Many aquaculture businesses are intent not only on maximizing productivity and profitability, but also accomplishing this using environmentally responsible practices. Efficient use of energy (e.g., pumping of water) and natural resources (surrounding environment, ambient water supply, and waste streams) are key elements in this approach. Land-based recirculating aquaculture systems facilitate greater control over culture water and waste discharge than flow-through systems (Blancheton et al. 2009). Though the surrounding environment may be enhanced by moderate volumes of aquaculture discharge (White et al. 2011), the trend toward larger land-based facilities (e.g., 1,000 metric tons finfish production per year) and the associated effluent waste may pose a risk of local eutrophication. Alternatively, integration of seaweed and land-based marine finfish culture can convert these nutrients to a usable product. Previous investigations into land-based seaweed integration have included
Atlantic halibut [
For this study, integration of
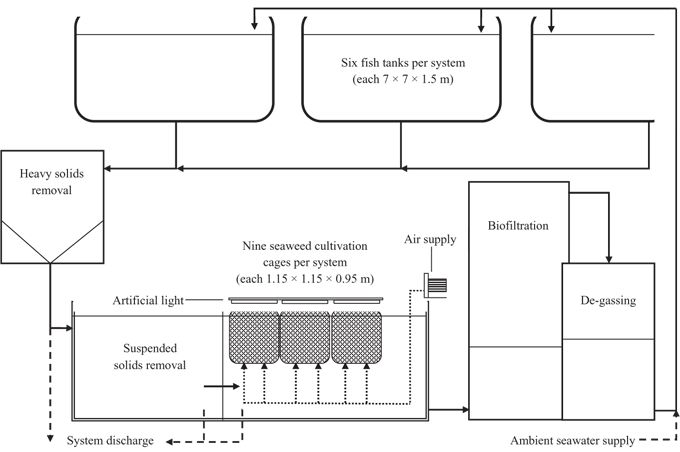
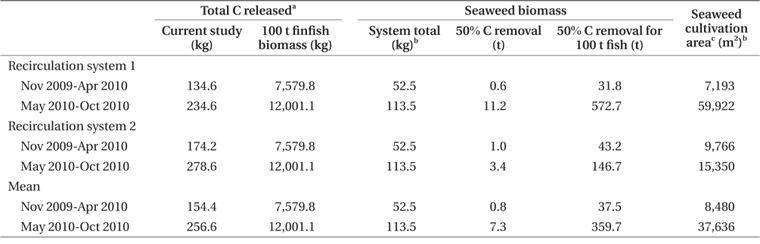
Total volume of each indoor commercial aquaculture system was approximately 500 m3 with a holding capacity of 6-8 t of Atlantic halibut. Solids filtration was accomplished by a swirl separator for removal of heavy waste, followed by diffused air flotation for removal of suspended solids. The seaweed was held in cages that were 0.95 m deep with a surface area of 1.32 m2 and volume of 1.25 m3 constructed of extruded plastic mesh (7 × 7 mm mesh size; Vexar, DuPont, Toronto, Canada). The cages were suspended at the water surface within the waste water collection sump (4.9 × 5.7 × 1.65 m deep) of the recirculating systems (Fig. 2). Nine of these cages were used in each of the three culture systems.
To keep the seaweed in suspension, air was supplied beneath each cage using four regenerative blowers each of 4-4.5 hp. Air was delivered continuously via two pipes about 0.6 m apart with 1.2 mm diameter holes with 5 cm spacing. Air was supplied beneath the cages continuously such that two rows of air bubbles, approximately 0.6 m apart rose through the cages. Light was supplied by natural sunlight through a translucent tarp structure, supplemented with 32 W, 5,000 K white T8 fluorescent lamps (six bulbs per cage; Philips, Somerset, NJ, USA). The minimum and maximum irradiance at the water surface, was 100 and 1,460 μmol photons m-2 s-1, respectively (Li-193SA spherical sensor, Li-Cor, Lincoln, NE, USA). A 16 : 8 L : D photoperiod was maintained throughout the year using the fluorescent lamps controlled by an automatic timer. Salinity was 29 to 31 psu over the study period.
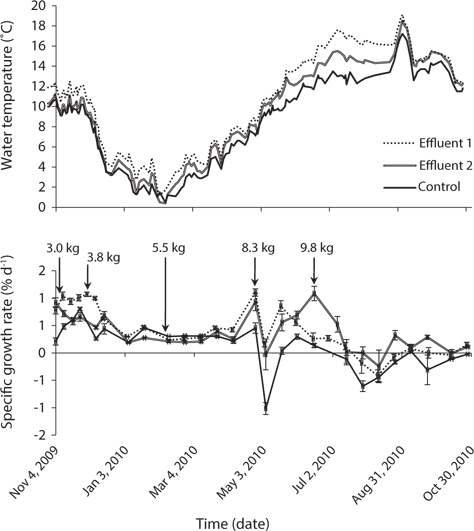
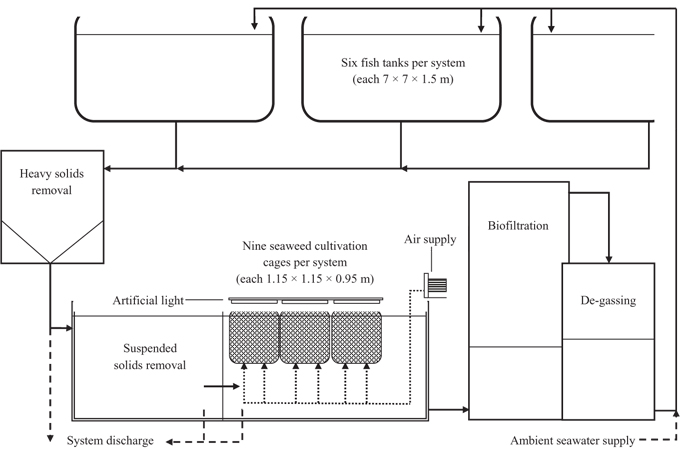
Water temperature ranged from 0.4°C in February, 2010 to 19.1°C in September, 2010 (Fig. 3). Temperature of Effluent 1 was about 1°C higher than Effluent 2 most of the year, increasing to about 2°C during June to August due to differences in water exchange rates between the systems. The rate of seawater inflow was governed according to fish biomass and feed loading of the system. The temperature of the control system ranged from 0.5°C in February and 17.2°C in September, and was on average 1.4°C cooler than the two effluent systems. The difference was greatest during June to August when water temperature of the control system was up to 3.6°C cooler than the effluent systems. The primary site of thermal losses and gains was carbon dioxide de-gassing by spilling over dispersal plates to maximize water surface exposed to the air within the effluent systems.
Mean pH of Effluent 1 and 2 systems was similar, 7.5 and 7.4 respectively, both ranging from 7.0 to 8.0. Control system pH was 8.0 on average (range 7.7-8.5).
Total nitrogen concentrations in all systems varied with season. From January to March, when water temperatures were sub-optimal for culture of Atlantic halibut, the appetite of the fish was low. This resulted in total nitrogen concentrations as low as 19 μM (standard error [SE], ±5.0) and 13 μM (±1.4) in Effluent 1 and 2, respectively. From May to October, when feeding rates were increased, mean total nitrogen concentrations were 126 (±24.4) and 166 μM (±27.8) for Effluent 1 and 2, respectively. Mean total nitrogen value of the control system was 18 μM (±4.4) between January and April, and 12 μM (±1.2) from May to October. Nitrate concentration as a proportion of total nitrogen averaged 81 to 88% for all three systems, originating from both nitrate in the natural environment, and the bacterial nitrification of ammonium excreted by the fish. Phosphorus concentration ranged from 2.6 μM (±0.03) in February to 17.7 μM in August PO43+ and 0.3 μM (±0.02) in March to 15.9 μM (±4.60) in October PO43+ for Effluent 1 and 2, respectively. Phosphate in the control system ranged from 0.2 μM (±0.08) in October to 5.5 μM (±2.82) in January PO43+. Mean nitrogen : phosphorus ratio in the water was 19.8 (±2.1), 23.4 (±1.7), and 24.9 μM (±2.2) for Effluent 1, Effluent 2, and control, respectively.
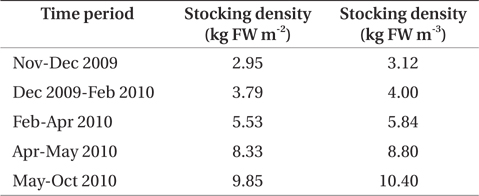
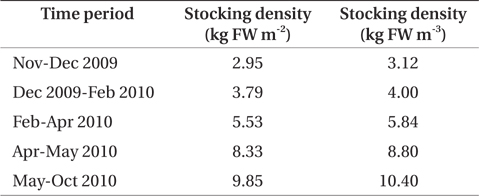
Stocking density of Palmaria palmata in seaweed culturing units at the land-based marine recirculating aquaculture facility of Scotian Halibut Ltd., Wood’s Harbour, Nova Scotia
DW and moisture content were determined by drying tissue samples to constant weight at 60°C. Using the biomass data collected bi-weekly, specific growth rate (SGR, expressed as % d-1) was calculated using the following equation:
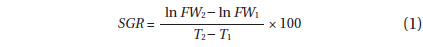
, where
Water samples were collected at 08:00, 14:00, and 20:00 the day following each biomass survey. Nitrate and nitrite concentration was determined by the nitrite cadmium reduction method (Jones 1984). The NH4+ concentrations were analyzed using the method of Liddicoat et al. (1975). Percent unionized ammonia was determined at different temperatures, pH, and salinities using the following equation (Bower and Bidwell 1978):
% Un-ionized ammonia = 100 [1 + antilog (pKa8(
Total ammonia nitrogen (TAN) was calculated using the tables in Bower and Bidwell (1978). The sum of nitrate, nitrite and TAN was considered as total nitrogen. Phosphate concentration was determined following Parsons et al. (1984). Availability of each nutrient was calculated as the mean of the three samples collected for any particular sample day.
Dried tissues were ground to powder with an MM301 ball mill (Retsch, Newtown, PA, USA), and then analyzed for total tissue N and carbon (C) using a CNS-1000 Carbon, Sulphur and Nitrogen Analyzer (Leco, St. Joseph, MI, USA). Total tissue N and C were determined for each biomass survey. N removal was calculated using the following equation after Kim et al. (2007):
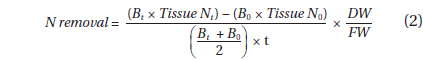
, where
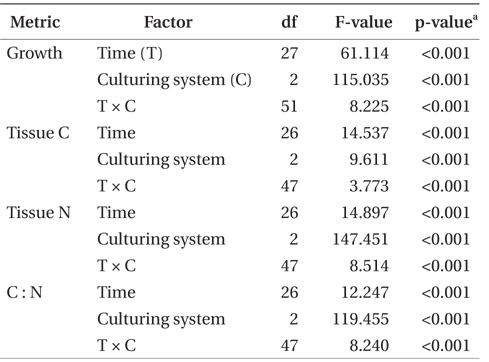
SGR, tissue nitrogen and carbon, and C : N ratio were examined as functions of the fixed factors time and culturing system using two-way ANOVA. Productivity was calculated based on the biomass increase per cage. Therefore, productivity was not statistically compared because the initial stocking density was increased as time progressed, affecting the productivity. C and N removal were not compared statistically since these were calculated values based on productivity and tissue C and N contents. Data were checked for homogeneity of variance prior to analysis. All data sets met this assumption. Post hoc analysis using Tukey’s honest significant difference test was used to make pairwise comparisons of treatment means when ANOVA indicated a significant treatment effect. All analyses were conducted using IBM SPSS Statistics version 20 (IBM Inc., Armonk, NY, USA).
Growth rate of
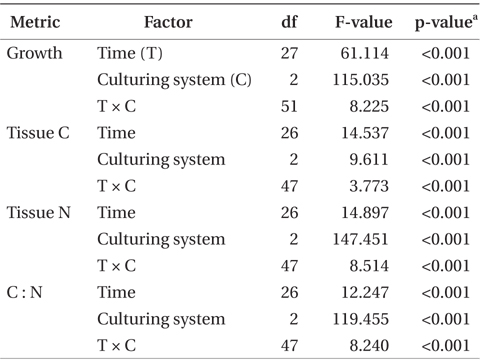
Effect of time and culturing system on growth rate (% d-1), tissue carbon and nitrogen (% dry weight) and C : N ratio
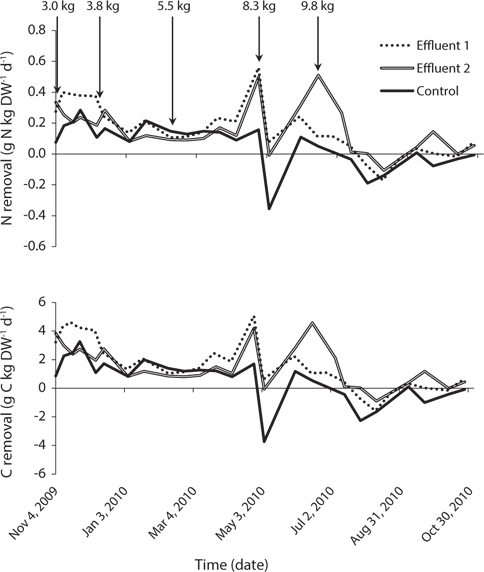
In the control system, growth rate was highest, 0.81% d-1, at around 10°C in November (Fig. 3). Growth rate was steady at about 0.3% d-1 from mid-December until the end of April, during which time temperature ranged from 0.5 to 9°C. When cage biomass was increased to 8.3 kg FW m-2 in April, growth rate decreased to a minimum of -1.10% d-1 over two weeks. Thereafter, during May-June, seaweed recovered to positive growth until biomass increased again to 9.8 kg FW m-2. Growth rate then declined to 0% d-1 in early July when temperature reached 13.5°C and remained at or below zero growth until the end of the experiment in October. Leading up to biomass increases to 8.3 and 9.8 kg FW m-2, the growth rate in both Effluent systems was 100% higher than that of the control. Moreover, growth in the control system was consistently lower than in either Effluent system from late-April until the end of the study.
Incidence of epiphytes on
>
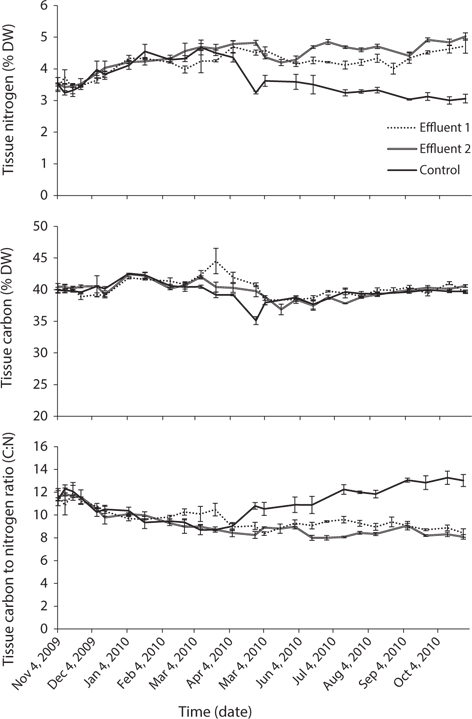
Tissue nitrogen, carbon, and carbon : nitrogen ratio
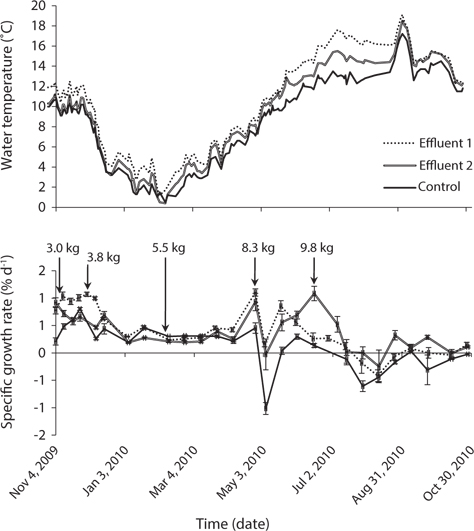
Seawater source (i.e., control or effluent), time, and the interaction of these two factors had an effect on tissue nitrogen content (p < 0.001) (Table 2, Fig. 4). Mean tissue nitrogen of
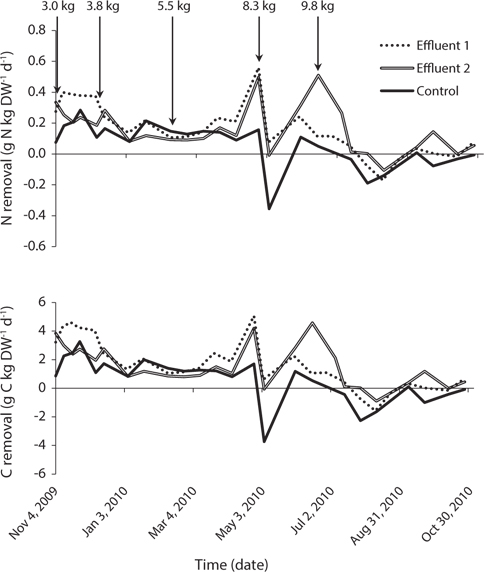
Nitrogen and carbon removal followed the same general pattern as growth rate (Fig. 5). Total nitrogen removal within each system remained stable from the start of the study in November 2009 until late-March 2010, ranging between 0.12-0.40, 0.09-0.34, and 0.08-0.28 g N kg DW-1d-1 in Effluent 1, Effluent 2, and control, respectively. Consistent total carbon removal was observed over the same time period with ranges of 1.05-4.61, 0.81-3.85, and 0.83- 3.27 g C kg DW-1 d-1 for Effluent 1, Effluent 2, and control, respectively.
Both nitrogen and carbon removal increased sharply near the end of April when water temperature was around 8°C in all systems, corresponding to a similar spike in growth rate. Nitrogen and carbon removal decreased following a biomass increase to 8.3 kg FW m-2, but subsequently increased in June as growth rate improved highest nitrogen removal rates of 0.56, 0.51, and 0.28 g N kg DW-1 d-1 for Effluent 1, Effluent 2, and control occurred in April, June, and November, respectively. Corresponding carbon removal during these same months was 5.06, 4.57, and 3.27 g C kg DW-1 d-1 for Effluent 1, Effluent 2, and control, respectively.
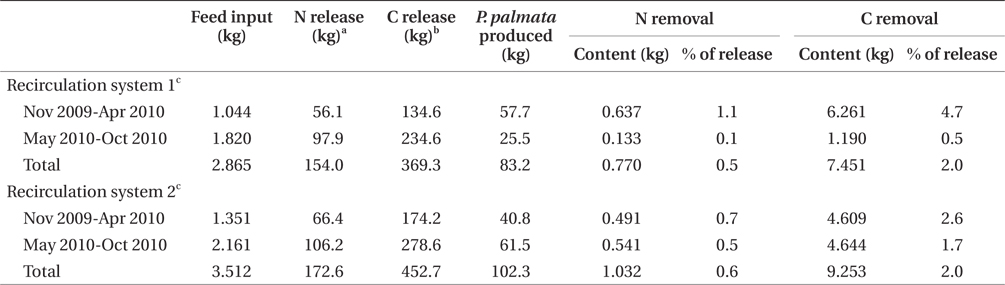
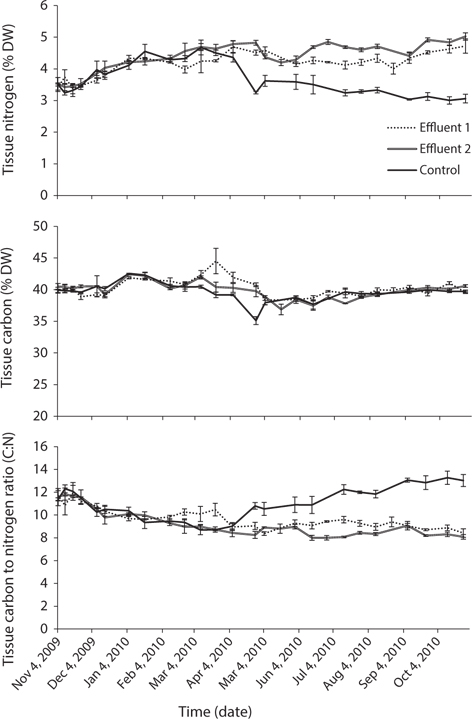
Nitrogen removal during winter, at mean seaweed system biomass of 52.5 kg, removed 1.1% of nitrogen released by feeding and fish metabolism in Effluent 1 (Table 3). The same result for Effluent 2 was 0.7%. During the same period, carbon removal was 4.7 and 2.6% of that produced by the fish in Effluent 1 and 2, respectively. Nitrogen and carbon removal efficiencies were lower during summer months for both systems. On average, both nitrogen and carbon removal were six times more efficient from November to April than during summer months (Tables 4 & 5).
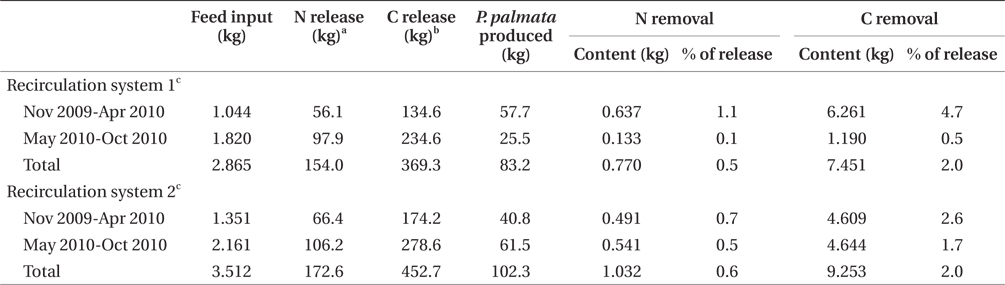
Nitrogen (N) and carbon (C) removal by Palmaria palmata relative to amount of feed available to Atlantic halibut in an integrated recirculating aquaculture system
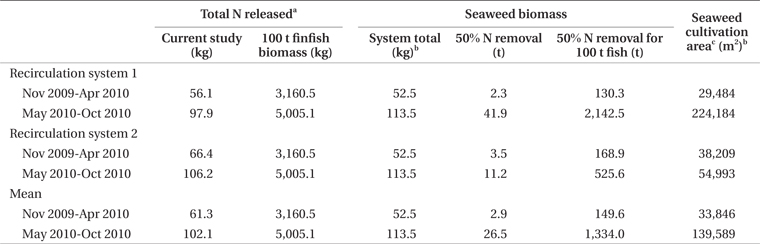
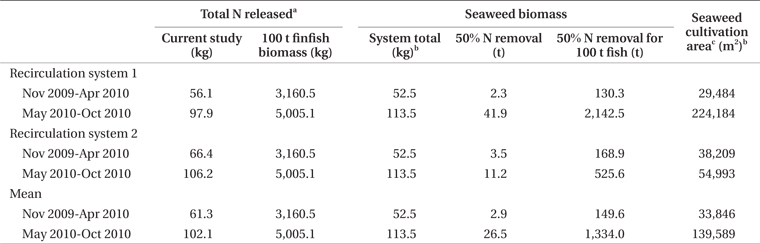
Required biomass and cultivation area of Palmaria palmata for nitrogen removal within integrated seaweed-halibut aquaculture
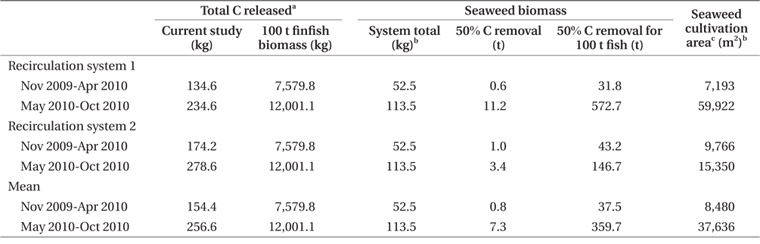
Required biomass and cultivation area of Palmaria palmata for carbon removal within integrated seaweed-halibut aquaculture
Clearly,
Seaweed cultivation area required for 50% carbon removal is 24 to 28% of the area required for the same removal of nitrogen (Table 5). The extent of integrated seaweed cultivation, then, may depend on the desired objectives. Carbon occurs in equilibrium as carbon dioxide (CO2), carbonate (CO32-), and bicarbonate (HCO3-) (Lobban and Harrison 1996), with CO2 readily dissociating to HCO3- at pH 7.8-8.2. Algal growth removes CO2 into organic matter by means of photosynthesis (Barsanti and Gualtieri 2006), though
Few studies have considered the carbon removal potential of seaweed species in seaweed-fish integration. In 2007, seaweed mariculture reached 1.4 million t in China, representing an estimated 0.34 million t of carbon removed from the marine coastal environment by seaweed harvesting (Tang et al. 2011). Investigating carbon dioxide fixation by microalgae in a photobioreactor, Otsuki (2001) found carbon removal rates of 13.75 g C m-2 d-1. Highest carbon removal rates in the current study were 8.00 and 10.94 g C m-2 d-1 for Effluent 1 and 2, respectively, similar to 11 g C removal m-2 d-1 by
Growth of
Matos et al. (2006) reported a yield of 40.2 g DW m-2 d-1 at 17-21°C for
Nitrogen removal by
Given the 1.5 : 1 seaweed : finfish ratio for 50% nitrogen removal in the current study, the challenge remains to reduce the surface area required for algal cultivation. Kim et al. (2013) determined that nitrogen removal by