 측정으로 어체의 길이는 체장 (Body length, BL)과 전장(Total length, TL)을 0.1 mm, 전중(Total weight, TW)은 0.1 g, 생식소 중량(Gonad weight, GW)은 0.01 g 단위까지 계측하였다. 홍가자미 암컷 및 수컷의 생식소 중량의 월별 변화를 통한 산란시기를 추정하기 위한 생식소 중량지수(Gonad Index, GI)는 전중에 대한 생식소 중량 비율로 구하였다. GI = GW/TW×100
생식소의 성숙단계를 판단하기 위하여 육안관찰과 조직학적 방법을 병행하여 조사하였다. 육안관찰의 방법으로 암컷 난소의 경우, 미숙기(Immature)는 생식소가 작고 육안으로 난립을 확인하기 어려운 상태, 중숙기(Maturing)는 생식소 부피와 난립이 커졌지만 투명란은 보이지 않는 상태, 성숙기(Mature)는 생식소의 부피가 최대이고 투명란이 나타나기 시작하는 상태, 방중기(Spawning)는 복부에서 투명란이 유출되어 생식소 부피가 줄어드는 상태, 방후기(Spent)는 생식소가 현저히 줄어들고 암자색 또는 적자색으로 변하게 되는 상태로 구분하였다. 수컷 정소의 경우, 미숙기(Immature)는 정소가 반투명한 연분홍색을 나타내는 상태, 성숙기(Mature)는 정소가 유백색으로 변화되며 유상정액은 흐르지 않는 상태, 방중기(Spawning)는 복부를 압박하면 유상정액이 유출되는 상태, 방후기(Spent)는 정소가 축소되고 어두운 회색으로 변하게 되는 상태를 기준으로 하였다(NFRDI, 1985; Wenner et al., 1990).
난소의 발달과정을 조직학적으로 관찰하기 위해 생식소 중 일부는 Bouin용액에 24시간 고정하고 탈수하여 paraffi ne 블록제작을 한 후, 마이크로톰 절단에 따라 4-6 μm의 두께로 연속절편을 만들어서 Mayer's Haematoxyline과 Eosin으로 염색하여 현미경을 사용하여 관찰하였다.
포란수와 난경을 측정하기 위해 생식소 일부를 절취하여 중량을 재고, Gilson용액에 보존한 후 관찰 시에는 희석용액 중 0.2 mL씩 취하여 만능투영기(Nikon V-16E)로 50배의 배율로써 난립수와 난경을 측정하였고 난경의 월별조성 및 모드의 변화를 관찰하였다.
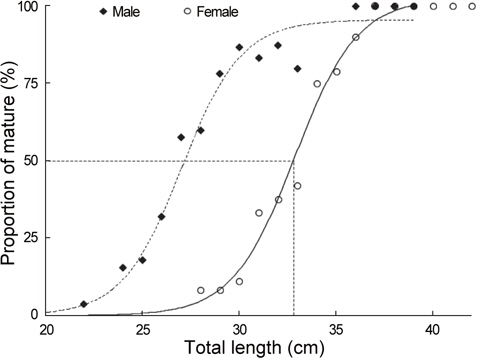
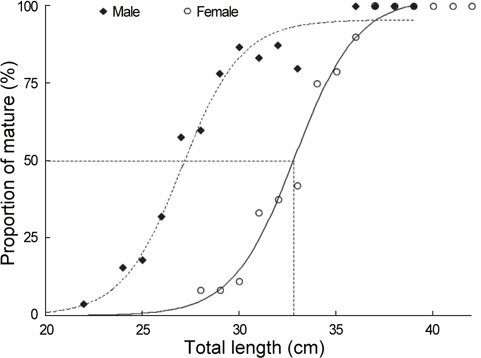
산란참여전장을 알기 위하여 군성숙도는 산란기간을 중심으로 전장을1 cm단위로 구분하여 전체 출현개체수에 대한 성숙 개체의 비율로서 구하였으며, 전장별 성숙비율(
a : 최대 성숙비율, b : 상수, χ : 전장, χ0: 50% 성숙전장
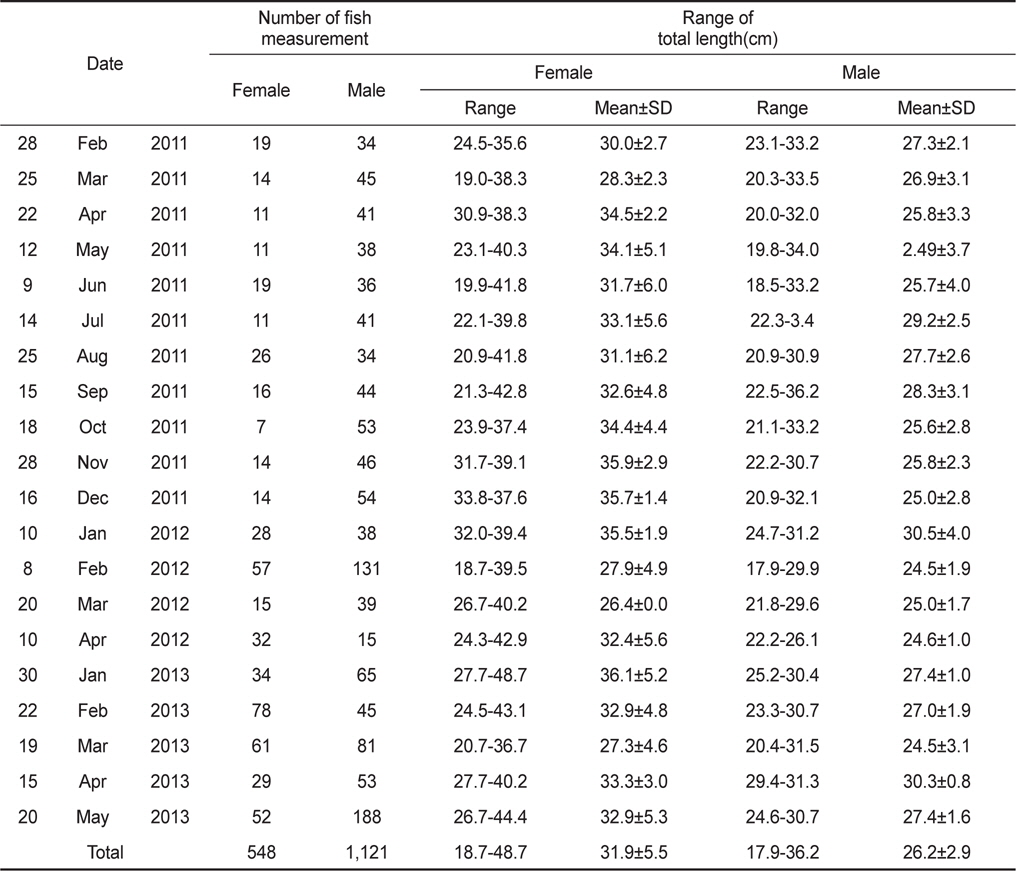
조사해역에서 채집한 홍가자미의 전장범위는 암수 각각 평균 31.9±5.5 (18.7-48.7)cm, 26.2±2.9 (17.9-36.2)cm로 나타났으며 전체 암수의 성비는 암컷 32.8%, 수컷 67.2%로서 수컷이 2배 이상 많았다(Table 1). 산란시기의 전장조성과 성비조성의 차이를 분석하기 위하여 산란시기(1-4월)와 비산란시기(5-12월)로 구분하여 통계검정을 실시한 결과, 평균전장은 암컷과 수컷 모두 산란시기가 비산란시기보다 작게 나타났다(Mann-Whitney
Sampling data of flathead flounder Hippogloides dubius off Gangwon Province, East Sea of Korea
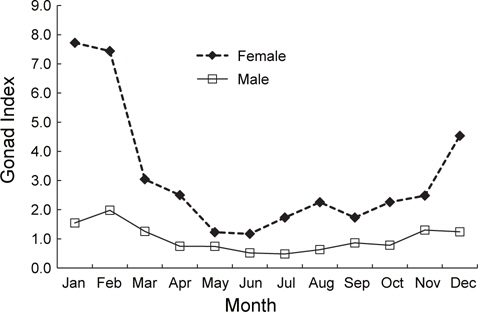
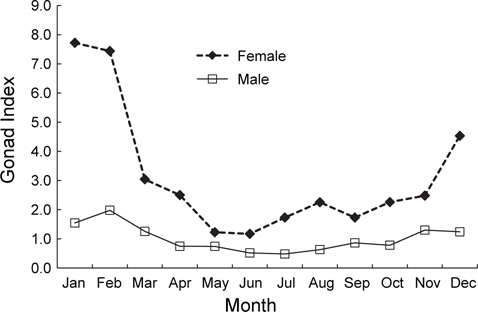
2011년 2월부터 2013년 4월까지 홍가자미 암컷과 수컷의 생식소 숙도지수의 월 변화를 분석하였다(Fig. 2). 홍가자미 암컷 생식소 숙도지수의 월 변화를 보면 1월과 2월에 평균 7.72, 7.44로 가장 높았으며 3월에 평균 3.04로 급격히 떨어졌다. 4월부터 11월까지 평균 1.17-2.50의 범위에서 변동을 보였으며 12월에 평균 4.53으로 증가하기 시작하였다. 홍가자미 수컷 생식소 숙도지수의 월 변화를 보면 2월에 평균 1.97로 가장 높았으며 3월에 평균 1.25로 감소하였다. 4월부터 10월 동안은 0.48-0.86의 범위에서 변동을 보였으며 11월부터 2월까지 점차 증가하는 것으로 나타났다. 따라서 월별 난소 및 정소의 생식소 숙도지수는 2월부터 4월 사이에 크게 떨어지는 것으로 나타났으며 2월에 가장 높은 값을 보이고 3월부터 숙도지수가 급격히 감소하는 것으로 보아 주 산란시기는 2-3월로 추정되었다.
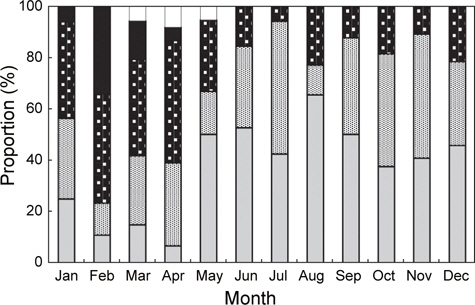
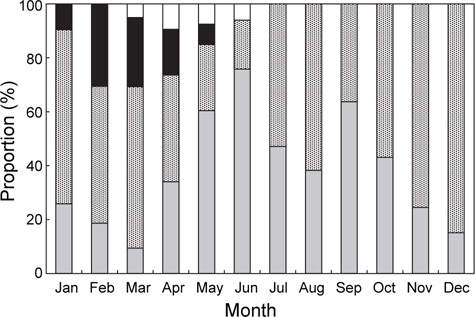
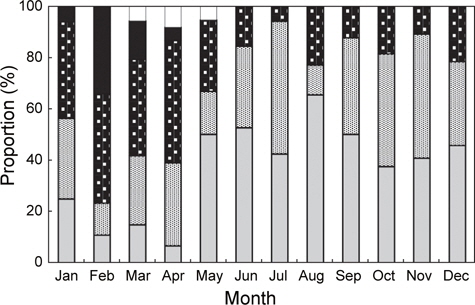
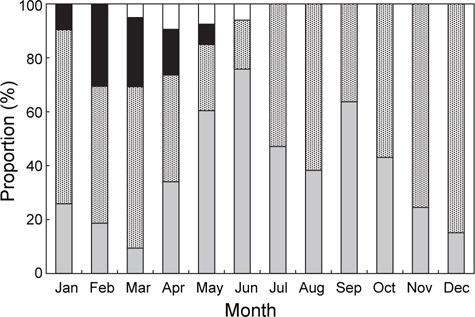
홍가자미의 월별 성숙비율을 알아보기 위해 전장 30 cm이상의 개체에 대한 성숙단계별 월별 출현빈도를 관찰하였으며 성숙도 구분은 미숙기, 중숙기, 성숙기, 방란기 및 방후기의 5단계로 분류하였다. 암컷은 미숙기, 중숙기 및 성숙기 단계를 보이는 개체는 연중 출현하는 경향을 보였는데 각각의 성숙단계별 출현비율을 살펴보면, 미숙단계 개체의 출현범위와 평균은 7-65%(평균 37%), 중숙단계는 12-52%(평균 31%), 성숙단계는 6-47%(평균 25%)로서 나타났다. 반면 방란중인 개체는 1월부터 4월까지 5-34%(평균 15%)로서 2월에 34%로 가장 높았으며, 방란후 개체는 3-5월에 출현하였고 범위와 평균은 6-8%(평균 7%)이었다(Fig. 3).
수컷의 연중 성숙단계별 출현비율을 보면 미숙과 성숙단계에 해당하는 개체는 연중 출현하였다. 미숙단계 개체의 출현범위와 평균은 9-76%(평균 38%), 성숙단계는 18-85%(평균 52%)이었고 방정중인 개체는 1월부터 5월까지 출현하였고, 2월과 3월에 각각 31%, 26%로 가장 높았다. 방후기 개체는 3월부터 6월에 출현하였고 출현범위와 평균은 5-9% (평균 7%)로 나타났다(Fig. 4).
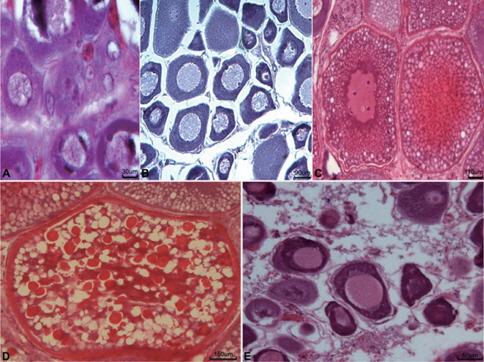
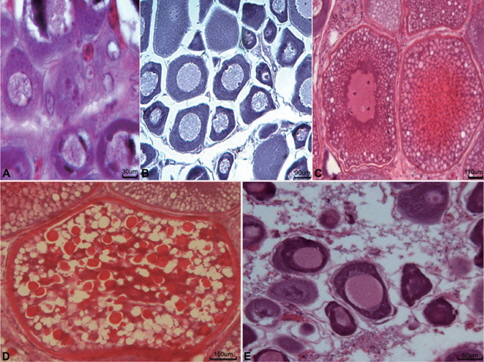
5-7월의 개체들에서는 난소가 활성화되기 시작하여 초기성장기의 난소소엽내에서는 난원세포(난경 3-10 μm)와 초기 난모세포(난경 20-100 μm)들이 출현하였다(Fig. 5A).
8-10월에 접어들면 대부분의 개체가 성장기의 난소를 갖는데, 난소내에서는 주변인기 난모세포(난경 80-300 μm)들이 난소 소엽을 채웠다(Fig. 5B).
11-1월에는 난소의 발달이 급격히 진행되어 대부분의 난소내에는 난황포기 및 난황과립을 갖는 난모세포와 다량의 난황이 축적된 난모세포(난경 250-800 μm)들이 출현한다(Fig. 5C).
2월과 3월에는 완숙기의 난소가 출현하였고, 이때 암컷의 난소소엽내에는 난경 700-1,300 μm 전후의 완숙난으로 가득 채워진다(Fig. 5D).
4월 이후 방란이 끝난 암컷개체들의 난소소엽내에는 미방출 된 난모세포들과 잔존여포들이 회복·흡수 되었으며 난소소엽은 위축되었다(Fig. 5E).
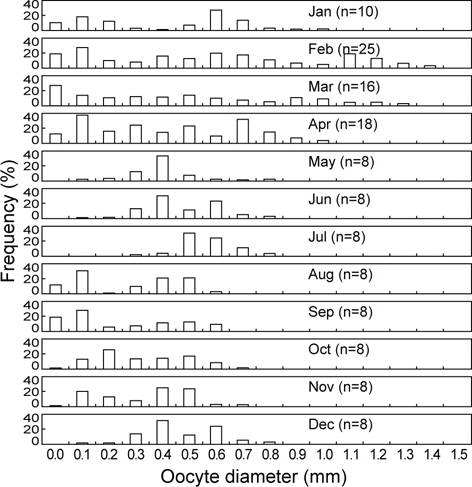
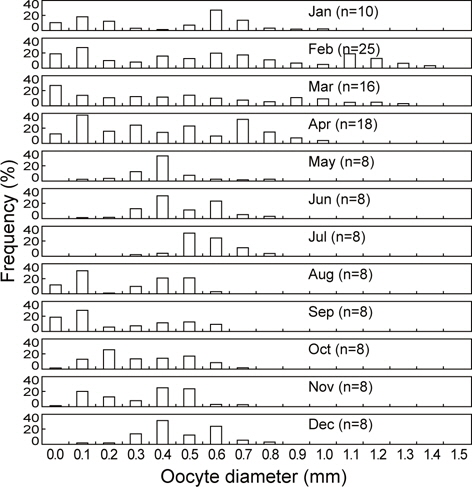
2011년 2월부터 2013년 4월까지 강원연안에 분포하는 107마리의 성숙개체에 대한 월별 난소내 난모세포의 난경조성을 분석하였다. 1월에는 난경모드 0.1 mm를 중심으로 하는 소형난경군(Small size group)과 난경모드 0.6 mm를 중심으로 중형난경군(Middle size group)으로 2개의 그룹을 형성하였으며 관찰된 가장 큰 난모세포는 난경 1.0 mm이었다. 2월에는 1월처럼 난경모드 0.1 mm의 소형난경군과 난경모드 0.6 mm의 중형난경군과 함께 산란이 진행중인 난경모드 1.1 mm의 대형난경군(Large size group)으로 3개의 그룹을 형성하였으며 최대 난경 크기는 1.4 mm이었다. 3월에는 난경모드 0.1 mm미만의 소형 난경군과 0.5 mm의 중형난경군과 난경모드 0.9 mm이상의 난경군에서 산란이 진행 중에 있었다. 4월에는 난경 0.1 mm의 소형난경군과 난경 0.7 mm의 중형난경군에서 주 모드를 보이고 있으며, 난경 0.9-1.0 mm이상의 대형난경군(Large size group)의 조성비율은 2월과 3월에 비해 감소한 것으로 나타났다.
5월부터 7월까지는 난경 0.4-0.6 mm의 범위에서 주 모드를 형성하고 있으며 대형난경군은 거의 관찰되지 않았으며 난경0.1 mm이하의 소형난경군도 출현하지 않았다. 8월과 11월까지는 주 모드가 난경 0.1-0.2 mm와 난경 0.4-0.6 mm인 소형 및 중형난경군이 주축을 이루며 난경 0.8 mm이상의 대형난경군은 나타나지 않았다. 12월에는 난경이 0.3-0.6 mm인 중형난경군이 주축을 이루고 있었으며 최대난경은 0.8 mm로 나타났다(Fig. 6).
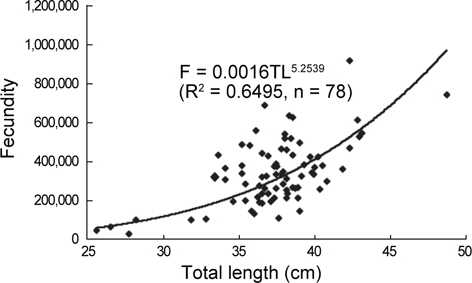
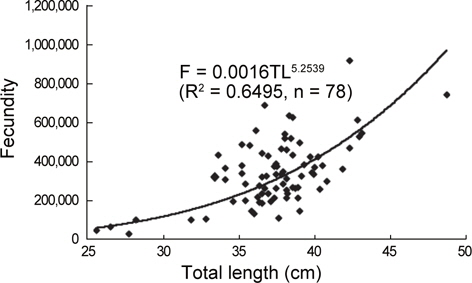
홍가자미 난소내의 포란수를 관찰한 결과, 전장 26.0-48.7 cm 범위에 대한 포란수는 27,372-915,209립으로 각 전장체급별로는 체급 25-30 cm의 평균포란수와 표준편차는 59,536±31,297립, 체급 31-35 cm에서는 259,984±128,861립, 체급 36-40 cm에서는 339,582±133,711립, 체급 40 cm이상에서는 502,971±196,807 립으로 나타났으며 전장(TL)과 포란수(F)와의 관계는
F=0.0016TL5.2539 (R2 = 0.6495)
의 포물선식으로 전장이 증가함에 따라 포란수는 크게 증대하는 것으로 나타났다(Fig. 7).
재생산에 참여하는 홍가자미의 군성숙전장을 알기 위해 산란기로 추정되는 1-4월의 암컷 107마리 (전장 24.7-42.9 cm)와 수컷 132마리 (전장 22.6-39.6 cm)를 대상으로 중숙단계 이상에서 출현하는 성숙개체의 출현비율을 구하였다(Fig. 8). 암컷은 28 cm에서 4.7%, 31 cm에서 24.3%, 33 cm에서 75.0%이었고 39 cm 이상에서는 100%로 나타났으며 수컷은 22 cm에서 3.8%, 25 cm에서 21.5%, 28 cm에서 64.1%이었고 36 cm에서 95%이상으로 나타났다. 홍가자미의 최소성숙전장은 암수 각각 28.4 cm , 22.6 cm 이었고 50% 성숙전장은 체급별 성숙개체의 출현 비율을 logistic식에 적용한 결과 암컷은 32.9 cm, 수컷은 27.1 cm로 추정되었다.
본 연구의 전체 조사기간 중 채집된 홍가자미의 성비에서 수컷비율이 우세하였는데, 산란기에는 암컷의 비율이 38.7% 로 비산란기에 비해 높은 비율을 보였다. Hirose and Minami (2007)의 보고에 의하면 교토연안 홍가자미 산란장에서 수컷이 암컷보다 성비가 우세하다는 것을 관찰하였는데, 수컷 개체수 비율의 우세가 산란시기 전부터 나타나는 것으로 보아 산란장에서 장기간 수컷들이 머무르는 것은 암컷과의 짝짓기 기회를 많이 갖도록 하기 위한 것으로 보고하였다. Yamasaki et al. (1999)은 암컷보다 수컷이 산란장에서 더 오래 머문다고 하였고 같은 산란장내에서 수컷들은 일찍 성숙하며 암컷보다 생식 활동을 보다 길게 한다고 제안하였다. 본 연구에서 산란기간 암수의 평균전장은 비산란기간의 평균전장에 비해 작은 것으로 나타났는데, 이는 상업어선이 산란기간 동안에는 수심이 얕고 비교적 작은 어체와 산란친어가 함께 서식하는 150-200 m층에서 조업을 하고 비산란기간에는 산란을 마친 큰 개체들이 이동하여 서식하는 300-400 m층에서 주로 조업을 하기 때문인 것으로 생각된다.
홍가자미 암컷의 평균 생식소숙도지수(GI)는 1월과 2월에 각각 7.72와 7.44로 가장 높은 값을 보이다가 3월에 3.04로 급격히 떨어지고, 성숙단계를 육안으로 관찰하여 숙도를 구분하였을 때 방란중인 개체는 1월부터 4월 사이에 출현하고 2월에 전체 개체수의 34%로 가장 높은 비율을 보였다. 또한 난소의 발달단계를 조직학적으로 분석한 결과에도 2월과 3월에 방란기의 완숙난이 관찰되었고, 4월이 되면서 방란 후기의 개체들이 관찰되는 것으로 보아 홍가자미의 주 산란시기는 2월과 3월로 추정된다. 홍가자미 수컷의 생식소 숙도지수(GI)도 1월과 2월에 각각 1.54와 1.97로 가장 높은 값을 보였으며 3월이 되면서 1.25로 급격하게 감소하였고, 생식소를 육안으로 관찰한 결과에서도 방란중인 개체가 1-5월까지 출현하지만 2월과 3월에 방정중인 개체의 비율이 각각 31%와 26%로 가장 높은 것으로 보 아 수컷도 암컷과 마찬가지로 2월과 3월이 산란성기로 판단된다. Yamada et al. (2008)의 일본 후쿠시마현 연안해역 홍가자미의 성숙과 산란에 관한 연구에서 홍가자미 암컷 성숙개체의 출현 빈도는 1월 하순에 높았다가 2월 상순부터 4월 하순까지 낮게 나타났으며, 성숙개체와 방란중인 개체는 1월 하순은 낮았고 2월 상순부터 3월 하순까지는 높게 나타났다가 4월 상순 이후에는 감소하였다. 방란후 개체는 1월 하순에는 나타나지 않고, 2월 상순부터 3월 하순까지 낮게 나타나고 4월 상순 이후 증가하였다. 이 결과로 후쿠시마현에 서식하는 홍가자미의 산란 시기는 2-3월로 추정되었으며 Yokohama et al. (1991)의 푼카만(Funka bay)에서 1-4월, 동해 동쪽에서는 2-4월 등으로 본 연구 결과 추정된 동해안 강원연안의 홍가자미의 산란시기와 크게 다르지 않았다.
월별 난경조성에서 산란시기인 1월부터 4월까지 출현하는 난경 0.9 mm이상의 난모세포는 방란을 통하여 산란을 하게 되며 산란을 하지 못한 난모세포는 5월 이후에 퇴화하거나 흡수되는 것으로 여겨진다. 산란기간 동안 전체 난모세포수에 대한 방란되는 난모세포수의 비율은 3.6-27.0%로서 이 수치는 포란수 (Fecundity)에 대한 산란비율로 홍가자미의 산란수(Fertility)로 간주 될 수가 있다. 산란수에 관한 연구로 멸치의 재생산력 연구 (Choi and Kim, 1988)에서 포란한 난중 방란되는 난의 비율은 4.3-13.1%로 부어류인 멸치에 비해 저서어류인 홍가자미의 방란비율은 높은 수준에 있는 것으로 나타났다.
자원관리 매개변수의 하나인 성숙전장은 본 연구에서 암컷의 경우, 최소성숙전장은 28.4 cm, 50% 성숙전장은 32.9 cm이었고 39.0 cm 이상에서는 100% 성숙을 하는 것으로 나타났다. 이에 비해 Ishida and Kitakata (1982)의 홋카이도 연안의 홍가자미에 관한 보고에서 최소성숙전장은 29-30 cm, 50% 성숙전장은 32-33 cm, 100% 성숙전장은 35 cm이었고, Yokohama et al. (1991)의 홋카이도 푼카만의 보고에서 최소성숙전장은 25.5 cm, 50% 성숙전장은 27-28 cm이었다. 또한, Yamada et al. (2008)의 후쿠시마현 연안해역에 대한 보고에서 최소성숙전장은 27 cm, 50% 성숙전장은 29.9 cm, 100% 성숙전장은 37 cm 이었다. 성숙전장에 관련한 이상의 연구 결과들을 기준으로 볼때, 본 연구의 최소성숙전장 및 50% 성숙전장은 각각 28.4 cm, 32.9 cm로서 Ishida and Kitakata (1982)의 29-30 cm, 32-33cm와 거의 일치하였고 100% 성숙체장은 39 cm 이상으로서 Yamada et al.(2008)의 37 cm와 유사한 것으로 나타났다. 같은 조사해역인 홋카이도에서 실시한 Ishida and Kitakata (1982)와 Yokohama et al. (1991)의 결과는 큰 차이를 보였고, Yamada et al.(2008)의 최소성숙전장과 50% 성숙전장은 Ishida and Kitakata (1982)와 Yokohama et al.(1991) 두 결과의 중간값 수준이고 100% 성숙전장은 가장 높은 값을 보였다. 이러한 성숙과 성장에 대한 차이는 조사해역의 개체군 특성에 따른 자원상황이나 조사시기와 관련한 해양환경의 변화 등과도 관계가 있을 것으로 생각된다.
어류의 성숙·산란과 수온의 중요성에 관한 연구로는 대부분이 인위적인 수온조절에 의한 사육실험(Solomon et al., 2011; Smith and Walker, 2004; Tan-Fermin et al., 1997)으로 수온 증가가 산란기작에 영향을 미치는 것으로 나타났으며, 자연상태의 조사로서는 조간대 수온상승이 망둑어류의 생식소 숙도 지수를 지속적으로 높인다는 결과가 연구된 바가 있다(Shiota et al., 2003). 가자미류는 저서정착성 생활을 하는 어종으로 표층과 중층 생활을 하는 다른 어류들에 비해서 온도에 대한 민감성이 상대적으로 낮은 것으로 알려져 있지만(Fonds, 1979; Murawski, 1993), 문치가자미의 경우 산란장의 수온증가에 의해 산란시기가 점차적으로 길어지고 산란 수층 또한 깊은 수심으로 이동하는 경향을 보인다고 하였다(Seo et al., 2010). 또한 비스카이만에서 가자미류 초기생활사 단계의 생체량과 개체수 는 산란시기의 수온과 관련 있는 것으로 겨울과 봄에 주로 산란 하는 차가운 북부해역의 가자미에 대해서는 해수온난화가 가입상황을 악화시키는 것으로 나타났다(Désaunay et al., 2006; Hermant et al., 2010). 동해안 저층수의 수온은 1960년대 이후 점차적으로 상승하여 계절별로 0.24-0.4℃ 정도 상승한 것으로 보고한 바 있어(Yoon et al., 2010), 우리나라 동해안의 깊은 수심에서 주로 서식하는 홍가자미를 비롯한 냉수성 가자미류의 생식생태와 산란장의 변화가 생길 지가 주목되는 것으로 효율적인 자원관리를 위해 해양수온 변동에 따른 생식특성과 가입 상황을 지속적으로 모니터링 해야 할 것으로 여겨진다.