We conducted eight surveys in the northern East China Sea (ECS) in winter (February – April), summer (July), and autumn (October) 2004-2009, to investigate the seasonal distribution of
살오징어(
살오징어의 수명은 1년 내외로, 산란 후 사망하는 것으로 알려져 있다. 표층에서 200 m 수심에 주로 분포하고 서식가능 수온은 5-27℃이다(Roper et al., 1984). 외양회유성 어종으로 알려진 이들은 동중국해 및 동해 남부에서 부화 후 북서태평양 일본 측 연근해와 동해 북부해역으로 섭이회유를 시작한다. 부화 후 7-8개월이 되어 외투장(mantle length, ML)이 약 20 cm 내외가 되면 성숙에 이르고, 산란을 위해 남하회유를 시작한다(Okutani, 1983). 산란시기의 어미는 수온 15-23℃ 범위의 밀도약층(pycnocline) 부근에 반부유성 난괴(egg mass)를 방출한다. 난괴는 지름 40-80 cm의 구상(spherical)으로 외벽은 젤리질로 이루어져 있고, 약 20만 립의 난을 포함하고 있다(Bower and Sakurai, 1996). 수온 15-23℃에서 정상적인 배발생(embryonic development)이 가능한 것으로 알려져 있으며, 18-19℃ 에서 4-6일 후 부화에 이른다(Sakurai et al., 1996). 부화유생은 표층으로 부상하며, 수온 17-23℃ 범위에서 주로 발생한다(Bower, 1997). 이후 성장하면서 유생의 분포수심은 점점 깊어지고, 20-80 m 수심에서 주로 분포한다(Yamamoto et al., 2007; Kim et al., 2011). 유영능력이 미약한 유생은 주로 해류에 의해 분포가 결정되는 부유유생 시기를 거친다(Sakurai et al., 1996).
연중 산란하는 것으로 알려진 살오징어는 산란이 집중되는 계절과 회유경로에 따라 여름, 가을, 겨울 산란군의 세 개 계군(population)으로 구분된다(Murata, 1989; Kim and Kang, 1995). 산란장은 동중국해의 대륙붕과 대륙사면을 따라 동해 남부 지역까지 분포하고 있으며, 각 계절 산란군의 산란시기에 따라 산란장의 북상(가을) 및 남하(겨울)가 발생한다(Shojima, 1972; Sakurai et al., 2000). 겨울 산란군은 타이완 북부 해역에서부터 큐슈 서부해역에 걸쳐 주로 산란하며(1-4월), 대부분의 유생은 쿠로시오(Kuroshio) 해류를 따라 일본 남부의 태평양 측으로 수송된다. 가을 산란군은 북부 동중국해부터 동해 남부해역에서 산란하며(10-12월), 유생은 대마난류(Tsushima Warm Current)를 따라 대한해협을 통과하여 동해에 주로 분포한다(Goto, 2002; Yamamoto et al., 2002). 한편 여름 산란군은 동중국해 중부해역부터 동해 남부해역에서 산란하며(6-8월), 다른 두 계절산란군에 비해 상대적으로 적은 자원량을 가진 것으로 알려져 있다(Nasu et al., 1991).
살오징어의 난을 해양에서 직접 채집한 기록은 아직 전무하나, 일본 연구자들이 ROV (Remotely Operated Vehicle)를 이용하여, 대한해협에서 난괴를 관찰한 보고가 있었다(Sakurai et al., 2002). 일본에서는 1973년 이후 가을 산란군을 대상으로 동해 남부해역에서 유생조사를 매년 시행해 오고 있고(JSRFRL, 1973), 또한 큐슈 남서부해역에서 겨울 산란군을 대상으로 한 유생조사를 1998년 이후 매년 시행하여, 해양환경 특성과 함께 유생분포특성을 규명하는 연구가 수행된 바 있다(Bower et al., 1999; Goto, 2002; Yamamoto et al., 2002, 2007). 이러한 장기적인 모니터링 결과를 활용하여, Goto et al. (2002)은 성어의 자원량이 급격히 증가한 1980년대 후반 이후의 유생분포범위가 이전과 비교하여 지리적으로 넓게 확장하였음을 보고하였고, Sakurai et al. (2000)은 유생밀도지수와 어획량간의 높은 상관관계를 통해, 유생밀도지수를 활용한 어획량 예측 가능성을 제시하였다. 우리나라에서 살오징어 유생에 관한 연구는 일본에 비하면 상당히 미미한 수준으로, Kim et al. (2011)에 의해 북부 동중국해 해역과 남해 근해에서의 살오징어 유생의 수직· 수평분포가 연구된 바 있다. 그러나 각 계절별로 1회씩 시행된 제한적인 유생조사와 비교적 적은 채집 개체수 등은 연구해역의 살오징어 유생분포 특성을 이해하는데 상당한 제한요인으로 작용하였다.
수산자원을 효율적으로 관리하기 위해서는 대상생물의 초기 생활사 단계에 대한 연구를 통해 가입매커니즘을 이해할 수 있어야 한다. 과거 동해에서 살오징어 자원과 대등한 생태적 지위를 차지하던 명태자원은 1990년대 들어 자취를 감추었다. 이는 무분별한 남획과 해양환경변화에 따른 가입량 감소 때문인 것으로 알려져 있다(Lee and Kim, 2010; Kang et al., 2013). 현재 가장 중요한 수산자원 중 하나인 살오징어 자원을 지속적으로 이용하기 위해서는 해양환경변화와 과도한 어획에 대처 할 수 있도록 효율적인 자원 관리 방안이 필요하며, 이를 위한 대상 자원의 산란장 규명과 유생분포특성에 관한 연구가 선행되어야 한다. 하지만 현재까지 우리나라 주변해역에서 살오징어를 대상으로 한 초기생활사 단계 및 가입기작에 관한 연구는 매우 부족한 실정이다.
본 연구는 살오징어 유생분포 특성에 관한 Kim et al. (2011)의 연구를 확장하여, 북부 동중국해에서의 계절적인 환경변동성과 살오징어 유생의 계절분포 특성을 밝히기 위해 계절 산란군별 유생조사를 2004-2009년에 걸쳐 시행하였다. 또한 관측결과를 바탕으로, 연구해역에서의 계절별 전선형성과 유생분포의 관계 및 계절 산란군별 산란장 분포특성을 파악하였다.
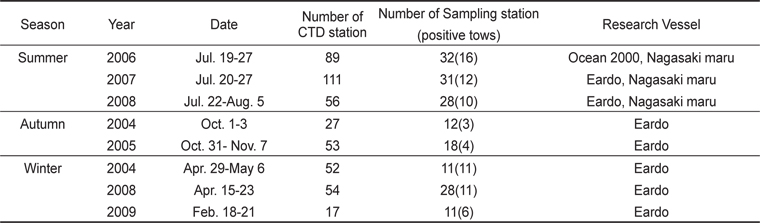
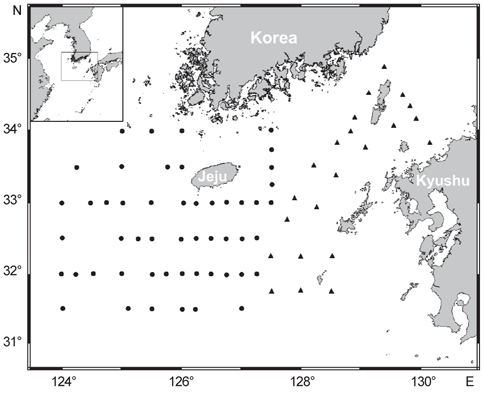
북부 동중국해에서의 살오징어 유생 계절분포 특성을 밝히기 위하여 2004-2009년 동안 총 8회의 살오징어 유생조사를 여름(3회), 가을(2회), 겨울(3회)철에 각각 실시하였다(Table 1, Fig. 1). 특히 2006-2008년에 걸쳐 연속적으로 수행된 여름철 조사는 북부 동중국해와 함께 큐슈 서부와 대한해협 등의 일본 EEZ내에서 동시에 조사가 수행되어, 더 넓은 해역에서의 유생분포 특성을 알아볼 수 있었다. 유생채집에 사용된 조사기구는 직경 60 cm의 봉고넷(bongo nets)으로, 어류플랑크톤 채집에 주로 쓰이는 0.333 mm의 망목을 사용하였다. 살오징어 유생은 표층-100 m 수심에서 분포하므로, 100 m 수심 상부의 전 수층에서 분포하고 있는 살오징어 유생을 채집하기 위해 경사채집(oblique tow)을 시행하였다. 봉고넷 예인 시, 배의 속도는 약 2 kt이며, 주로 100 m 수심에서 표층까지 예인 하였으나, 수심이 얕은 정점에서는 바닥 부근까지 내린 후 예인 하였다. 네트가 최대수심에 도달하면, 수층 내에서 네트가 안정화 될 수 있도록 침강시간의 1/10을 조정시간으로 가졌다. 예인 당시의 네트위치와 수온 등의 환경자료 수집을 위해 네트에 data recorder (TDR-2050, RBR Ltd., Canada)를 부착하였고, 각 네트의 여수량 측정과 채집 유생의 정량화를 위하여 입구 중앙에 유량계(flowmeter)를 부착하였다. 또한 연구해역의 전반적인 환경 특성을 관측하기 위해 CTD (SBE-911, Sea-Bird Elec. Co., U.S.A.)를 이용하여 수온, 염분 등의 환경자료를 수집하였다.
[Table 1.] Summary of survey for collecting Todarodes pacifiucs paralarvae during 2004-2009
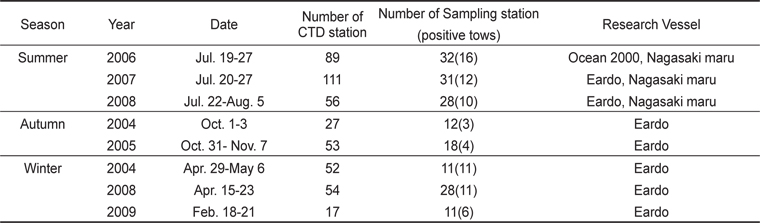
Summary of survey for collecting Todarodes pacifiucs paralarvae during 2004-2009
채집된 어류·동물플랑크톤 시료는 즉시 약 5°C 냉장고에서 1-2시간 보관되었으며, 이후 선상에서 동물플랑크톤 시료로부터 살오징어 유생이 선별되었다. 유생은 5% 중성 포르말린 용액에 고정되었고, 실험실에서 해부현미경으로(10-100배) 관찰하여, 외투장(mantle length)의 길이를 0.1 mm 단위까지 측정하였다. 유생조사에서 채집된 살오징어 유생의 동정에는 Okutani (1965)와 Wormuth et al. (1992)이 제시한 살오징어 유생외투(mantle)의 형태적 특징, Ommastrephidae과(Family) 유생단계에서 특징적인 융합된 촉수(tentacles)의 형태, 외투 표면의 색소포(chromatophore) 분포 패턴과 촉수말단의 흡반(suckers) 형태 및 배열 등이 이용되었다.
연구해역인 북부 동중국해의 전반적인 계절별 해양환경특성을 알아보기 위하여, T-S 다이어그램과 표층 및 50 m 수층의 수온, 염분 수평분포도를 이용하였다. 살오징어 유생의 수직분포 범위는 표층-100 m 수심으로 알려져 있으며, 부화직후 일시적으로 표층으로 부상한 후 50 m 수심을 중심으로 가장 높은 밀도를 보인다(Watanabe, 1965; Yamamoto et al., 2007; Kim et al., 2011). 본 연구에서는 살오징어 분포특성에 관한 이전연구(Goto, 2002; Yamamoto et al., 2002)와 동일하게 환경특성과 관련한 유생분포 특성을 밝히기 위해 50 m 수층의 수온, 염분값을 이용하였다. 유생분포 지역에서의 유생의 밀도는 유량계의 회전수에 보정실험을 통해 얻어진 보정계수 0.027을 곱하여 해수면 10 m2 아래에 존재하는 개체수로 환산하였다(개체수/10 m2).
한편 살오징어 유생의 분포와 조사기간 중 연구해역에서 형성되는 열전선(thermal front) 위치의 관계를 밝히기 위해 각 계절과 조사시기 별 열전선의 위치를 파악하였다. 열전선은 주로 수온의 수평적인 미세구조로서 파악되며 수온변화가 수평적으로 극심한 지역을 전선으로 정의할 수 있다. 본 연구에서는 전선이 형성되는 지역의 수온범위에서 중앙값(median)을 취하여 각 조사시기 별 전선의 중심위치로 정의하였으며(per. com. Han, KODC/NFRDI), 각 계절별 전선의 위치와 계절별로 통합된 유생분포를 이용하여 계절별 전선형성과 유생분포 특성을 살펴보았다.
각기 다른 계절에 채집된 유생의 평균 외투장간 유의차 검정과 유생 성장에 따른 환경변동특성을 규명하기 위하여 SPSS (version 12.0) 통계프로그램을 이용하였다. 계절간 유생체장 분석에 이용된 자료는 정규성(normality)과 등분산성(homogeneity of variance) 검정을 통해, 이들 가정 성립 시 one-way ANOVA test를 수행하였고, 사후검정을 위해 Scheffe’s test를 실시하였다. 또한 성장에 따른 환경변동성 파악에는 부화유생 (1 mm≤ ML <2 mm)과 2 mm ML 보다 큰 유생의 분포해역에서 수집된 수온·염분값(50 m 수심)을 이용하였고, 두 그룹간 서식환경의 유의차 검정을 위해 T-test를 실시하였다.
동중국해 북부해역에서 살오징어 유생조사를 통해 채집된 가장 작은 유생의 크기 집단은 1-2 mm ML였다. 유생의 분포를 바탕으로 조사해역과 주변해역에서의 계절별 살오징어 산란장 형성 가능성과 산란장 분포특성을 알아보기 위해 2 mm ML 보다 작은 개체들의 계절별 밀도분포를 이용하였다.
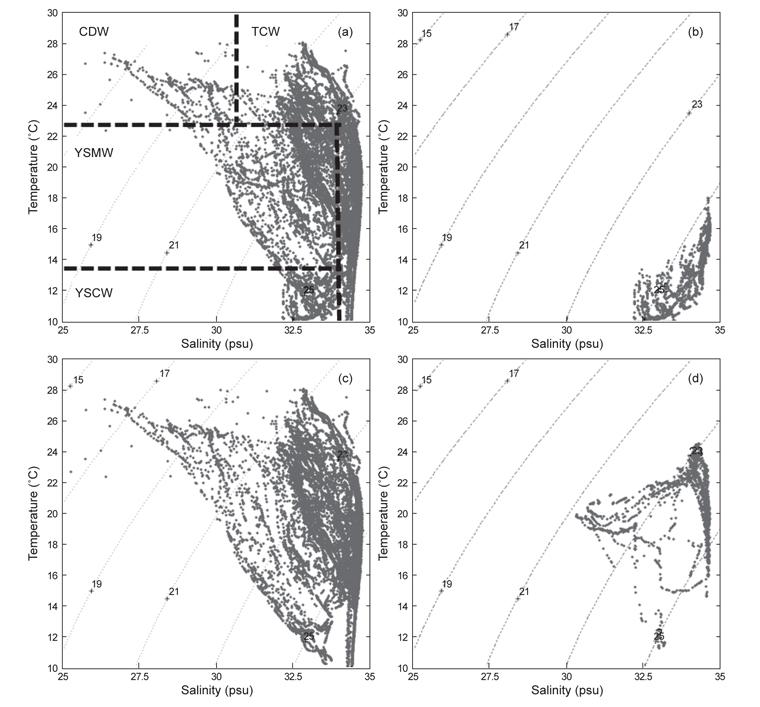
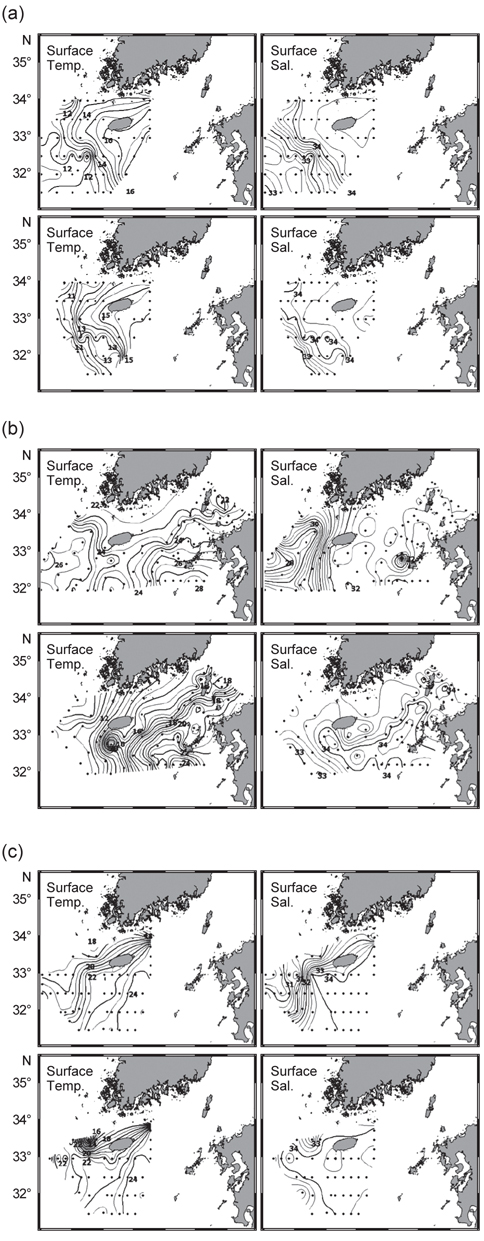
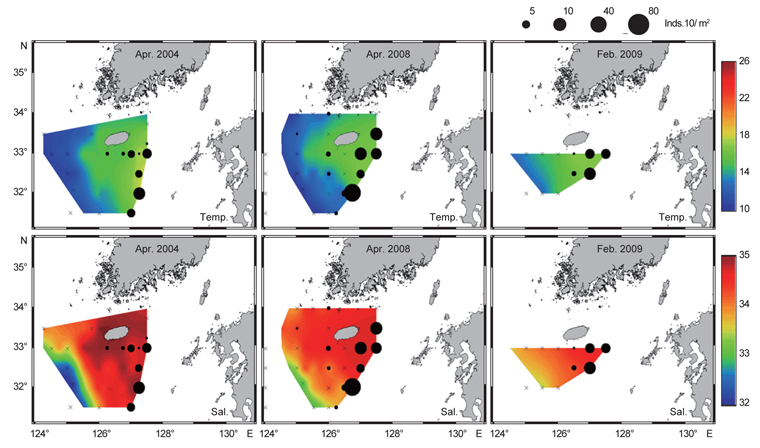
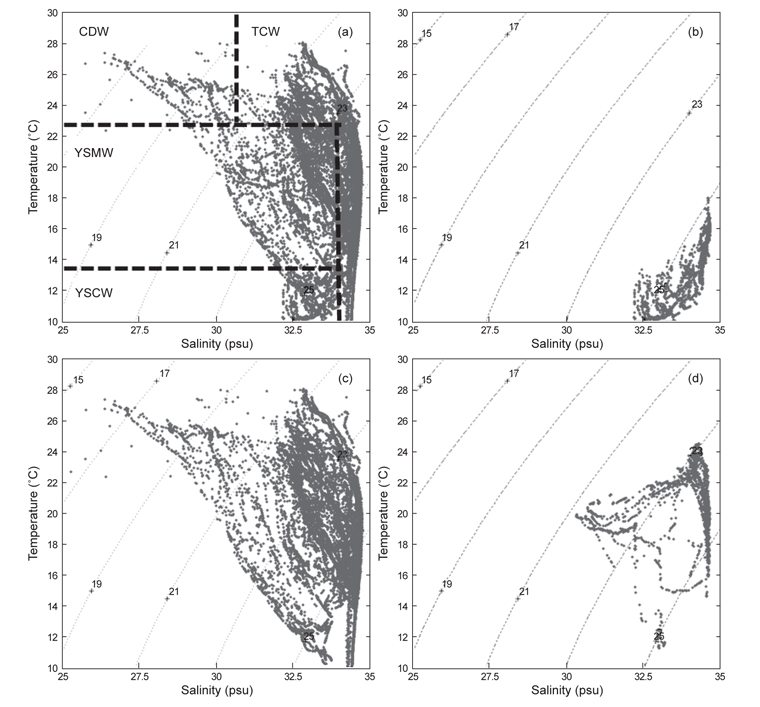
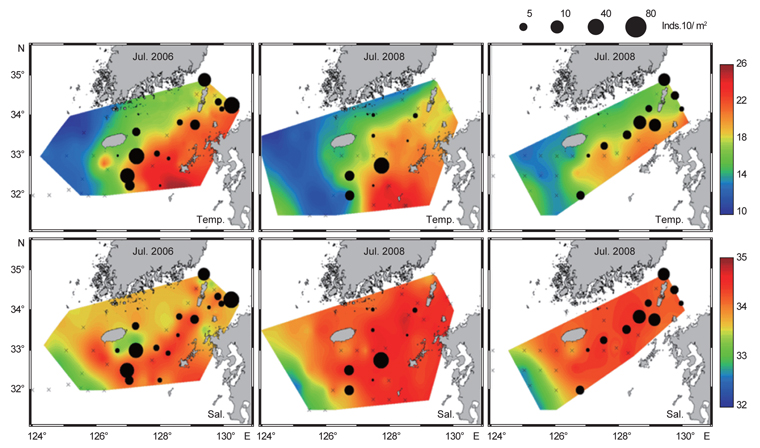
북부 동중국해의 계절적인 해양환경특성을 알아보기 위하여, 계절별로 2008년 4월(겨울), 2006년 7월(여름)과 2005년 10월(가을)의 해양관측을 통해 얻어진 T-S 다이어그램(Fig. 2)과 표층 및 50 m 수층의 수온·염분 분포도(Fig. 3)를 나타내었다. 먼저 T-S 다이어그램을 통해 연구해역의 환경특성을 살펴보면, 북부 동중국해의 수괴는 대마난류수(고온·고염수), 황해저층냉수(저온수), 양자강 희석수(저염수)와 황해혼합수로 특징지어졌고(Fig. 2a, KORDI, 2010; Gong et al., 1996), 계절적으로 현저히 다른 환경특성을 보였다(Fig. 2b-d). 2008년 4월의 수온·염분의 범위는 10-18℃, 32.2-34.6 psu로 나타났고, 고온의 대마난류수와 상대적으로 낮은 수온의 중국 연안기원의 저염수가 관찰되었다(Fig. 2b). 2006년 7월은 상층에서 대마난류수와 양자강 저염수가 관측되었고, 저층에서는 황해저층냉수가 분포했다(Fig. 2c). 수온은 10.2-27.1℃, 염분은 26.6-34.7 psu 범위로 나타났다. 2005년 10월은 수온 10.2-27.1℃, 염분 26.6-34.7 psu의 범위로 나타났고, 여름철 성층화와 겨울철 연직혼합의 전이(transition) 형태를 보였다(KORDI, 2010) (Fig. 2d).
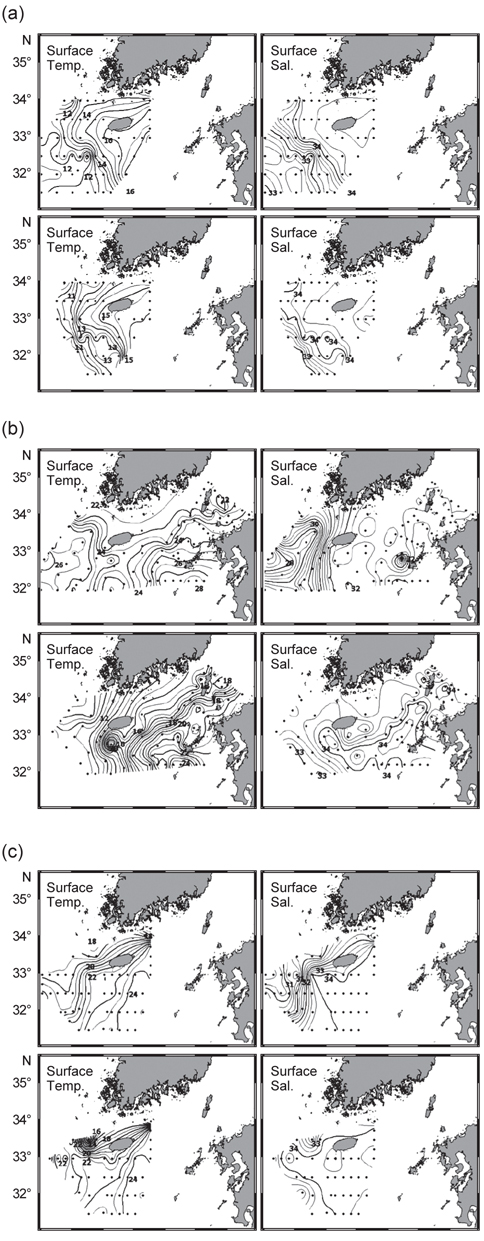
2008년 4월은 대마난류와 중국 연안수 사이의 열염전선(thermohaline front)이 연구해역의 서부에서 북서-남동 방향으로 형성되었다(Fig. 3a). 표층과 50 m 수층의 수온 차를 살펴보면, 연직 혼합이 잘 이루어져 있음을 알 수 있었다. 2006년 7월의 경우 표층에서, 30 psu 이하의 양자강 희석수가 제주도 서쪽 근해까지 확장하는 형태가 관찰되었고(Fig. 3b) 또한 50 m 층에서는 제주도 북서부 해역에서 황해저층냉수가 남쪽으로 확장해있었다. 대마난류수와 연안수 사이에 형성되는 열전선(thermal front)은 제주 남부해역에서 남북방향으로 형성되어, 대마도 부근까지 길게 연결되는 형태로 나타났다. 또한 계절적 특성으로 연구해역 전반에 걸쳐 성층화가 이루어져 표층과 50 m 수층에서의 큰 수온 차가 관찰되었다. 2005년 10월은 연구해역이 고온·고염의 동편과 저온·저염의 서편으로 구분되는 특징을 나타냈다(Fig. 3c). 제주도 서부해역, 특히 표층에서 강한 열염전선이 관찰되었다. 표층과 50 m 수층에서 대마난류의 영향을 받은 34 psu 등염분선이 제주도 서부해역까지 넓게 확장하는 형태를 보였다. 또한 제주 서부 근해에서 국지적인 저온수의 발생(50 m 수심)이 관찰되었다.
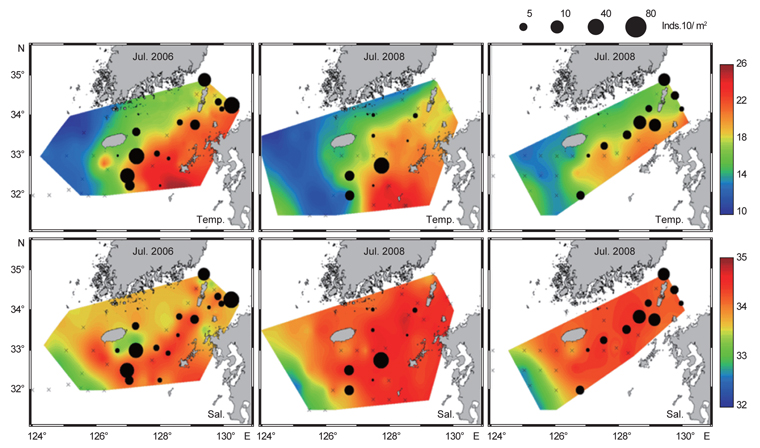
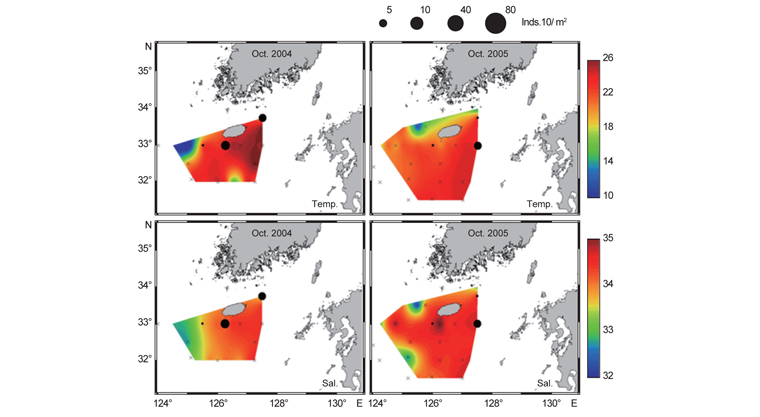
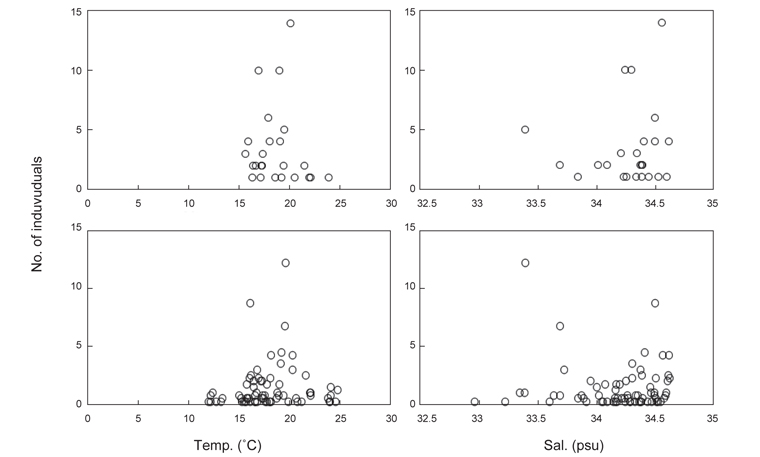
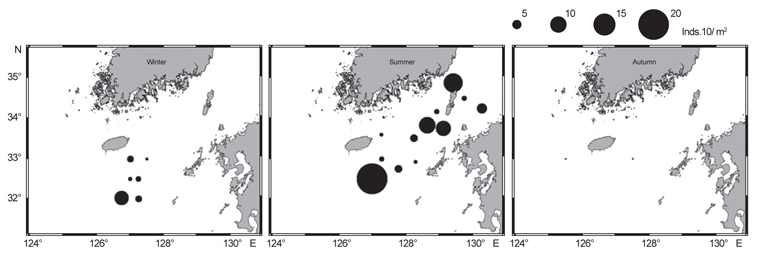
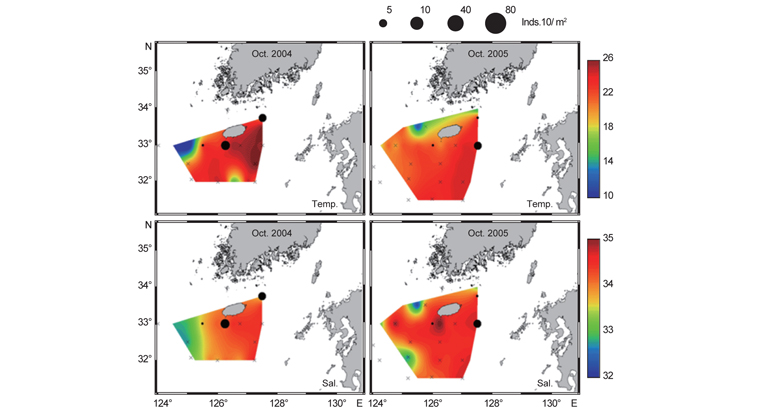
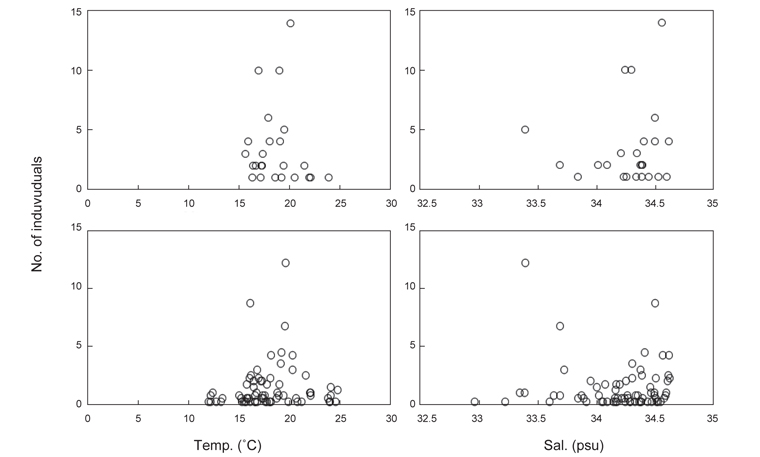
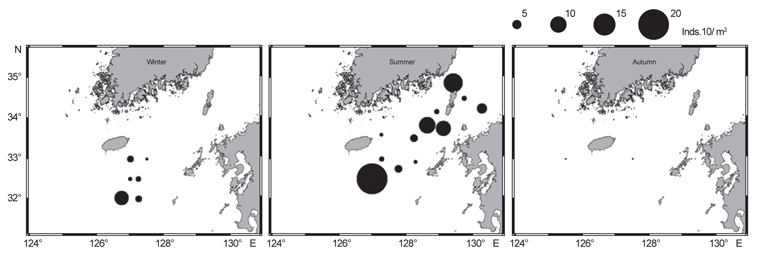
북부 동중국해 해역에서 살오징어 유생의 계절적 수평분포 특성을 밝히기 위하여, 2004-2009년의 연구기간 동안 총 8회의 유생조사를 실시하였고, 총 482개체의 유생표본이 채집되었다. Fig. 4는 각 조사시기별 50 m 수심의 수온·염분과 유생 밀도 분포를 나타낸다. 겨울철(2-4월) 유생출현 정점에서의 유생밀도는 1-105개체/10 m2 범위로 나타났다. 유생은 수온 13.3-17℃, 염분 34-34.6 psu 범위의 제주 남동부 근해에서 높은 밀도로 발생하였다. 비록 제주 남동 해역 보다는 상대적으로 낮은 밀도지만, 유생의 일부는 제주도 남서부와 서부해역에서도 발생했다. 2008년 4월, 제주 남동 해역에서의 유생밀도는 2004년 4월과 비교하여 상대적으로 높게 나타났다. 2009년 2월의 경우 악천후로 인해 상대적으로 제한된 지역적 범위에서 조사가 이루어졌으나, 유생의 분포 패턴과 해양환경은 4월 조사와 비슷한 형태를 나타내었다. 여름철(7월) 유생출현 정점에서의 유생밀도는 1-83개체/10 m2 범위로 나타났다(Fig. 5). 유생은 주로 수온 17-21℃, 염분 33.5-34.5 psu의 제주 남동해역과 큐슈 북서부의 대한해협에서 발생했다. 유생밀도는 2006년에 가장 높았으며, 2008년, 2007년 순으로 나타났다. 특히, 2007년은 다른 두 해에 비해 대한해협에서의 유생분포 밀도가 상당히 낮게 관찰되어 이들 분포의 연변동성이 크게 나타났다. 가을철(10월) 조사의 경우 유생출현 정점에서 유생밀도는 1-13개체/10 m2의 범위였다(Fig. 6). 유생은 수온 22.7-24.5℃, 염분 33.6-34.3 psu 범위의 제주도 남서부 근해와 북동쪽 정점에서 발생하였다. 2004년 조사에서는 제주 남서부 해역과 북동쪽에서 유생이 발생하였으나 2005년 조사에서는 남동부해역에서 유생이 발생하는 연변동성이 관측되었다. 특히, 가을철 조사에서는 다른 두 계절에 비해 가장 낮은 유생 밀도를 나타냈다. 또한 유생발생의 남방한계가 북위 33°N로서 여름철(32°N)과 겨울철(31.5°N)에 비해 가장 높게 나타나는 특징을 보였다.
전반적으로, 연구해역에서 겨울철과 여름철 유생의 밀도가 높았으며, 그 분포범위 또한 넓게 나타났다. 제주 남동부해역은 전 조사시기 동안 유생이 발생하였고, 유생밀도 역시 다른 지역과 비교하여 높게 나타났다. 유생분포 해역에서의 환경특성으로서 수온·염분 범위는 전 계절을 통틀어 각각 13.3-24.5℃, 33.5-34.6 psu였고, 채집된 유생의 90% 이상이 수온 15-23℃에서 발생하였다. 한편 각 연도별로 동일한 시기에 수행된 유생조사들 간에는 유생의 지역적 분포 패턴 및 밀도 상의 연변동성(특히, 여름철 7월말 조사)이 관찰되었다.
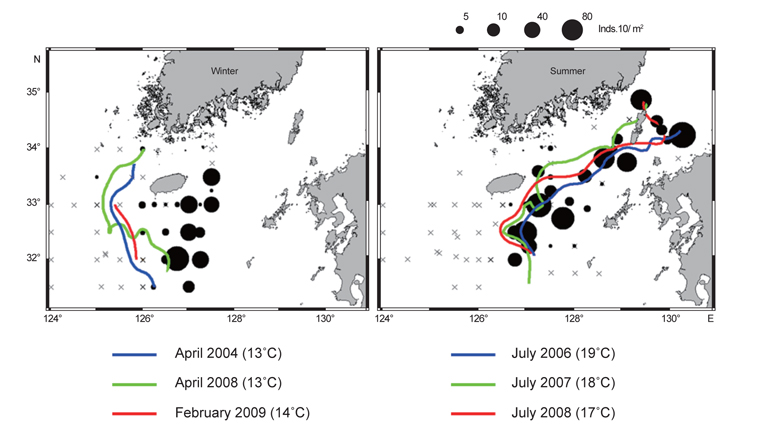
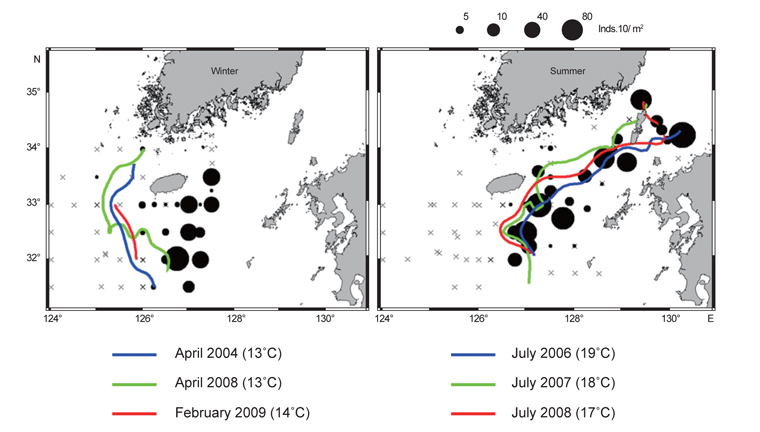
Fig. 7은 여름철과 겨울철 조사 시기에 형성되는 열전선(thermal front)의 위치와 유생분포를 함께 나타내었다. 겨울조사 시기의 전선은 동경 126°이서해역인 제주 서부 근해에서 북동방향으로 형성되었다. 4월에는 13℃ 등온선을 중심으로 전선이 형성되며 2월조사에서는 14℃ 등온선을 중심으로 전선이 형성되었다. 유생분포는 전선주변해역에서 상당히 낮게 나타났고, 제주 남동부 해역에서 높은 밀도를 보였다. 전선의 동쪽인 제주 서부해역에서는 상대적으로 낮은 밀도지만 지속적으로 유생이 출현하였다. 여름철 전선은 동경 126.5°동쪽, 제주 남동 근해에서 남북방향으로 형성되었다. 2006년에는 19℃ 등온선을 중심으로 전선이 형성되며, 2007년에는 18℃, 2008년에는 17℃ 등온선을 중심으로 전선이 형성되었다. 유생분포는 제주 남동해역의 전선역 주변에서 상당히 높게 나타났고 대한해협까지 전선역을 따라 높은 밀도분포를 보였다. 가을조사에서 전선은 동경 125°동쪽해역에서 동서방향으로 형성되며, 전선역 주변에서 유생 밀도가 높게 나타났다. 그러나 가을 조사의 경우 전선이 조사해역의 북단에서 형성되었으나, 전선역 주변에서 충분한 유생조사가 이루어지지 못해 전선형성과 유생분포특성을 이해하기에는 관측자료의 한계가 있었다.
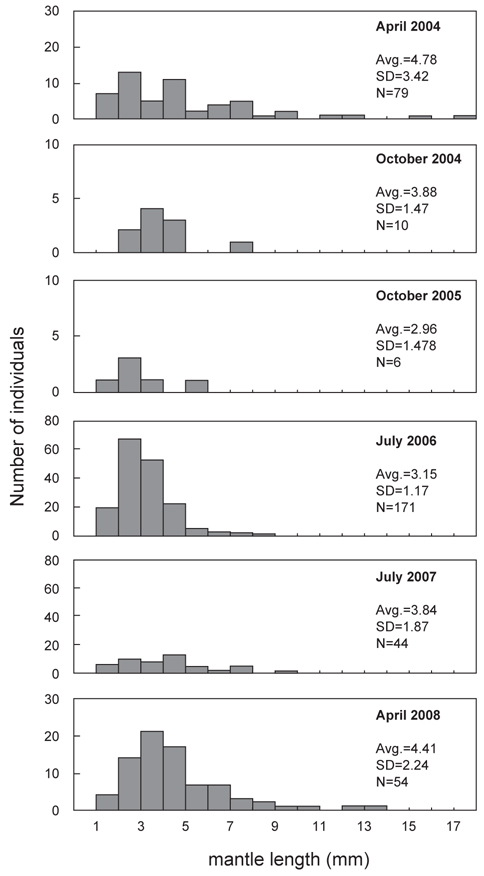
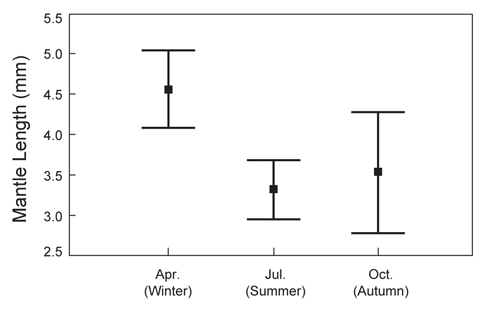
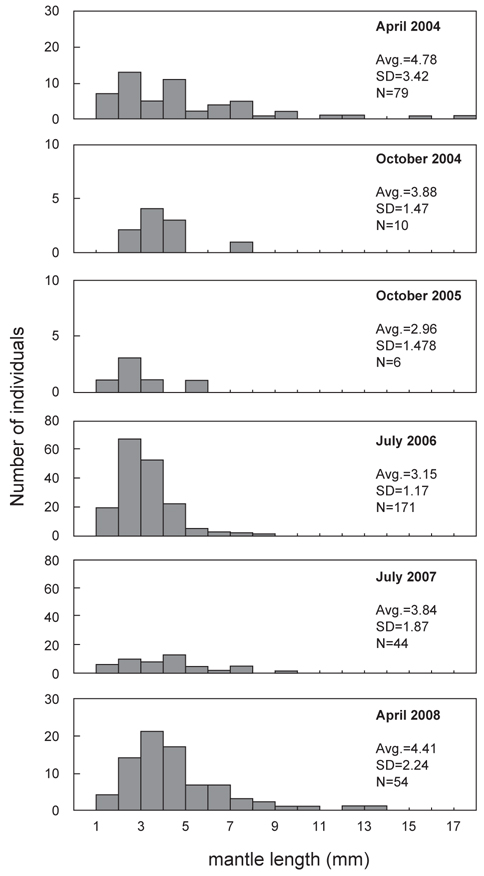
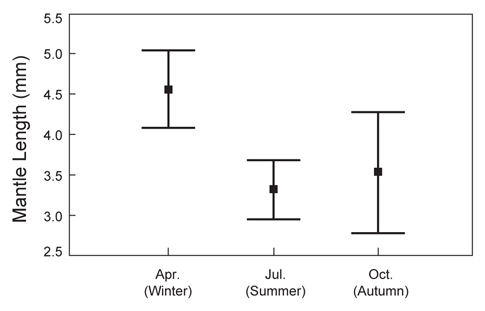
총 8회의 유생조사 중 각 계절을 대표할 수 있도록 동일한 월(4월, 7월, 10월)에 실시된 유생조사에서 얻어진 유생의 외투장 빈도 분포를 나타내었다(Fig. 8). 동일한 해역에서 발생한 유생의 외투장을 비교하기 위하여, 여름철 동경 134°E 이상의 지역에서 채집된 유생은 분석에서 제외하였다. 4월 채집된 유생은 외투장 1.2-17.0 mm 범위였으며, 2004년에 4.78 mm (SD=3.42 mm)와 2008년에 4.41 mm (SD=2.24 mm)의 평균 외투장을 나타내었다. 7월의 유생 외투장의 범위는 1.0-9.5 mm로 나타났고, 평균 외투장은 2006년에 3.15 mm (SD=1.17 mm)와 2007년에 3.48 mm (SD=1.87 mm)로 나타났다. 10월에 채집된 유생의 외투장은 1.8-7.3 mm의 분포를 보였으며 평균 외투장은 2004년에 3.88 mm (SD=1.47 mm) 이고 2005년에 2.96 mm (SD=1.50 mm)로 나타났다.
전반적으로, 본 연구에서 채집된 유생의 외투장은 총 8회의 조사를 통틀어 1.0-17.0 mm의 범위로 나타났으며, 부화직후의 1.0 mm 이하의 유생은 채집되지 않았고, 10 mm 이상의 상대적으로 큰 유생은 4월에 수행된 조사에서만 나타났다. 각 계절별로 채집된 유생의 외투장 간에는 통계적으로 유의한 차이를 나타내었으며(single-factor ANOVA, F=7.074,
유생의 성장에 따른 유생분포 환경의 변동특성을 알아보기 위하여, 2 mm 이하의 유생(부화유생)과 2 mm 보다 큰 개체들이 출현한 수온/염분 환경(50 m 수심)을 나타내었다(Fig. 10). 부화유생이 출현한 해역은 수온 15.5-23.8℃, 염분 33.4-34.6 psu 범위였으며, 2 mm 보다 큰 유생이 출현한 해역의 환경은 수온 11.8-24.6℃, 염분 33.0-34.6 psu 였다. 부화유생과 2 mm 이상의 유생에 출현한 환경은 통계적으로 유의한 차이가 없는 것으로 나타났다(T-test,
본 연구를 통해 채집된 각 계절별 부화유생(1 mm ≤ML< 2 mm)의 분포를 Fig. 11에 나타내었다. 겨울철 부화유생은 주로 제주 남동부해역에 분포하였다. 유생의 밀도는 1-10개체/10 m2 범위로 나타났다. 여름철 부화유생은 제주 남동부해역에서부터 대한해협부근까지 발생범위가 광범위하였으며 다른 두 계절에 비해 현저히 높은 밀도를 보였다. 유생밀도는 2-20개체/10 m2 였다. 가을철의 경우 연구해역에서의 유생밀도는 다른 두 계절과 비교하여 상당히 낮게 나타났고(1개체/m2), 유생분포의 남방한계는 가장 고위도에 위치하였다.
Bower et al. (1999)은 큐슈 남부에서의 살오징어 유생조사를 통하여 유생이 쿠로시오와 연안수(inshore waters) 사이에서 형성되는 전선역에서 가장 높은 풍도를 나타낸다고 보고하였다. 또한 Yamamoto et al. (2002)은 동해 남서부 해역의 오키섬(Oki Island) 부근의 유생조사를 통해 오키섬 남서부의 난수(warm water)와 북동쪽의 냉수(cold water) 사이에 형성되는 수온 21-22℃의 전선역에서 대부분의 유생이 분포한다고 보고한 바 있다. 전선역에서 두족류 유생의 밀도가 높은 분포특성은 살오징어와 비슷한 생활사를 가지는 종으로 알려져 있는 북동 대서양의
동해 남서부 해역의 오키섬 주변에서 가을철 채집된 살오징어 유생은 수온 16.2-22.3℃범위에서 발생하였다(Yamamoto et al., 2002, 2007). 본 연구에서 채집된 유생은 비록 여러 계절에 걸쳐 채집되었지만, 난의 정상적인 배발생과 유생생존에 적합한 수온으로 알려져 있는 15-23℃ 범위(Sakurai et al., 1996)에서 대부분 채집되었다. Yamamoto et al. (2007)은 부화직후의 유생이 수온 18.9-22.3℃에서 주로 분포하며, 이후 유생이 성장함에 따라 서식가능 수온 범위가 넓어지고, 특히 저온에 대한 생존 가능 범위가 점차 넓어진다고 하였다. 본 연구 결과에서 유생의 성장에 따른 유생발생 해역의 수온·염분 범위간에는 유의한 차이가 없는 것으로 나타났으나, 수온·염분의 하한값은 유생의 성장에 따라 감소하였다. Furukawa and Sakurai (2008)는 살오징어 부화유생의 생존을 위해 염분 29.5 psu 이상이 요구된다고 보고한 바 있다. 본 연구해역에서의 30 psu 이하의 저염수는 여름철 표층에서 제주도 서부근해까지 출현하였다. 이는 여름철 양자강을 통해 북부동중국해로 유입되는 양자강 유출수의 영향인 것으로 알려져 있으며, 연구해역에서 여름철에 관찰되는 특징적인 환경특성이다(KORDI,2010). 밀도약층 부근에서 부화하여 일시적으로 표층으로 부상하는 살오징어 부화유생의 경우 이러한 저염수의 영향을 받을 가능성이 있다. 그러나 여름철 제주 남부해역에서 형성되는 전선은 이들 유생의 분포를 제주 남동부 해역으로 제한하며, 30 psu 이하의 저염분수가 출현하지 않은 제주 남동부해역 등에 주로 분포하는 유생의 생존에는 큰 영향이 없는 것으로 판단된다. 따라서, 연구해역에서 염분보다는 수온이 이들의 생존에 중요한 역할을 하는 환경변수로 생각되며, 특히, 겨울철 저온수가 주로 존재하는 제주 서부해역에서의 낮은 유생의 밀도는 저수온에 대한 유생의 취약성을 나타내는 것으로 사료된다.
부화 시의 유생 외투장이 0.95 mm (Sakurai et al., 1996) 정도인 것을 고려하여, Goto et al. (2002)은 동해 남동부 해역에서 10-11월에 채집된 1.5 mm보다 작은 유생의 분포로부터 잠재적 산란장을 추정한 바 있다. 그 결과로서, 이들의 산란장은 대한해협과 동해 남서부의 혼슈 연·근해지역으로 보고되었다. Sakurai et al. (2000)과 Nasu et al. (1991)등은 산란장 분포의 계절적 변화를 제안하였다. 즉, 겨울철에는 동중국해 남·중부해역의 대륭붕과 대륙사면에서 주로 산란이 발생하며, 여름철에는 동중국해 중부해역에서부터 대한해협까지 산란장이 형성된다. 또한 가을철에는 대한해협에서부터 동해 남서부 해역에서 산란이 발생한다고 하였다. 본 연구에서 겨울철 부화유생은 주로 제주 남부해역에 분포하였다. Kim (2012)은 동중국해에서의 살오징어 알과 유생의 수송과정에 관한 연구를 통해 동중국해 남부의 대륙붕단에서 산란된 개체가 북부 동중국해에 도달하기 위해서는 60일 정도가 소요됨을 밝혔다. 또한 겨울철 동중국해 중부해역(29-30°N, 126.5-127.5°E)에서 산란된 개체는 본 논문의 연구해역으로 도달하는데 15-30일 정도 소요됨을 보고하였다. 부화까지 걸리는 난발생기간(4-5일)과 부화 후 1.5 mm ML에 도달하는데 걸리는 시간(약 15일)을 고려하면 이들의 산란장은 연구해역 이남의 동중국해 중부해역으로 판단된다. 여름철 부화유생은 대부분 제주 남부해역과 대한해협부근에서 발생하였다. 이는 연구해역의 남부에서 산란이 이루어짐과 동시에 대한해협부근까지 산란장이 확장함을 나타내고 있다. 가을철의 경우 연구해역에서 유생밀도는 다른 두 계절과 비교하여 상당히 낮게 나타났고, 유생분포의 남방한계는 가장 고위도에 위치하였다. 또한 살오징어 산란장으로서 적합한 지역은 수온 15-23℃ (50 m 수심)의 해역으로 알려져 있으나 가을철 연구해역의 수온, 특히 산란장 형성 가능성이 큰 제주 남동부해역의 수온은 23℃ 이상으로서 계절에 따른 산란장 북상가능성이 크다(Fig. 3c). 따라서 연구해역의 북부해역(대한해협 및 동해 남부해역)에서 주로 산란이 이루어질 가능성이 큰 것으로 판단된다. 본 연구에서 채집된 부화시기에 가까운 2 mm ML보다 작은 유생들의 계절별 분포패턴은 살오징어의 계절별 산란장 변동특성을 설명해줄 수 있는 중요한 정보로 사료된다.
2004-2009년 동안의 유생조사에서 채집된 살오징어 유생의 외투장은 1-17 mm 범위이며, 평균치는 최저 2.96 mm (2005년 10월)에서 최대 4.78 mm (2004년 4월) 범위였다. Moriken(per. Com., FRA)은 이석을 이용한 연령사정법을 통해 유생의 초기성장에 관한 연구를 수행하여 부화시기의 유생의 외투장은 1.0 mm보다 작으며, 부화 후 7일이 경과하면 1.25 mm로 성장함을 제시하였다. 또한 그는 이후 연속적으로, 외투장 1.8 mm는 부화 후 20일, 4.0 mm는 40일, 10 mm의 유생은 60일이 경과한 것으로 보고하였다. 이들 외투장과 일령의 관계를 고려해 보면, 본 연구에서 채집된 유생의 상당수는 부화 후 30-40일이 경과한 유생으로 추정된다. 이는 곧 부화직후의 유생이 30-40일 동안의 해류에 의한 수송과정을 거쳤음을 뜻한다. 살오징어와 같은 수산생물의 초기 수송과정에 관한 연구는 이들 생물의 분포를 이해하고, 가입 매커니즘을 이해하는데 있어 필수적이다. 즉, 유생이 유영 능력이 미약하여 그들의 분포가 해류에 의해 결정되는 초기 수송과정을 거치면서, 자신들의 생존에 적합한 해역(환경특성 및 먹이환경)으로 수송되느냐 아니면 그렇지 않은 해역으로 수송되느냐에 따라 향후 대상자원으로 가입이 이루어지는지 여부가 결정될 수 있다. 따라서 향후 연구에서, 살오징어 유생의 분포와 함께 해류에 의한 수송과정이 함께 고려된다면, 초기 생활사 단계에서의 유생의 이동경로파악과 함께 동, 서, 남해에서 어획되는 성어자원의 해역별 가입기작을 이해하는데 중요한 역할을 할 것으로 기대된다.
본 연구는 동중국해 북부해역에서 계절별 유생분포특성을 밝히고 계절 산란군별 산란장 형성 가능성에 대해 토의하였다. 그러나 계절별로 수행된 각각의 유생조사 결과를 살펴보면, 유생분포의 계절 변동성과 함께 이들 분포와 풍도 상에서 관측되는 연변동성 또한 상당히 큰 것으로 판단된다. 특히 여름철 제주도 동부해역에서 대한해협까지의 광범위한 지역에서의 유생분포는 2006년(고밀도)과 2007년(저밀도) 사이에 상당한 연변동성을 나타내었다. 이러한 분포밀도의 연변동성은 연구해역에 영향을 미치는 다양한 환경요인의 복합적인 효과로 생각해 볼 수 있다. 먼저 조사해역에서 2006년 표층에서 강하게 나타났던 양자강 저염수의 영향은 2007년에는 조사해역에서 미미하게 관찰되었다. Chang and Isobe (2003)은 북부 동중국해에 영향을 미치는 양자강 유출수 세력의 강약이 바람의 방향과 세기에 따라 결정되며 특히, 여름철 남풍계열의 바람은 저염수를 대한해협 방향으로 이동시키는 주요 요인이라 하였다. 연구해역에서 6월과 7월의 평균바람방향(QuickSCAT wind)은 2006년에는 남풍계열이 우세하였으나 2007년에는 동풍계열의 바람이 우세하여 양자강 저염수가 연구해역으로 수송되지 못하였다(KORDI, 2010). 여름철 영양염이 풍부한 양자강 저염수의 북동방향으로의 확산은 북부 동중국해에 분포하는 영양염의 주요 기원(Gong et al., 2004; KORDI, 2010)으로 알려져 있는데, SeaWiFs 해색위성(NOAA)에 의해 탐지된 조사해역에서의 7월 평균 클로로필 a의 농도는 2007년에 비해 2006년에 두 배 이상 높게 나타났다(Kim, 2008). 이러한 해양의 일차생산력의 차이는 식물플랑크톤을 섭취하는 동물플랑크톤의 생체량에 영향을 미칠 수 있으며, 동물플랑크톤을 주요한 먹이원으로 이용하는(Song per. com., Hokkaido Univ.) 살오징어 유생의 생존 및 분포를 결정하는 주요 원인으로 생각할 수 있다. 따라서 2006년의 환경조건은 2007년에 비해 유생의 생존에 유리하게 작용하였을 가능성이 크다. 향후 연구에서 이들의 분포를 결정하는 해류의 연변동성과 연계한 수송과정 연구와 함께 유생분포와 풍도의 연변동성을 결정하는 환경요인에 관한 연구는 이들의 시공간적 분포특성에 대한 이해를 넓히기 위해 필수적이라 생각된다.
살오징어 유생의 풍도는 성어 자원량 추정에 효과적인 지시자(indicator)로 알려져 있다(Murata, 1989; Sakurai et al., 2000; Goto et al., 2002). 현재 일본과 달리, 우리나라에서는 살오징어 유생에 관한 정기적인 모니터링이 이루어지지 않고 있다. 유생조사를 통해 살오징어 자원의 풍도를 예측하기 위해서는 우리나라 주변에서 살오징어 유생조사를 정기적으로 수행할 필요가 있다. 아울러 살오징어 유생의 분포 및 풍도에 관한 총괄적인 이해를 위해서는 동중국해와 동해 남부해역 등 광범위한 지역에서 산란하고 우리나라와 일본 해역을 경계 없이 이동하는 이들의 특성을 고려하여, 일본과의 공동조사 및 연구협력 프로그램 개발이 필수적이라 판단된다.