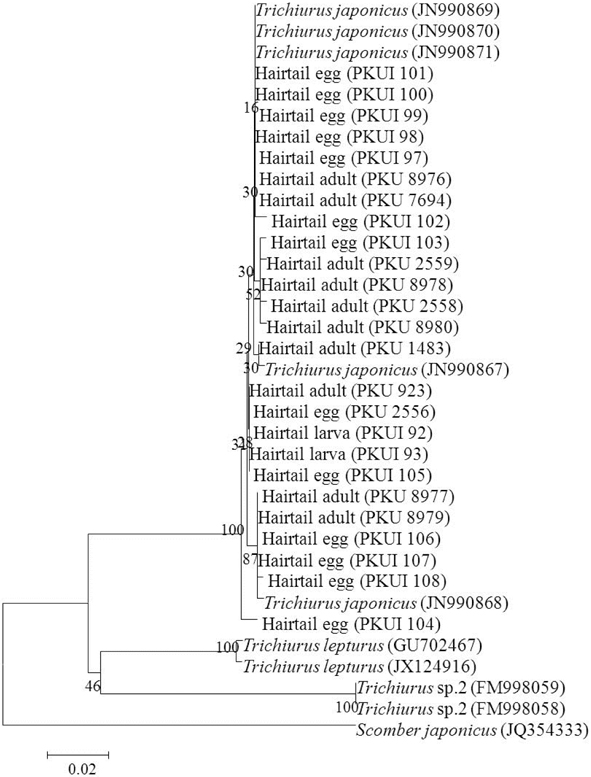
The hairtail (currently recognized as Trichiurus lepturus in Korea) is one of the most important commercial fish species in Korea, Japan, China, and Taiwan. Because the amount of catches has been steadily declining, we must determine the early life stages of the hairtail from the viewpoint of resource management. Furthermore, the taxonomic status of the hairtail is unclear among ichthyologists, potentially creating management difficulties. Therefore, the purpose of this study was to compare morphological and molecular information on eggs, larvae, and adults of hairtail from Korea with that of T. lepturus from the Atlantic Ocean, and to review the taxonomic status of the hairtail. A total of 510 base pairs of the mitochondrial DNA cytochrome oxidase subunit I sequences of 12 eggs, 2 larvae, and 11 adults of the hairtail from the Korean waters clearly matched those of Trichiurus japonicus adults (d = 0.000-0.014) from the East China Sea rather than those of T. lepturus (d = 0.100-0.110) from the Atlantic Ocean. Our results also showed that larvae of the Korean hairtail are different than those in the Atlantic Ocean in having no melanophores along the ventral edge of the lower jaw. Therefore, our findings suggest that the hairtail in the Korean waters may not be T. lepturus, but T. japonicus.
The hairtail (currently recognized as
Species in the genus
In Korea, the hairtail was first reported by Mori (1928) as
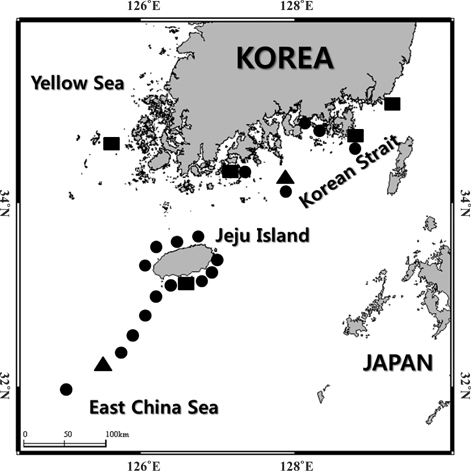
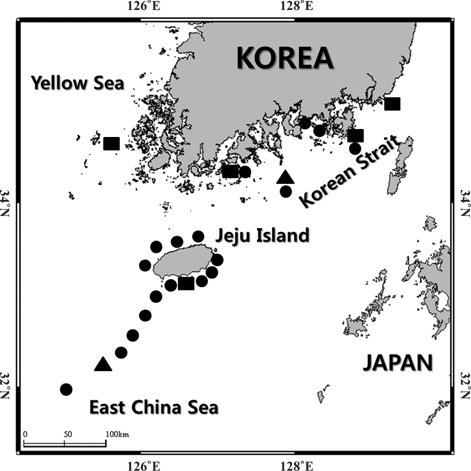
Hairtail eggs and larvae were collected mainly on Jeju Island, including the Korean Strait, between 2009 and 2012, using an ichthyoplankton net (mouth opening diameter of 0.8 m, mesh size of 330 μm) (Fig. 1). Samples were immediately preserved in 99% EtOH and RCL2 solution (RCL2-CS 1000; Alphelys, Plasir, France).
>
Morphological identification
Measurements followed the methods of Okiyama (1988) and terminology followed those of Russell (1976). Each body part was measured to the nearest 0.1 mm using the Image program (Active measure program, Korea). Morphological identification followed Okiyama (1988), Richards (2006), and Kim et al. (2011a). We used a camera (Moticam Pro 205A;Motic, Xiamen, China) attached to a stereomicroscope (SZX- 16; Olympus, Tokyo, Japan) to take photographs of eggs and larvae. Hairtail adults were collected on Jeju Island and the Korean Strait between 2008 and 2013, using a long-line and set-net method. Measurements of adults followed the methods of Nakabo (2002). Adult specimens were deposited at the Pukyong National University (PKU), and eggs and larvae were deposited at the Ichthyoplankton Laboratory of PKU (PKUI).
Genomic DNA was extracted from 12 eggs according to a method of modified Aranishi (2006). Genomic DNA was also extracted from eyeballs of two hairtail larvae, and from muscle tissues of 11 hairtail adults using 10% Chelex 100 resin (Table 1). A polymerase chain reaction (PCR) was used to amplify the mtDNA COI region using previously designed primers (Ivanova et al., 2007). The PCR conditions were as follows: initial denaturation at 94℃ for 2 min; 34 cycles of denaturation at 94℃ for 45 s, annealing at 53℃ for 1 min, and extension at 72℃ for 1 min 40 s; and a final extension at 72℃ for 7 min. The DNA was sequenced using an ABI 3730XL sequencer (Applied Biosystems, Foster City, CA, USA) and the ABI PRISM BigDye Terminator v 3.0 Ready Reaction Cycle Sequencing Kit (Applied Biosystems). For comparison, we obtained mtDNA COI sequences of
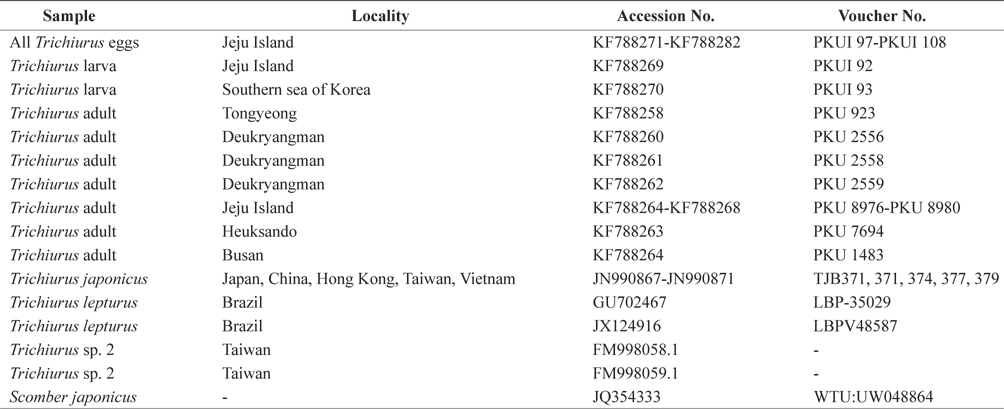
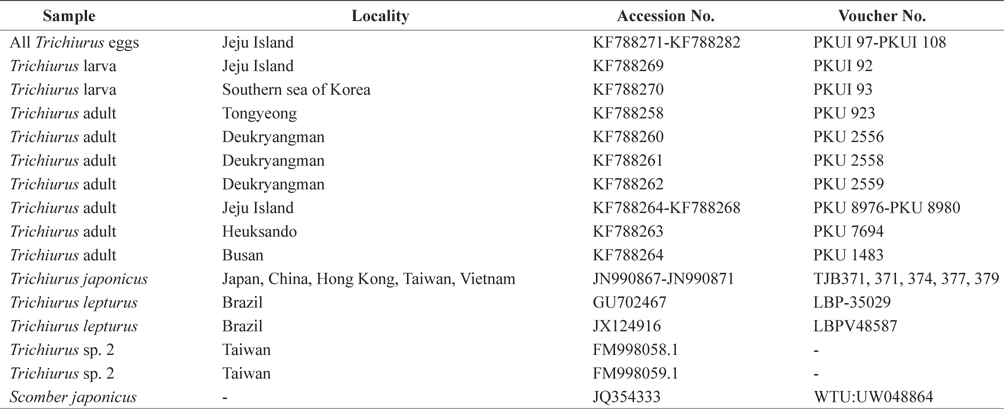
List of specimens of Trichiurus species including outgroup used for molecular analyses in the present study.
>
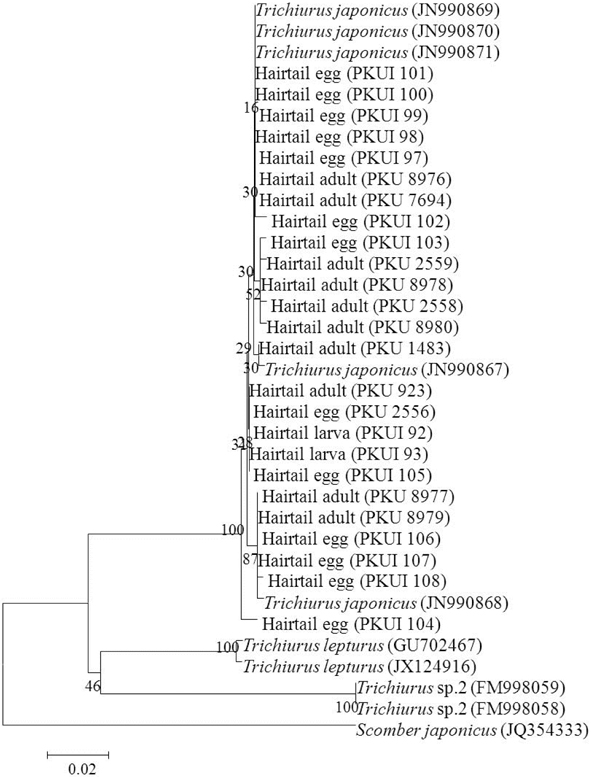
Molecular identification of eggs and larvae
Based on morphological characters, 291 eggs, 3 larvae, and 11 adults of the hairtail were identified as belonging to the genus
>
Morphological description of eggs and larvae
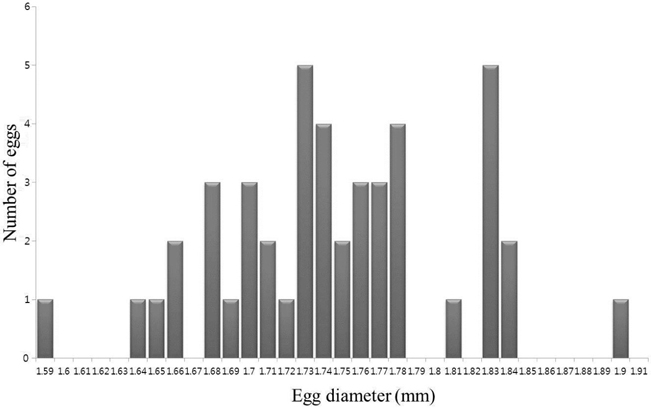
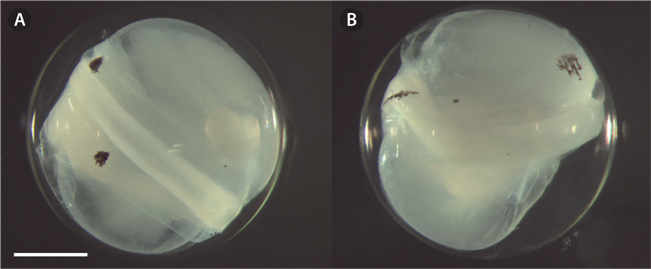
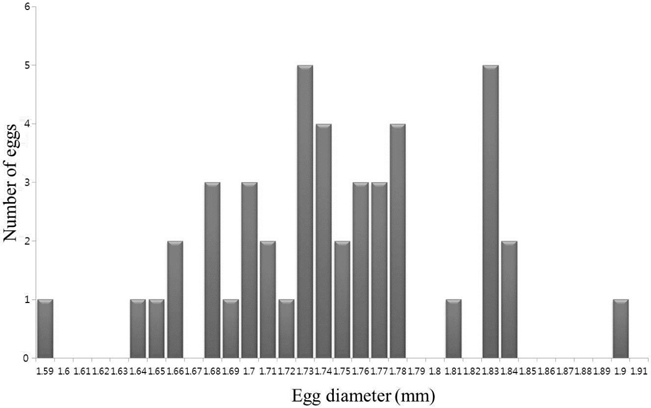
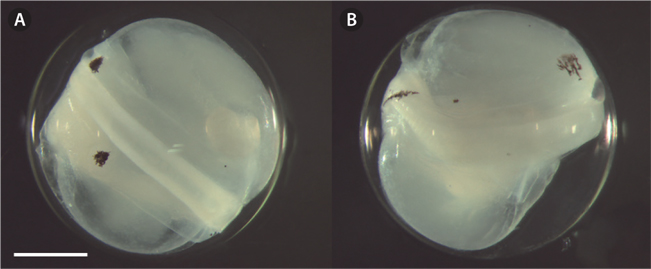
The eggs were spherical, with a narrow perivitelline space. The surfaces of the egg membrane and yolk sac were smooth and lacked structure. The mean egg diameter was 1.75 mm (range was usually 1.66-1.84 mm;
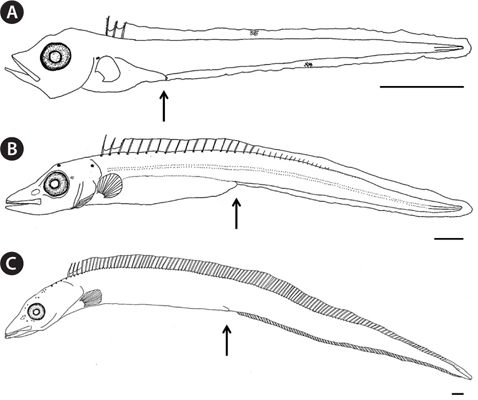
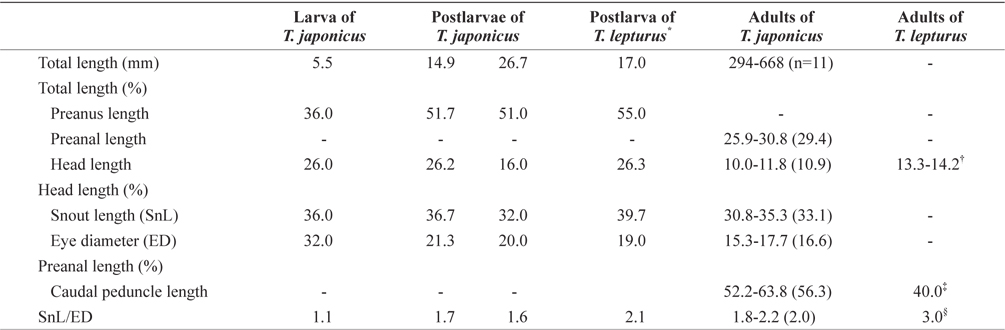
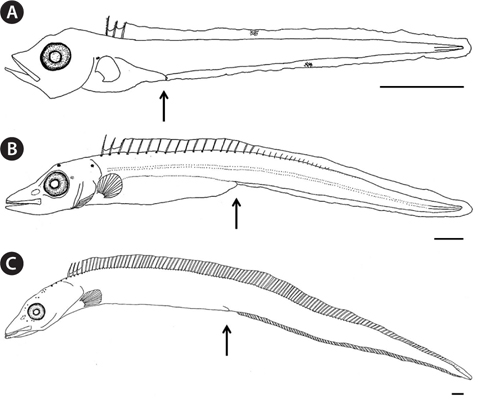
The proportional measurements of larvae and adults of the Korean hairtail are presented in Table 2. The larva (5.5 mm in total length [TL]) has a compressed and elongated body, and the anus was located on the anterior. Snout length and eye diameter were similar. Melanophores were faintly present, distributed on the dorsal body and on the dorsal and ventral fin folds. Three serrated dorsal spines were present on the anterior dorsal fin (Fig. 5A). The postlarva (14.9 mm TL) had a compressed and elongated body, and the anus was located around the middle of the body. The snout was slightly longer than the eye. Stellate-shaped melanophores first occurred on the dorsal head and opercle, and punctate- and stellate-shaped melanophores were found along the dorsal fin base. Silverwhite skin occurred slightly on the anterior body and was detached from the body after preservation (Fig. 5B). The postlarva (26.7 mm TL) resembled hairtail adult. The snout was slightly longer than the eye. Punctate-shaped melanophores also were observed on the dorsal head, and the tips of the jaws. And silver-white skin entirely covered the body. The caudal fin was absent (Fig. 5C). The adults (294-668 mm TL;
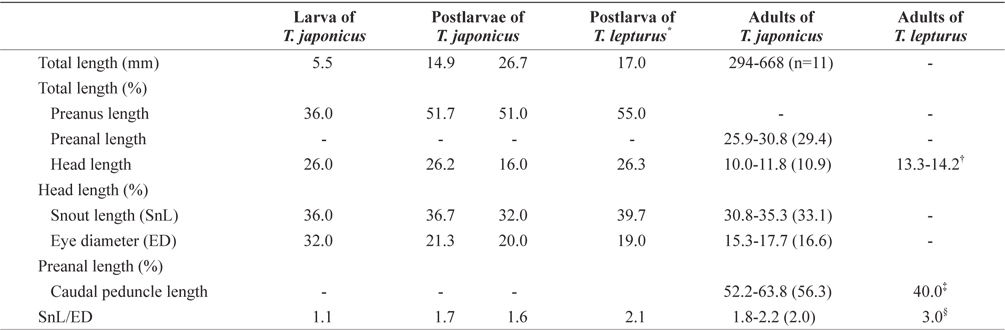
Proportional measurements of larvae and adults between Trichiurus japonicus and Trichiurus lepturus
In morphology, the hairtail larvae from Korea are very similar to those of
>
Taxonomic note on the hairtail in Korea
We compared the morphological characters of hairtail adults from Korea with reference data for
In the northwestern Pacific,