The feeding habits of
갯골은 간조시 담수와 해수를 연결하여 영양염류와 같은 에너지를 해양환경으로 공급하는 이동통로의 기능(Mitsch and Gosselink, 1993)을 가지고 있으며, 조수의 영향으로 인해 물리적 변화가 큰 생물학적 조건을 가진 생태계로 보고되었다(Haedrich, 1983; Kneib, 1997).
민물두줄망둑(
민물두줄망둑의 선행연구로는 샌프란시스코 하구역에 출현하는 민물두줄망둑의 식성연구(Scott and Larry, 2005)가 있었을 뿐, 국내에 민물두줄망둑의 생태에 관한 연구는 전무한 실정이다. 그리고 민물두줄망둑은 IUCN의 Red List에 최소관심종(Least Concern)으로 등재(IUCN Red List, 2009)되어 있어, 이들의 생태적 연구가 필요하다.
식성연구는 연구 대상종이 서식하는 생태계의 먹이망 구조(Huh and Kwak, 1998a)와 기능적인 면(Huh and Kwak, 1998b)을 파악하기 위한 정보를 제공하기에 중요하다. 그러므로 본 연구의 목적은 갯골에 출현하는 민물두줄망둑의 위내용물 분석을 통하여 주요 먹이생물, 크기와 계절에 따른 위내용물 조성, 섭식전략을 확인하여 특수한 환경에 성공적으로 적응한 민물두줄망둑의 식성을 알아 보고자 한다.
본 연구에 사용된 민물두줄망둑은 2011년 4월부터 2012년 3월까지 매월 1회 전라남도 순천시 해룡면 상내리 갯골에서 간조시에 입구 크기 가로 120 cm, 세로 100 cm, 전체 길이 225 cm, 그물코 크기 5 mm의 push net을 이용하여 5분씩 3회 예인하였다(Fig. 1). 채집된 민물두줄망둑은 현장에서 즉시 10% 중성포르말린에 고정하여 실험실로 운반한 후, 각 개체의 체장(standard length)과 체중(weight)을 각각 0.1 cm와 0.1 g까지 측정하였다(Fig. 2). 민물두줄망둑과 같은
위내용물 분석결과는 각 먹이생물의 출현빈도(%
%F= Ai/N×100
%N= Ni/Ntotal×100
%W= Wi/Wtotal×100
여기서,
먹이생물의 상대중요성지수(index of relative importance, IRI)는 Pinkas et al. (1971)의 식을 이용하여 구하였다.
IRI=(×100 %N+%W)×%F
상대중요성지수는 백분율로 환산하여 상대중요성지수비 (
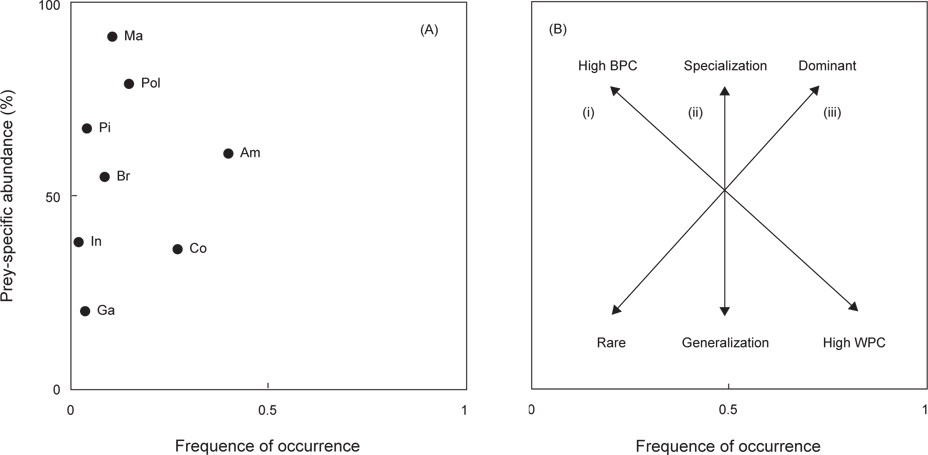
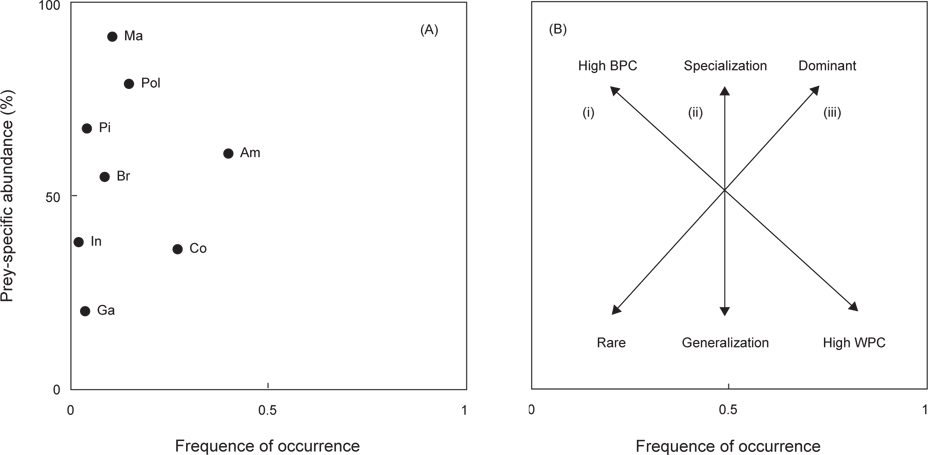
민물두줄망둑의 먹이중요도(dominant or rare), 섭식전략(specialist or generalist), 섭식폭(niche width)은 도해적방법(graphical method)을 사용하여 나타내었다(Amundsen et al., 1996). 이 방법은 출현빈도(
Pi=(ΣSi/ΣSti)×100
여기서,
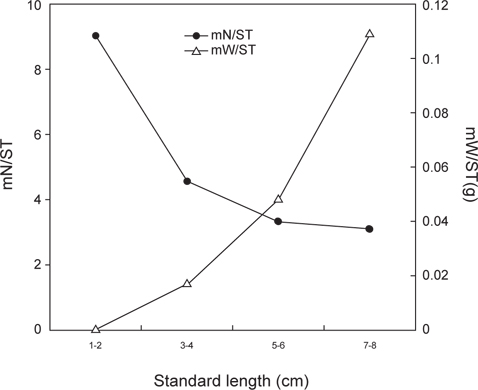
성장과 계절에 따른 민물두줄망둑의 먹이생물 변화를 파악하기 위해서 채집된 시료를 1 cm 간격, 4개의 크기군(1-2 cm, n=38; 3-4 cm, n=472; 5-6 cm, n=201; 7-8 cm, n=22)으로 나누었으며, 계절은 춘계(3-5월, n=134), 하계(6-8월, n=68), 추계(9-11월, n=365), 동계(12-2월, n=166) 사계절로 구분하여 총 733개체 중에서 위내용물이 발견된 706개체를 이용하여 먹이생물의 조성을 확인하였다. 크기와 계절에 따른 우점 먹이생물 조성의 통계적 차이를 분석하기 위하여 카이검정(x2-test)을 실시하였고, 체장에 따른 먹이섭식 특성을 파악하기 위해 크기 군별 개체당 먹이의 평균 개체수(mean number of preys per stomach,
SCI=SCW/BW×100
여기서
체장과 계절의 먹이생물의 중복도(Schoener, 1970)는 dietary overlap index를 이용하여 다음과 같이 구하였다.
Cxv=1-0.5(Σ│Pxi-Pxy│)
여기서,
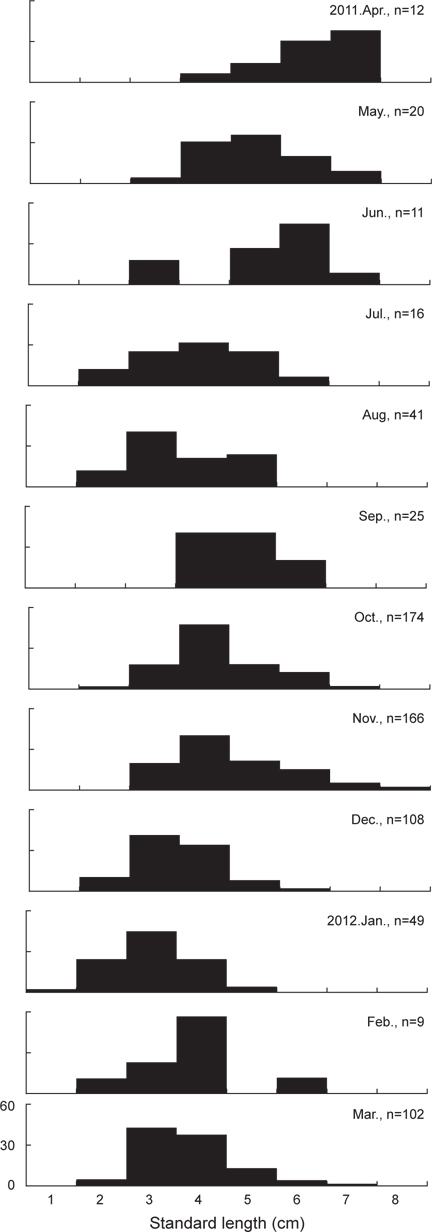
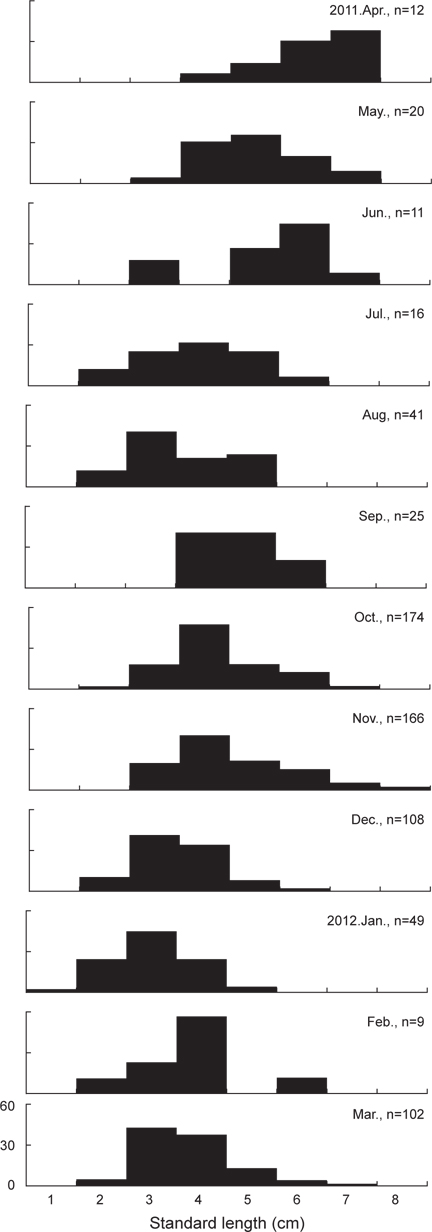
본 연구에 사용된 민물두줄망둑은 총 733개체로 체장(Standard length)은 1.5-8.2 cm의 범위를 보였으며, 3-4 cm 크기군의 개체가 전체 개체수의 57.6%를 차지하여 가장 많이 채집되었다. 1월에 가장 작은 크기인 1.5 cm 개체가 채집되었으며, 11월에 8.7 cm의 가장 큰 개체가 채집되었다(Fig. 3).
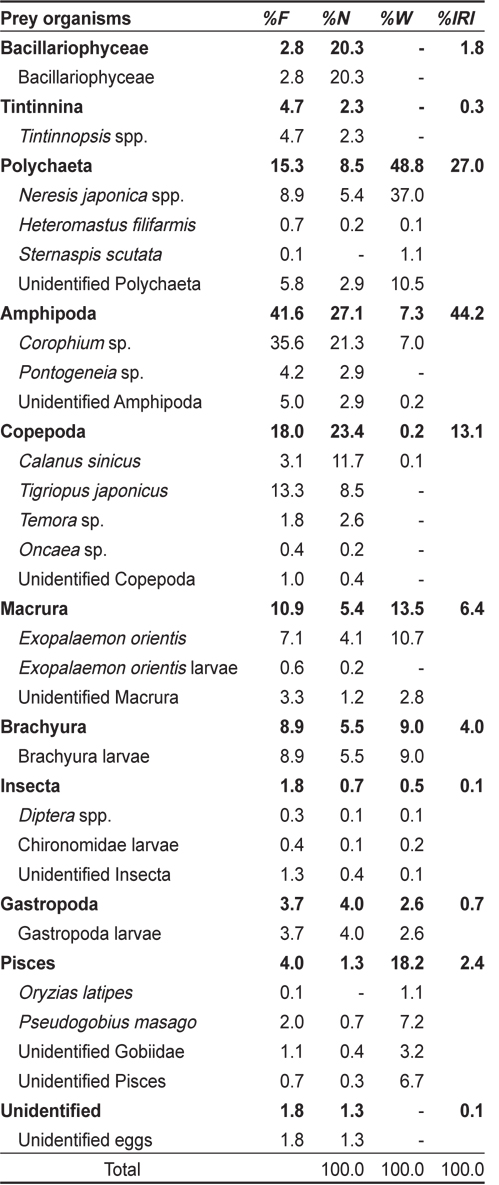
총 733개체의 민물두줄망둑 중 위내용물이 전혀 발견되지 않은 개체는 27개체로 3.7%의 공복율을 나타냈다. 위내용물이 발견된 706개체의 위내용물 분석결과(Table 1), 민물두줄망둑의 가장 중요한 먹이생물은 출현빈도 41.6%, 개체수비 27.1%, 습중량비 7.3%, 상대중요성지수비 44.2%를 차지한 단각류(Amphipoda) 였다. 단각류 중에서는
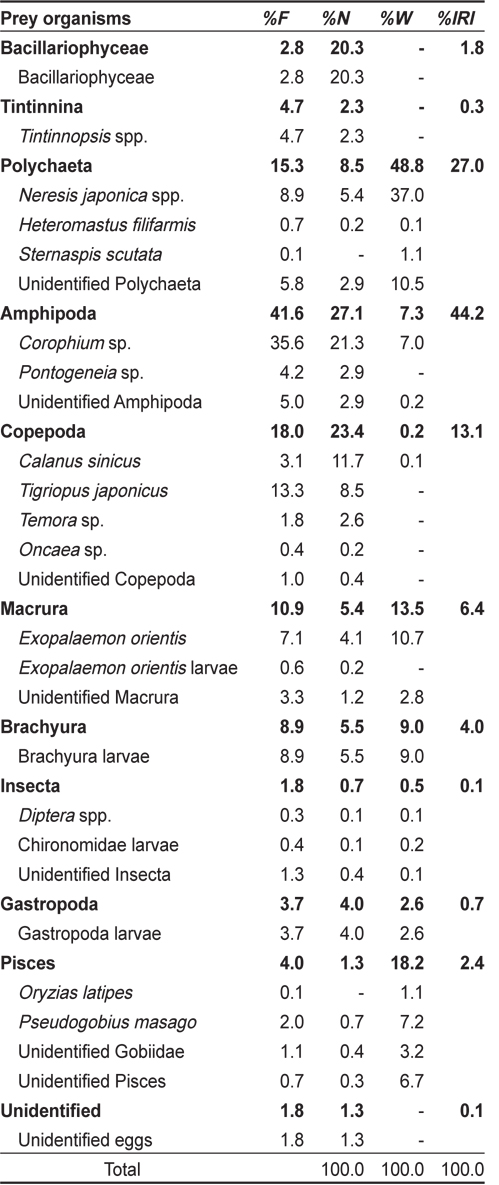
Composition of the stomach contents of Tridentiger bifasciatus by frequency of occurrence, number, wet-weight and index of relative importance (IRI)
민물두줄망둑의 섭식형태와 섭식특성을 조사한 결과(Fig. 4), 비교적 단각류를 섭식하는 경향을 나타냈지만, 좁은 섭식폭에 비하여 새우류를 비롯한 갯지렁이류, 어류, 게류 등 다양하게 섭식하는 것으로 보아 generalists임을 확인 할 수 있었다.
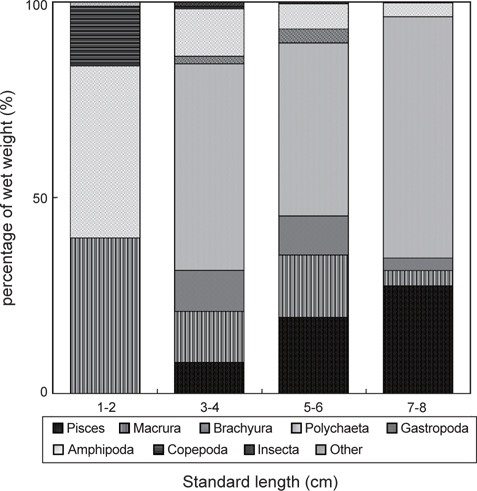
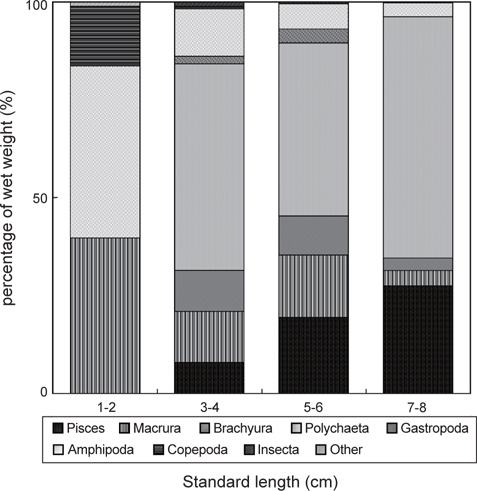
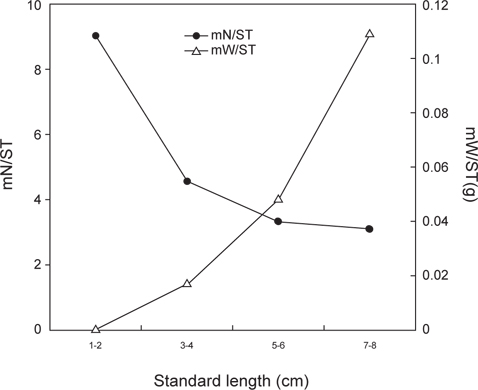
민물두줄망둑의 성장에 따른 먹이조성의 변화 분석한 결과(Fig. 5), 1-2 cm인 가장 작은 크기군에서는 단각류와 새우류가 각각 습중량비 43.9%와 39.8%로 가장 중요한 먹이 생물이었으며, 그 다음으로 요각류가 습중량비 15.2%를 차지하였다. 3-4 cm 크기군에서는 1-2 cm 크기군에서 출현하지 않았던 갯지렁이류의 습중량이 52.8%로 가장 높게 나타났으며, 어류, 게유생, 복족류 유생 또한 출현하였다. 그러나 단각류와 요각류의 습중량은 12.1%와 0.7%로 급격히 감소하였으며, 새우류 또한 감소한 것을 확인 할 수 있었다. 5-6 cm 크기군에서는 갯지렁이류의 습중량이 44.2%로 가장 높았으며, 어류의 습중량은 19.5%로 증가하였다. 7-8 cm 크기군 또한 갯지렁이류의 습중량이 61.7%로 가장 높았다. 어류 또한 27.6%로 비교적 증가하였지만, 단각류, 새우류, 게 유생의 경우 습중량이 5% 이하로 감소하였으며, 복족류 유생과 요각류는 출현하지 않았다.
민물두줄망둑의 크기군별 먹이생물의 변화는 유의한 차이를 보이지 않았다(
크기군별 먹이중복도 결과(Table 2), 3-4 cm 크기군, 5-6 cm 크기군, 7-8 cm 크기군 사이의 값은 0.7 이상의 중복도 값을 나타냈으며, 3-4 cm와 5-6 cm 크기군 사이의 값이 0.84로 가장 높게 나타났다. 하지만 1-2 cm 크기군과의 사이의 값은 0.2 이하로 낮게 나타났으며, 가장 작은 크기군인 1-2 cm와 가장 큰 크기군인 7-8 cm의 사이의 값은 0.07로 가장 낮은 중복도를 보였다.
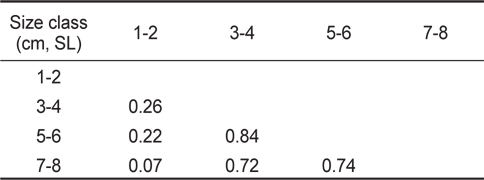
Proportional food overlap coefficients (Schoener's index) of the diet among Tridentiger bifasciatus size classes
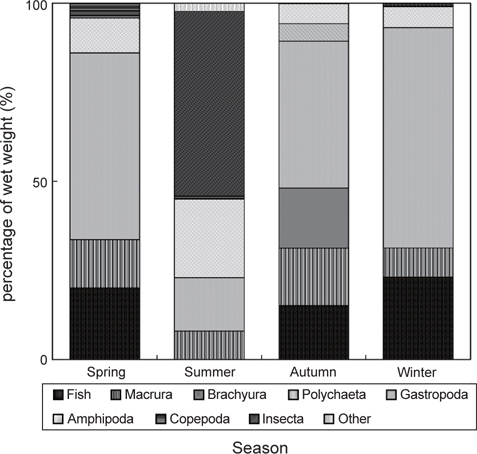
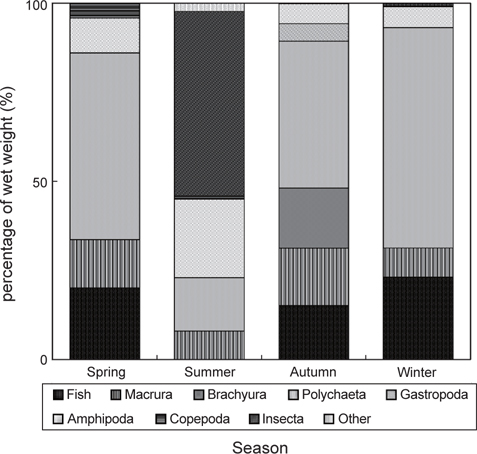
민물두줄망둑의 계절별 먹이조성 결과(Fig. 7)를 살펴보면, 하계를 제외한 춘계, 추계, 동계에는 갯지렁이류의 습중량비가 전체 먹이생물 습중량비의 40% 이상을 차지하여 가장 중요한 먹이생물이었으며, 어류를 비롯한 새우류, 단각류, 요각류가 지속적으로 출현하여 유사한 먹이조성 결과를 보였다. 하지만 추계에 경우, 게 유생과 복족류 유생이 민물두줄망둑의 먹이생물로 나타났으며, 그 습중량비는 전체 먹이생물 습중량비의 16.2%와 4.9%를 차지하였다. 하계에 경우에는 갯지렁이류의 습중량이 14.9%로 현저히 감소하는 한편, 곤충류의 습중량이 51.8%를 차지하여 가장 중요한 먹이생물이었으며, 단각류의 습중량비 또한 22.0%로 비교적 증가하였지만, 어류는 섭식되지 않았다.
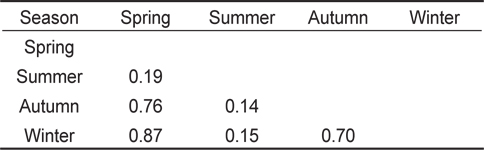
계절별 먹이중복도 결과(Table 3), 하계를 제외한 춘계, 추계, 동계의 사이의 값은 0.7 이상으로 나타났으며, 춘계와 동계 사이의 값이 0.87로 가장 높은 중복도 값을 보였다. 하계를 포함한 나머지 계절에 사이의 값은 0.19 이하의 값을 보였으며, 하계와 추계 사이의 값이 0.14로 가장 낮은 중복도 값을 나타냈다.
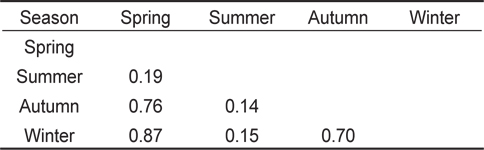
Proportional food overlap coefficients (Schoener's index) of the Tridentiger bifasciatus diet among season
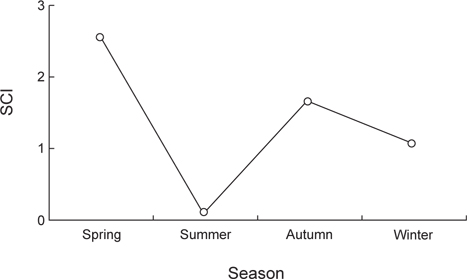
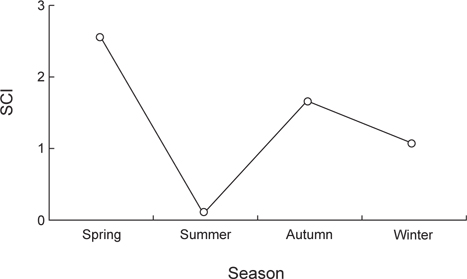
민물두줄망둑의 계절별 섭식율을 확인한 결과(Fig. 8), 춘계가 2.5로 가장 높은 섭식율을 보였으며, 하계는 0.1로 가장 낮은 섭식율을 나타냈다. 이후 춘계에 차츰 증가하였다가 동계에는 비교적 감소하는 경향을 나타냈다.
본 연구에서 민물두줄망둑은 다른 많은 망둑어과 어류와 마찬가지로 다양한 먹이생물을 섭식하는 generalists임을 알 수 있었다(Magnhagen and wiederholm, 1982; Miller, 1984; Mesa et al., 2008). 국내에 출현하는 망둑어과 어류의 공복율을 살펴보면, 점줄망둑(
민물두줄망둑의 가장 중요한 먹이생물은 단각류였는데, 단각류 중에서
크기군별 먹이조성의 차이를 확인한 결과, 전 크기군에서 유의한 차이를 보이지 않았다. 하지만 가장 작은 크기군인 1-2 cm는 단각류, 새우류 유생, 요각류를 주로 섭식하였는데, 이는 잘피밭에 출현하는 같은 망둑어과 어류인 점줄망둑 1-2 cm 크기군의 먹이생물과 유사하였다(Kwak et al., 2010). 가장 작은 크기군을 제외한 모든 크기군에서는 갯지렁이류가 중요한 먹이생물이었는데, Salgado et al. (2004)은 갯지렁이류가 단각류, 요각류와 비교 하였을 때, 더 높은 열량을 충족시키기 때문에 망둑어과 어류가 선호한다고 설명하였다. 본 연구지역은 주변 환경이 갯벌인 특성상 갯지렁이류의 풍부도가 높아 1-2 cm의 크기군을 제외한 나머지 3개의 크기군 모두에서 갯지렁이류의 비율이 높은 것으로 생각된다. 일반적인 어류에서 체장의 증가에 따른 먹이전환은 에너지 효율적 측면에서 일반화된 사실이며(Wainwright and Richard, 1995), 종내경쟁을 감소시키기 위해서 동일한 먹이라도 양을 달리하거나 먹이를 전환한다고 알려져 있다(Mesa et al., 2008).
계절에 따른 먹이조성을 살펴보면 하계를 제외한 전 계절에서 먹이생물의 중복도는 높은 값을 보였지만, 하계와는 낮은 중복도를 보였다. 이는 하계에 곤충류를 많이 섭식하였기 때문인 것으로 생각된다. 갯벌서식지와 지류에 출현하는 다른 망둑어과 어류의 위내용물에서도 곤충류가 출현한 것으로 보아(Hempel and Cattrijsse, 2004; Bob-Manuel, 2011), 담수의 유입이 있는 서식지에는 이 곳에 서식하는 어류들에게 곤충류가 비교적 쉽게 섭식되어지는 먹이생물인 것으로 생각된다. 곤충류 중 깔다구과 유생(Chironomidae larvae)의 비율이 높았는데, 본 조사지역은 직접적인 담수의 유입으로 인해 민물두줄망둑이 깔다구과 유생을 많이 섭식하였던 것으로 생각된다. 이러한 깔다구과 유생은 담수생태 내에서 중요한 구성요소로 보고되었다(Yoon and Chun, 1992). 계절별 섭식율을 조사한 결과, 하계에 0.1의 낮은 섭식율을 나타냈으며, 춘계와 추계에는 각각 2.5와 1.6의 높은 섭식율을 나타냈다. 하계에 가장 낮은 섭식율을 보인 이유는 어소를 보호하는 등 산란활동에 집중하기 위한 것으로 판단되며(Daoulas et al., 1993), 춘계와 추계에 섭식율이 높게 보인이유는 산란을 위한 높은 영양상태를 유지하고, 산란 후 부족한 영양을 회복하기 위한 것으로 생각된다. 이러한 경향은 같은 망둑어과 어류인 별망둑(