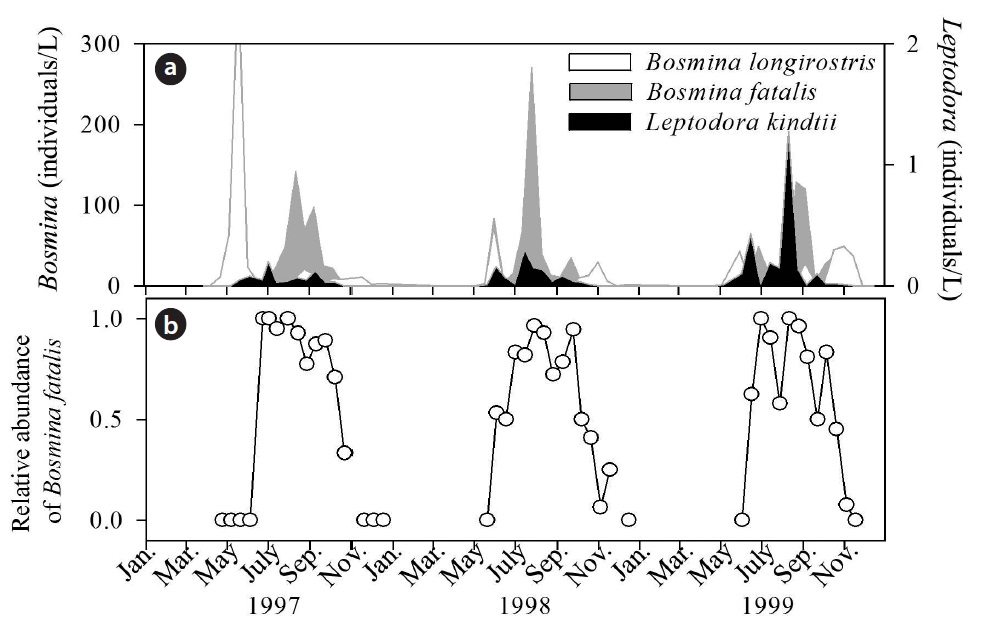
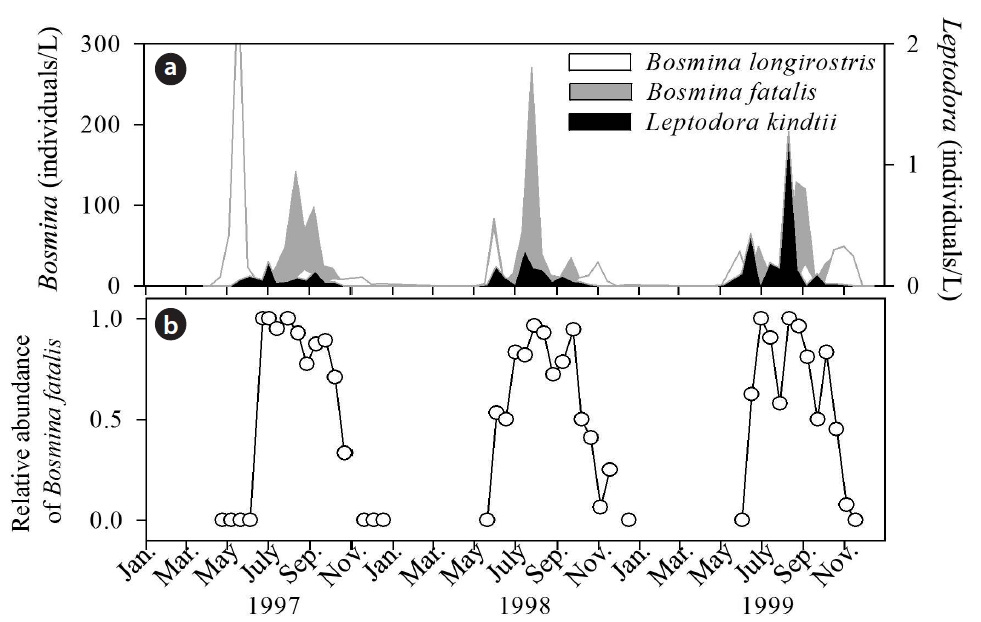
Natural communities in freshwater ecosystems show seasonal successions, which are caused by changes in environmental conditions and biological interactions (Connell and Slatyer 1977, Drake 1991, Murdock et al. 2010). Seasonal succession of two small coexisting bosminid cladocerans is commonly observed in Japanese eutrophic lakes, where Bosmina longirostris dominate the zooplankton community in spring and B. fatalis dominate in summer (Fig. 1).
There are two possible mechanisms explaining this succession pattern. The first is selective predation by Leptodora kindtii, which occurs in early summer (Chang and Hanazato 2003). The direct effect of temperature could not explain the seasonal succession of two Bosmina species (Hanazato and Yasuno 1985a), and thus Chang and Hanazato (2003) concluded that selective predation by L. kindtii on B. longirostris influenced the competition between two populations of Bosmina. The second is food density (Hanazato and Yasuno 1987). When the two species were reared together with the same initial numbers of individuals with a large food supply, B. fatalis overcame B. longirostris. The opposite occurred with a small food supply. These results indicate that the threshold food concentration (TFC), the ambient concentration of food that allows population losses to be compensated for by the production of offspring (Lampert 1977, Kreutzer and Lampert 1999), for B. longirostris is lower than that for B. fatalis. Moreover, a high population density of B. longirostris inhibits the subsequent increase of B. fatalis, indicating a density effect of B. longirostris on the population growth of B. fatalis. However, the importance of density effects in the seasonal succession of cladocerans has not
been experimentally examined.
Here, the question of whether a high population density of B. longirostris inhibits the growth of a B. fatalis population will be addressed. In the experiment, the summer decline of B. longirostris was simulated by preparing high and low density groups representing their seasonal populations. Food resources were also manipulated to examine whether the effect of B. longirostris on the population growth of B. fatalis is affected by food concentrations.
The stock culture of each species (single clone) was established from an individual collected in Lake Suwa (36°2′ N, 138°5′ E) a year before the experiment. The animals were maintained in a 20-L cylindrical polyethylene tank (30 cm diameter, 31 cm height) with 20 L of dechlorinated tap water for two weeks before the experiment. The tanks (one tank for each Bosmina species) were kept at a constant temperature of 22 ± 1 ˚C and a regime of 16 h light and 8 h darkness. The green alga, Chlorella vulgaris (Chlorella Industry Co. Ltd., Fukuoka, Japan; 2 × 109 cells), was introduced into tanks as food for Bosmina every second day. The culture medium in each tank was exchanged 24 h before the experiment, in which aggregated individuals near the light source (Bosmina shows positive phototaxis) were transferred to a new tank by pipetting. After the exchange of medium, no food was added into the tank until the start of the experiment.
The experiments were carried out in sixteen 2-L polymethylpentene beakers (Sanplatec Co. Ltd., Osaka, Japan) as experimental aquariums. To create conditions of high and low B. longirostris densities, 40 ml of the B. longirostris stock culture was added to 8 experimental beakers and 200 mL to the other beakers before the inoculation of B. fatalis, respectively. The numbers of individuals in three samples of the 40-mL and 200-mL stock culture media were counted, and the initial densities (mean ± SE) of B. longirostris (NBl-0) were 22.0 ± 1.0 individuals/L and 126.0 ± 8.4 individuals/L in low- and high-density conditions, respectively. The balance of each beaker was filled with dechlorinated tap water so that each contained 2 L. Experimental aquariums were assigned to low and high food concentrations of 0.5 × 105 and 5 × 105 cells/mL, respectively, every second day. All combinations of B. longirostris initial density and food concentration were run in four replicates. Experimental aquariums were maintained for eight days to increase populations of B. longirostris.
Eight days after the start of the experiments, the contents of the beakers were gently mixed, and 500 mL of water from each was sampled to estimate the density of B. longirostris (NBl-8). Each of the collected media was filtered through a 40 µm mesh net and preserved in a 4% sugar-formalin solution. After the sampling, B. fatalis was introduced into each beaker by adding 40 mL of gently mixed
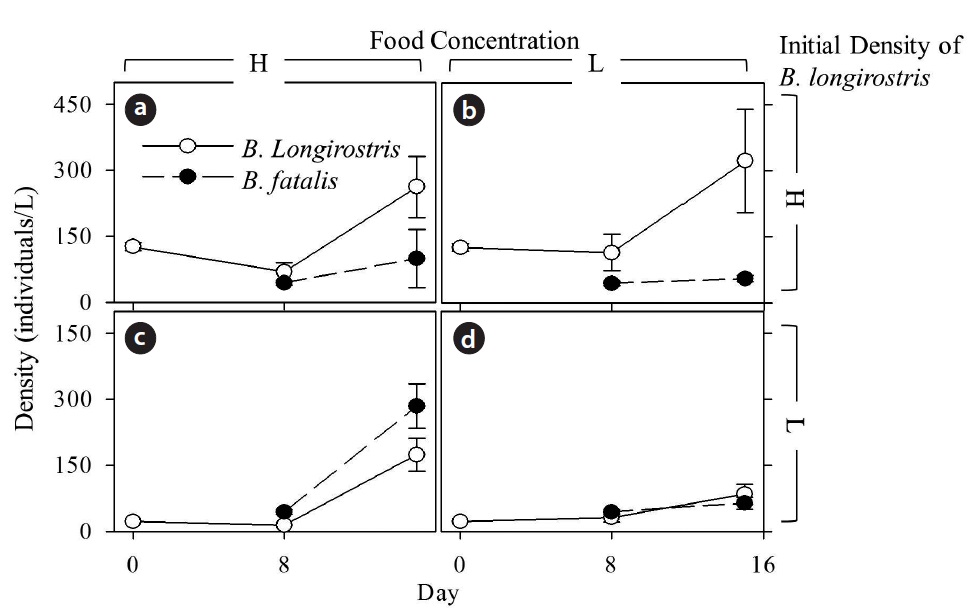
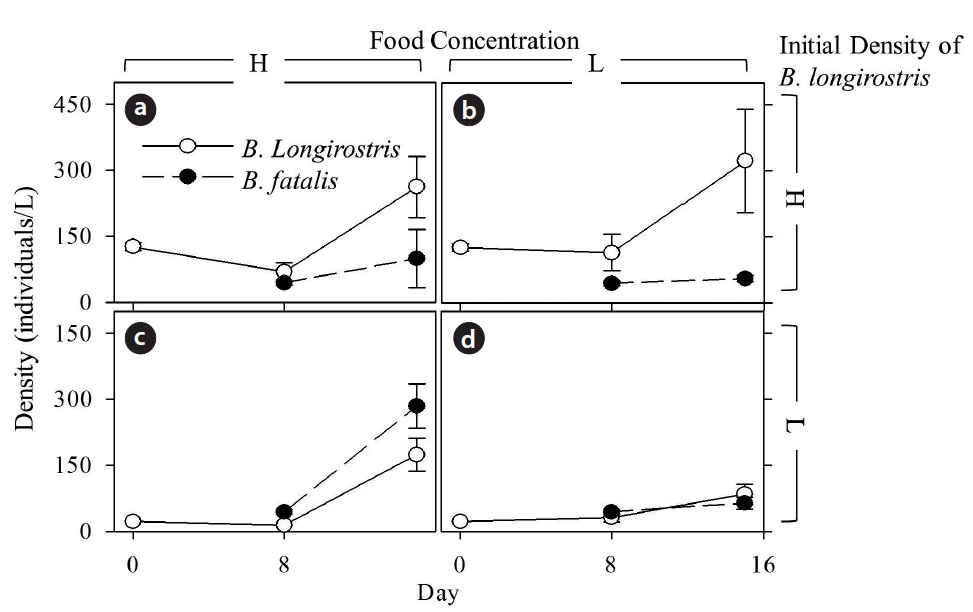
[Fig. 2.] Changes in density (mean ± SE) of Bosmina longirostris and B. fatalis cultured with high (a, c) or low (b, d) food concentrations, which is 5 × 105 or 0.5 × 105 cells/mL, respectively. Upper (a, b) and lower (c, d) figures show results of high and low B. longirostris initial density treatments, respectively. High- and low initial density of B. longirostris, are 22.0 ± 1.0 individuals/L and 126.0 ± 8.4 individuals/L, respectively. Open and filled circles indicate B. longirostris and B. fatalis, respectively.
stock culture medium. The density of B. fatalis (NBf-8) was estimated by counting the individual numbers in 40 mL of medium obtained from the other three samplings. Fifteen days after the start of experiments, the water remaining in each beaker (1.54 L) was filtered, and the samples were preserved in a 4% sugar-formalin solution. To estimate the densities of B. fatalis and B. longirostris at 15 days (NBf-15 and NBl-15), the number of each species in each sample was counted under a dissecting microscope and the total was recalculated into number per liter. The experiment was conducted under the same laboratory conditions as used for the stock cultures.
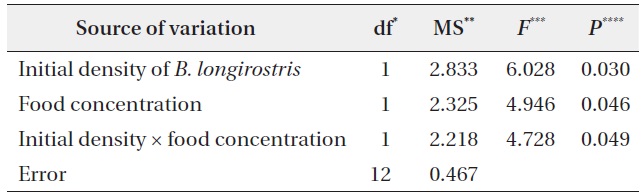
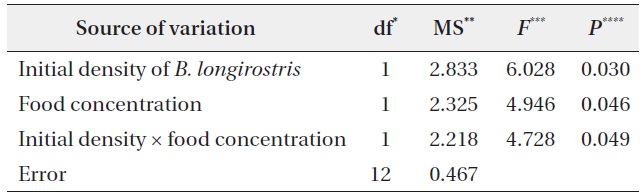
Population densities of B. longirostris and B. fatalis were analyzed to test the hypothesis. To confirm the B. longirostris density treatment and to examine the temporal change in its density for each condition, the population densities of B. longirostris at 8 and 15 days (NBl-8 and NBl-15) were analyzed with three-way repeated-measures ANOVA with time, initial density, and food concentration. The effects of B. longirostris population density and food concentration on the population density of B. fatalis (NBf-15) were tested with two-way ANOVA. To test increases in the population density of B. fatalis for each condition, a multiple comparison between the density of B. fatalis (NBf-8) and the B. fatalis density of each condition at 15 days (NBf-15) was performed using Dunnett’s test. Before statistical analyses, the densities of B. longirostris and B. fatalis were log-transformed to achieve variance homogeneity. Statistical analyses were conducted with R ver. 2.11.1 (R Development Core Team 2010).
Temporal changes in densities of B. longirostris and B. fatalis cultured with high and low food concentrations were shown in Fig. 2. Although density treatment and time significantly affected the density of B. longirostris, there were no effects of food concentration or interaction terms (Table 1). These results confirmed that the density of B. longirostris increased with time irrespective of food concentrations.
The NBf-8 value was 43.3 ± 4.6 individuals/L (Fig. 2). The values of NBf-15 were influenced by B. longirostris density, food concentration, and the interaction term between them (Table 2). Only the NBf-15 in the condition with low initial density of B. longirostris and high food concentration was significantly higher than all NBf-8 (Dunnett’s test, t = 3.718, P = 0.007). These results support the prediction that the growth of B. fatalis population was inhibited by the abundant B. longirostris. That B. fatalis increased only with a low B. longirostris density at a high food concentration suggests the presence of a density-dependent effect.
Results suggest that food competition can be a possible mechanism underlying the observed density-dependent effect of B. longirostris. In previously conducted competition
experiments, B. fatalis was depressed by B. longirostris at low food concentrations (Hanazato and Yasuno 1987, Chang and Hanazato 2004). Therefore, B. fatalis may have a higher TFC than does B. longirostris. In the present experiment, the algal concentration in the low-food conditions might be comparable with the TFC value for B. fatalis and higher than that for B. longirostris. In the condition with a high food concentration and a high density of B. longirostris, consumption by abundant B. longirostris individuals might decrease the algal concentration down to the TFC of B. fatalis. On the other hand, in the condition with a high food concentration and a low density of B. longirostris, B. fatalis was able to feed enough to reproduce. Previous studies showed that when they had the same initial density, the larger of two species was a better competitor than the smaller species under high food conditions because it could feed more (Romanovsky and Feniova 1985). As B. fatalis is larger than B. longirostris (Hanazato and Yasuno 1987), the feeding rate of B. fatalis may be higher than that of B. longirostris and thus be dominant under conditions with a high food concentration and a low density of B. longirostris.
The population shift from B. longirostris to B. fatalis is commonly observed during the early summer season in Japanese eutrophic lakes, as shown in Fig. 1 (Hanazato and Yasuno 1985b, Chang and Hanazato 2003). The present results demonstrate that the seasonal succession unconditionally requires very low population densities of B. longirostris. L. kindtii is an effective predator, which can reverse the dominance relationship between the two species through selective predation of B. longirostris (Chang and Hanazato 2003, 2004). In this study, laboratory experiments were conducted by using B. longirostris and B. fatalis collected from one site, Lake Suwa. Because B. longirostris is a cosmopolitan species and B. fatalis inhabits in East and Southeast Asia, they can present intraspecific variations in traits related to food competition. To test the effect of the intraspecific variations on the density effect of B. longirostris on the population growth of B. fatalis, further experiments by using the two species from other lakes are required. Investigating the density effect of species that are already present in a community on other species that appear in the community at some later time may help us to explain seasonal successions in zooplankton communities in natural lakes and ponds.