Sleep is vital to maintain health and well-being because of its primary function of providing rest and restoring the body’s energy levels (Krueger et al., 2008). Sleep disorders impair not only cognitive and psychological functioning but also physical health (Brand and Kirov, 2011). In addition, obesity and cardiovascular disease are closely related to sleep disorders (Gangwisch et al., 2005; Wolk et al., 2005; Miller and Cappuccio, 2007).
Coffee is one of the most consumed beverages in the world (Tuomilehto et al., 2004), and has an arousal effect on the central nervous system (Davis et al., 2003). Caffeine, the major psychoactive constituent in coffee (Brown et al., 2001), induces wakefulness by inhibiting adenosine A2A receptors (Huang et al., 2005). According to the “Caffeine Intake by the U.S. Population” report, the average amount of caffeine consumed is approximately 300 mg/person/day (Somogyi, 2010). A large amount of caffeine may lead to sleep disorders, such as insomnia (Sanchez-Ortuno et al., 2005; Strassnig et al., 2006). Therefore, to identify foods, and their constituents, that can inhibit the arousal effect of caffeine would be promising from the perspective of the food industry.
In our previous studies (Cho et al., 2012a; Cho et al., 2012b), we have demonstrated that phlorotannin extracts from the brown alga
The high-purity phlorotannin preparation (HP-PRT, lot no. SD-GT-E-004) was obtained from S&D Co., Ltd. (Yeongigun, Chungcheongnam-do, Korea). HP-PRT was purified from the ethanol extract of the brown alga
All procedures involving animals were conducted in accordance with the animal care and use guidelines of the Korea Food Research Institutional Animal Care and Use Committee (permission number: KFRI-M-12027). Every effort was made to minimize the number of animals used and any pain and discomfort that they might experience. ICR mice (male; 23–28 g; 4 weeks) were purchased from Koatech Animal Inc. (Pyeongtaek, Korea). The animals were housed in an insulated, soundproof recording room maintained at an ambient temperature of 23 ± 0.5°C, with a relative humidity (55 ± 2%) on an automatically controlled 12-h light/12-h dark cycle (lights on at 5:00). They had free access to food and water.
Pentobarbital-induced sleep test
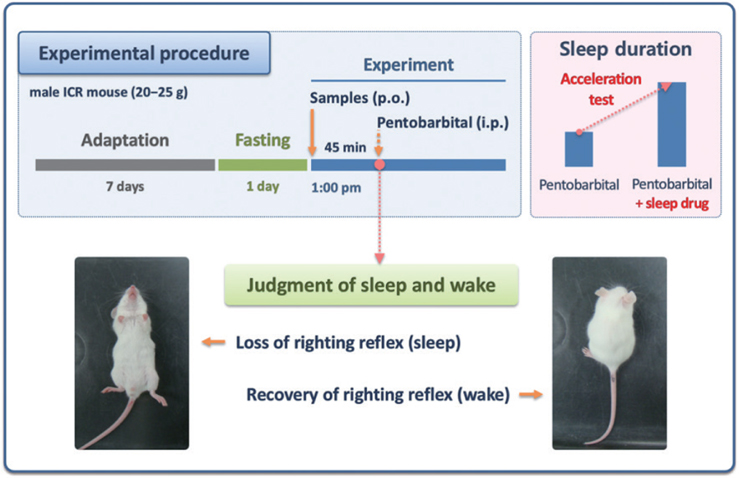
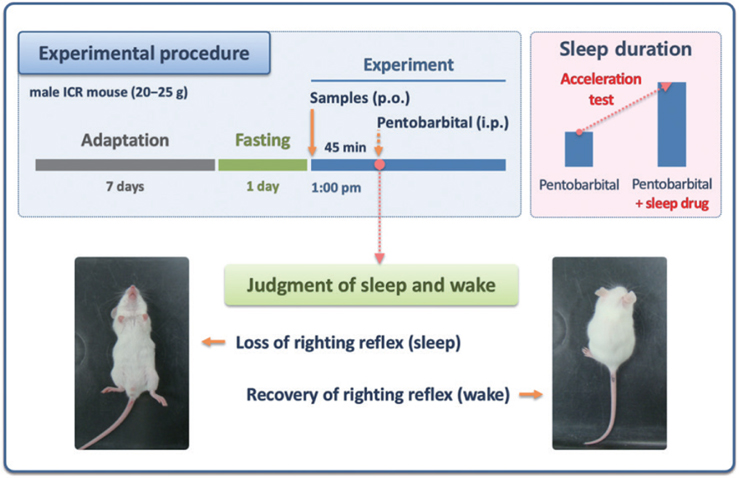
For better understanding the pentobarbital-induced sleep test in mice, the experimental procedure is shown in Fig. 1. All samples were dissolved in 0.5% (w/v) carboxymethyl cellulose (CMC)-physiological saline before use. Control mice (0.5% CMC-saline, 10 ml/kg) were tested in parallel with the animals receiving test sample treatment. All experiments were performed between 13:00 and 17:00, and mice were fasted for 24 h prior to the experiment. Test solutions were administered (post-oral injection, p.o.) to mice by using a sonde needle 30 min prior to the intraperitoneal injection (i.p.) of pentobarbital (42 mg/kg). Following the pentobarbital injection, the mice were placed in individual cages and observed for measurements of sleep latency and sleep duration. Observers were blinded to the individual treatments. The sleep latency was recorded from the time of pentobarbital injection to the time of sleep onset, and sleeping duration was defined as the difference in time between the loss and recovery of the righting reflex.
For multiple comparisons in the pentobarbital-induced sleep test, data were analyzed using one-way ANOVA followed by Dunnett’s test. Comparisons between group data were analyzed using an unpaired Student’s
Although a large amount of caffeine intake is known to lead to sleep disorders such as insomnia, few studies have been conducted on foods and their constituents with inhibitory effects on caffeine-induced wakefulness. In this study, we evaluated the effect of HP-PRT with sedative-hypnotic activity on caffeine-induced wakefulness by using the pentobarbitalinduced sleep test in mice. This method is useful to evaluate
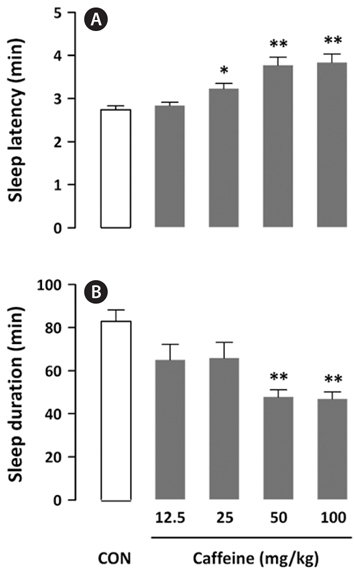
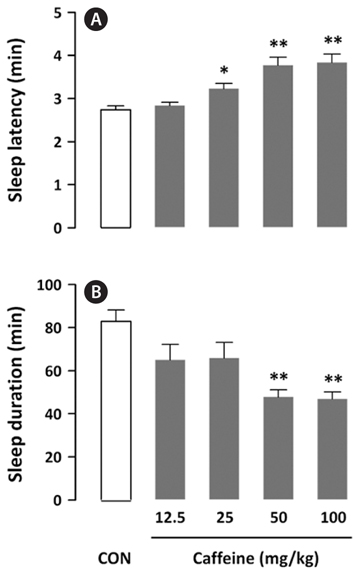
To determine a statistically significant dosage of caffeine, its effects on sleep latency and sleep duration in mice were evaluated. With a hypnotic dose of pentobarbital (42 mg/kg), caffeine (12.5, 25, 50, and 100 mg/kg, p.o.) produced a dosedependent increase in sleep latency and a decrease in sleep duration (Fig. 2). It showed a significant (
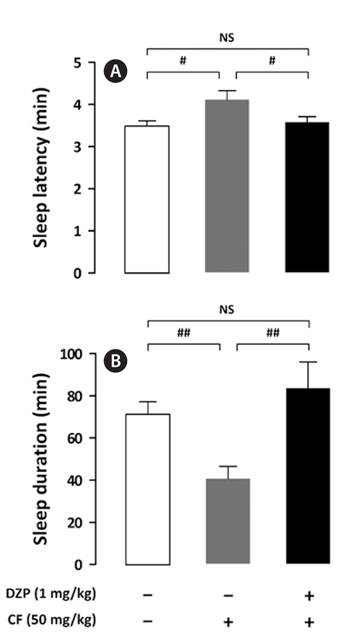
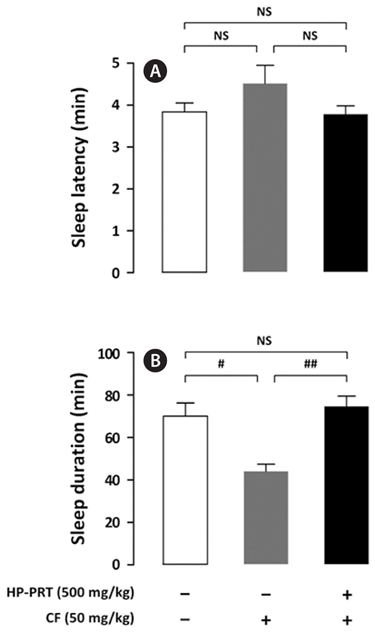
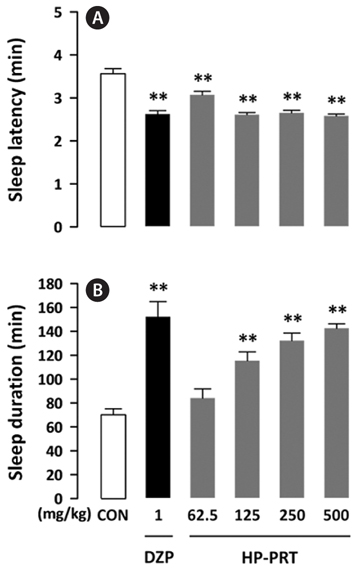
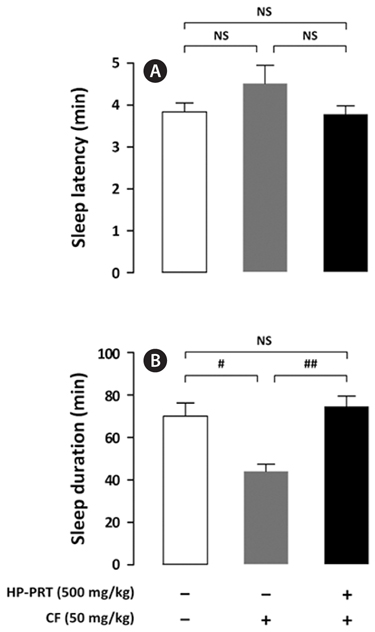
The inhibitory effect of DZP, a positive control on caffeineinduced wakefulness, is shown in Fig. 4. DZP induces sleep via positive allosteric modulation of the benzodiazepine site of the GABAA receptor (Kopp et al., 2003; Tobler et al., 2001). As expected, administration of caffeine (50 mg/kg) alone showed significant arousal effects; however, sleep latency and sleep duration of mice treated with both DZP (1 mg/kg) and caffeine were similar to those of the control group. HPPRT (500 mg/kg) also reversed the arousal effect of caffeine (Fig. 5). The pentobarbital-induced sleep test using rodents is a useful in vivo assay for evaluating hypnotic or arousal effect. Pentobarbital, an anesthetic, produces hypnosis and anesthesia by modulating the barbiturate site of GABAA receptor (Gerak et al., 2004). Regardless of molecular mechanisms of sleep-wake regulation, hypnotic or arousal compounds can accelerate or decelerate the pentobarbital-induced sleep. In this study, arousal effect of caffeine via the adenosine A2A receptor was offset by hypnotic effect of DZP and HP-PRT via the benzodiazepine site of the GABAA receptor. Therefore, this offset did not alter duration and latency of the pentobarbital-induced sleep. In addition, according to the report by El Yacoubi et al. (2003), caffeine reduced the hypnotic effect of alcohol by modulating the GABAA receptor (Koob, 2004; Lobo and Harris, 2008; Kumar et al., 2009).
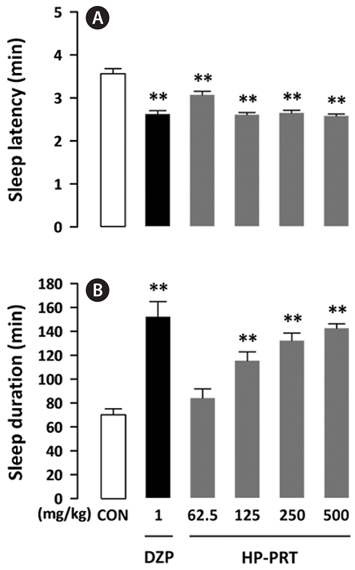
In this experiment, no adverse effects were observed following HP-PRT administration. It is known that benzodiazepine agents, such as DZP increase non-rapid eye movement sleep without altering rapid eye movement sleep (Qiu et al., 2009) but decrease delta activity, an indicator of sleep quality (Bastien et al., 2003). HP-PRT does not alter delta power (data not shown) and would thus be a better agent for inhibiting caffeine-induced wakefulness. Phlorotannins, which are oligomers and polymers of phloroglucinol (1,3,5-tri-hydroxybenzene), have a brown color and a bitter taste (Shibata et al., 2002). Their sensory characteristics would be suitable as an additive to coffee. In addition, phlorotannins have various biological activities, such as antioxidative (Zou et al., 2008), anti-inflammatory (Kim et al., 2009), antibacterial (Nagayama et al., 2002), and antiallergic (Sugiura et al., 2006) effects. Therefore, HP-PRT may be a good additive for the preparation of coffee products without an arousal effect. Further studies are needed to evaluate the effects of co-administration of HP-PRT and caffeine on sleep-wake architecture and profile.