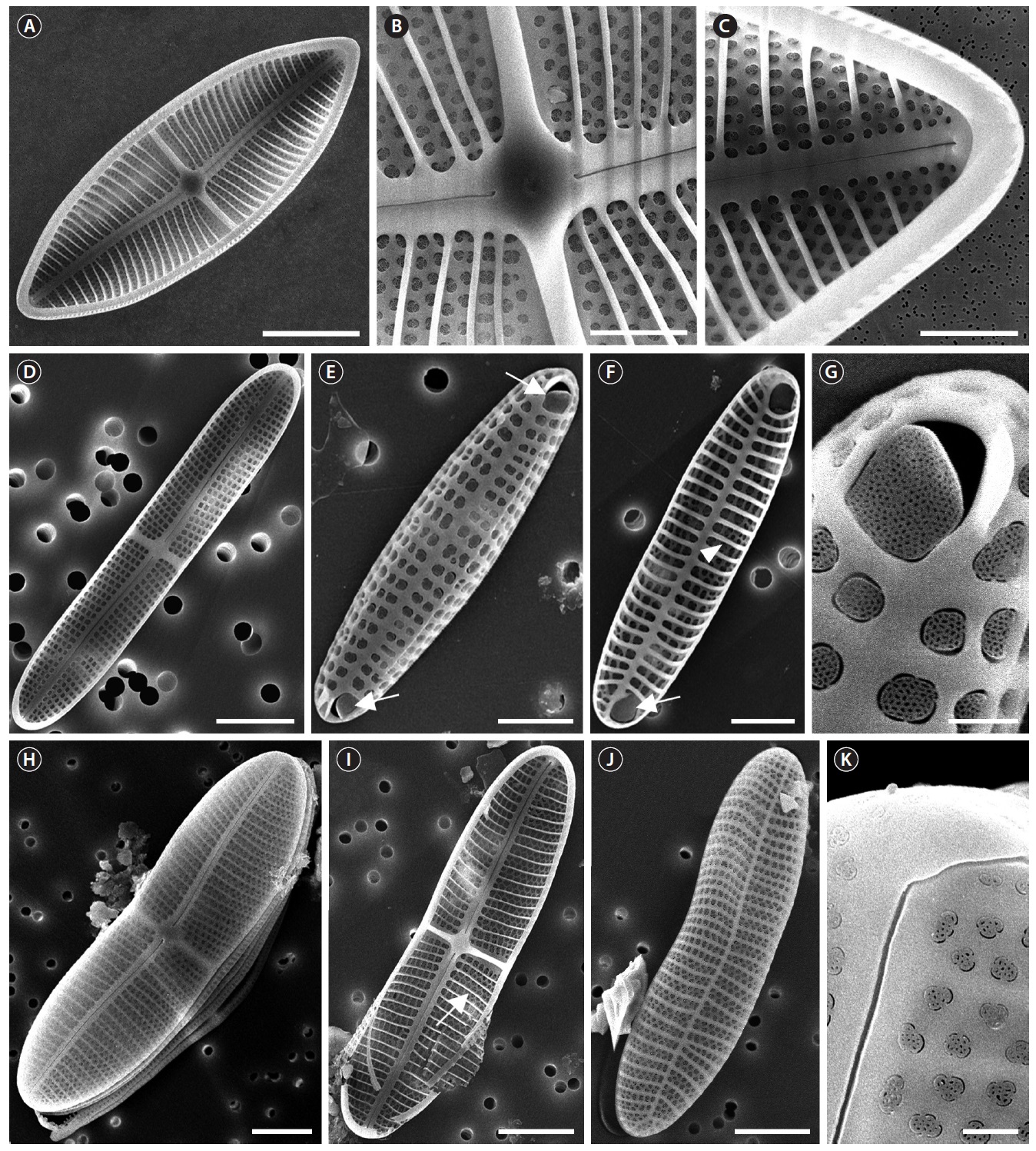
The genus
Toyoda et al. (2006) pointed out that there is no taxonomic information of the type and revised the type of the genus as
Thirty six taxa of
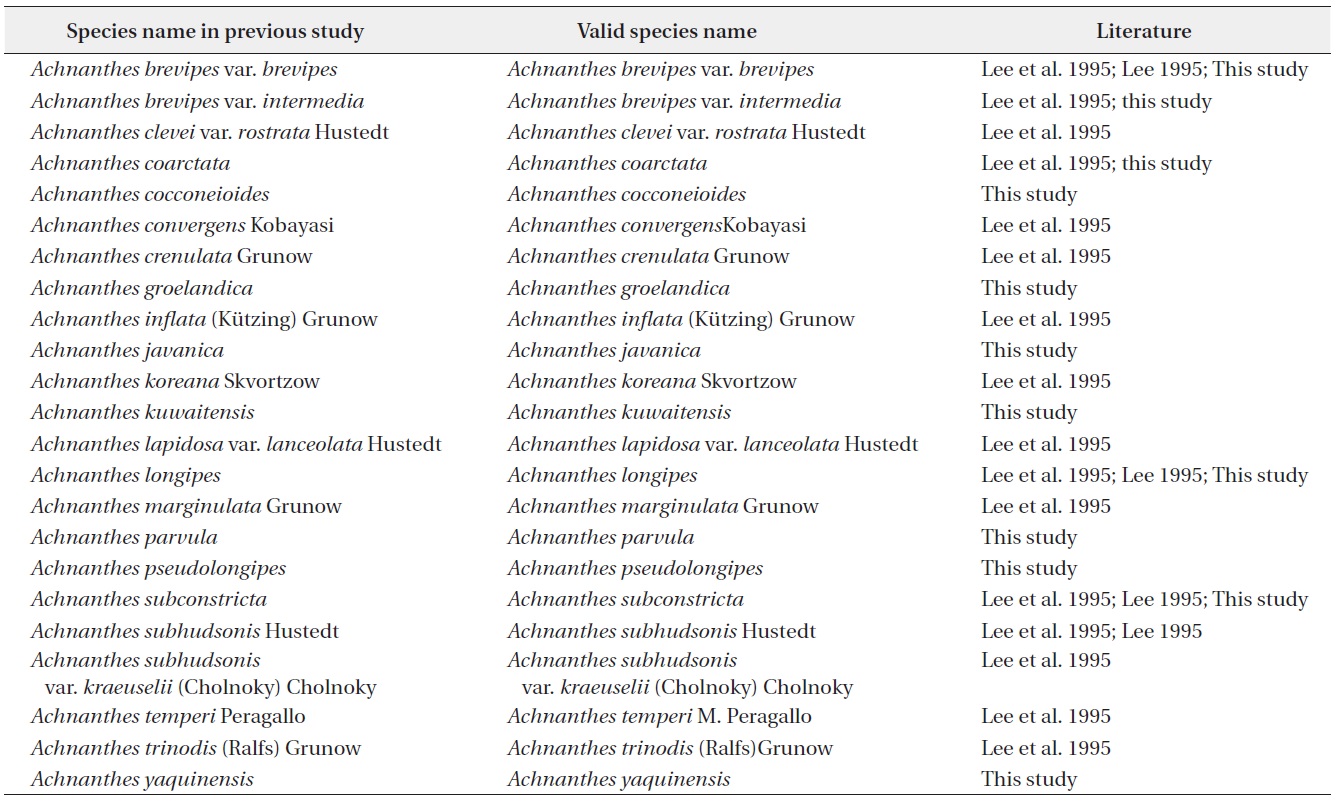
[Table 1.] List of Achnanthes species in Korea
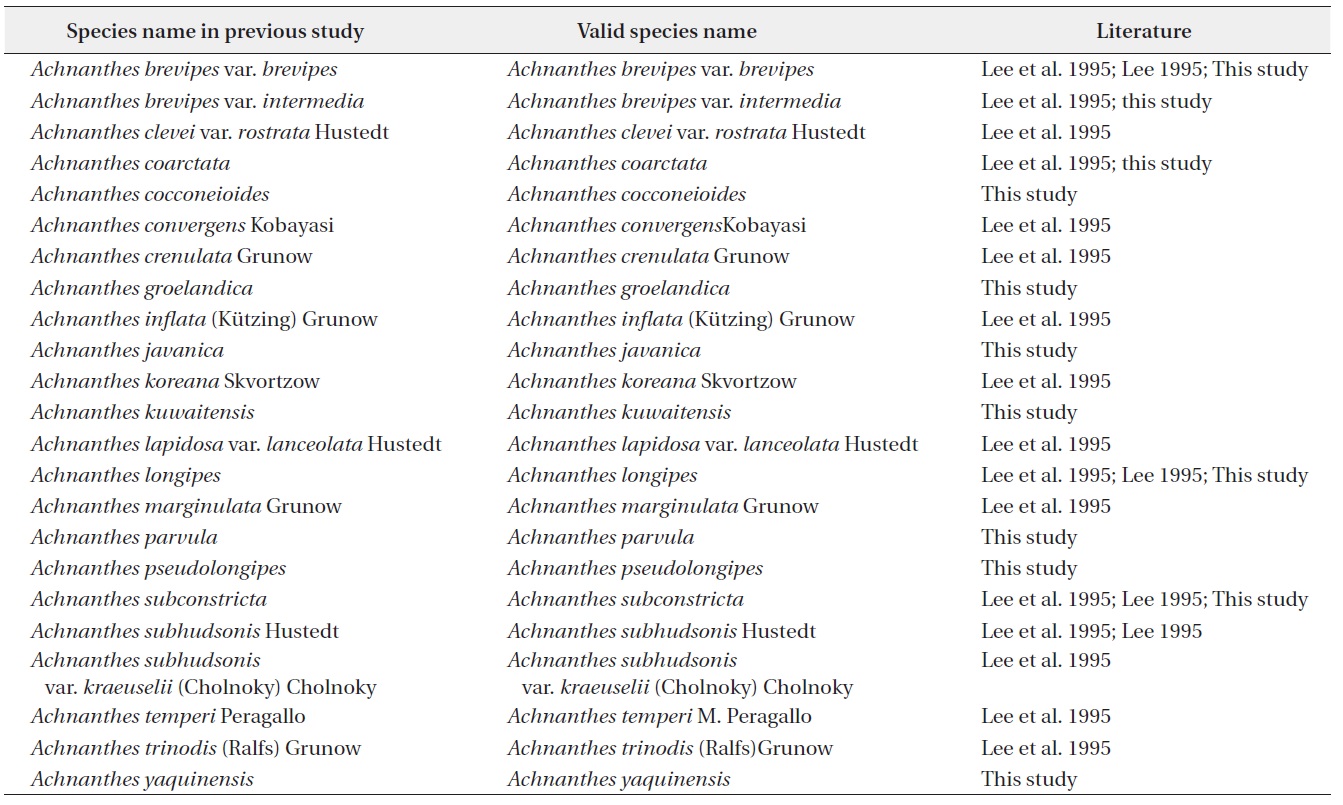
List of Achnanthes species in Korea
on each sample on a cover slip. The cover slip was put face-down on a slide glass and slightly heated using an alcohol lamp to evaporate the solvent. All specimens were observed using an Eclipse 80i light microscope equipped with bright-field and differential interference contrast optics (Nikon, Tokyo, Japan) with a DS-Fil digital camera (Nikon). Some cleaned specimens were individually dropped onto an aluminum stub and coated with gold-palladium. Coated samples were examined using a JSM-5600LV scanning electron microscope (JEOL, Tokyo, Japan). Sizes of the diatom species were analyzed with Axio Vision AC v. 4.5 image calculation software (Carl Zeiss, Oberkochen, Germany). Terminology followed general proposals by Anonymous (1975), Ross et al. (1979) and Toyoda et al. (2003).
Twelve
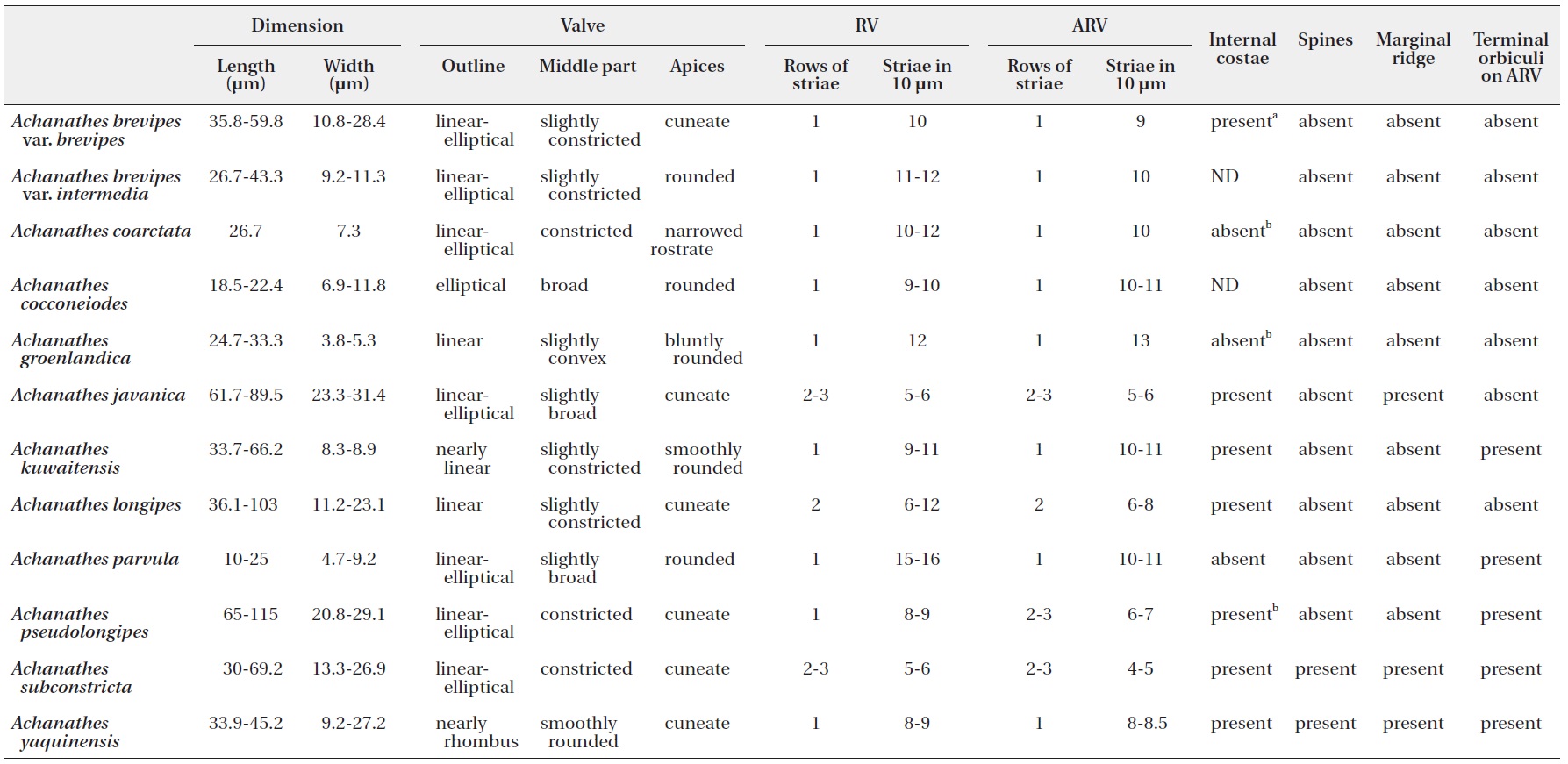
[Table 2.] Morphological characteristics of Acnanthes species
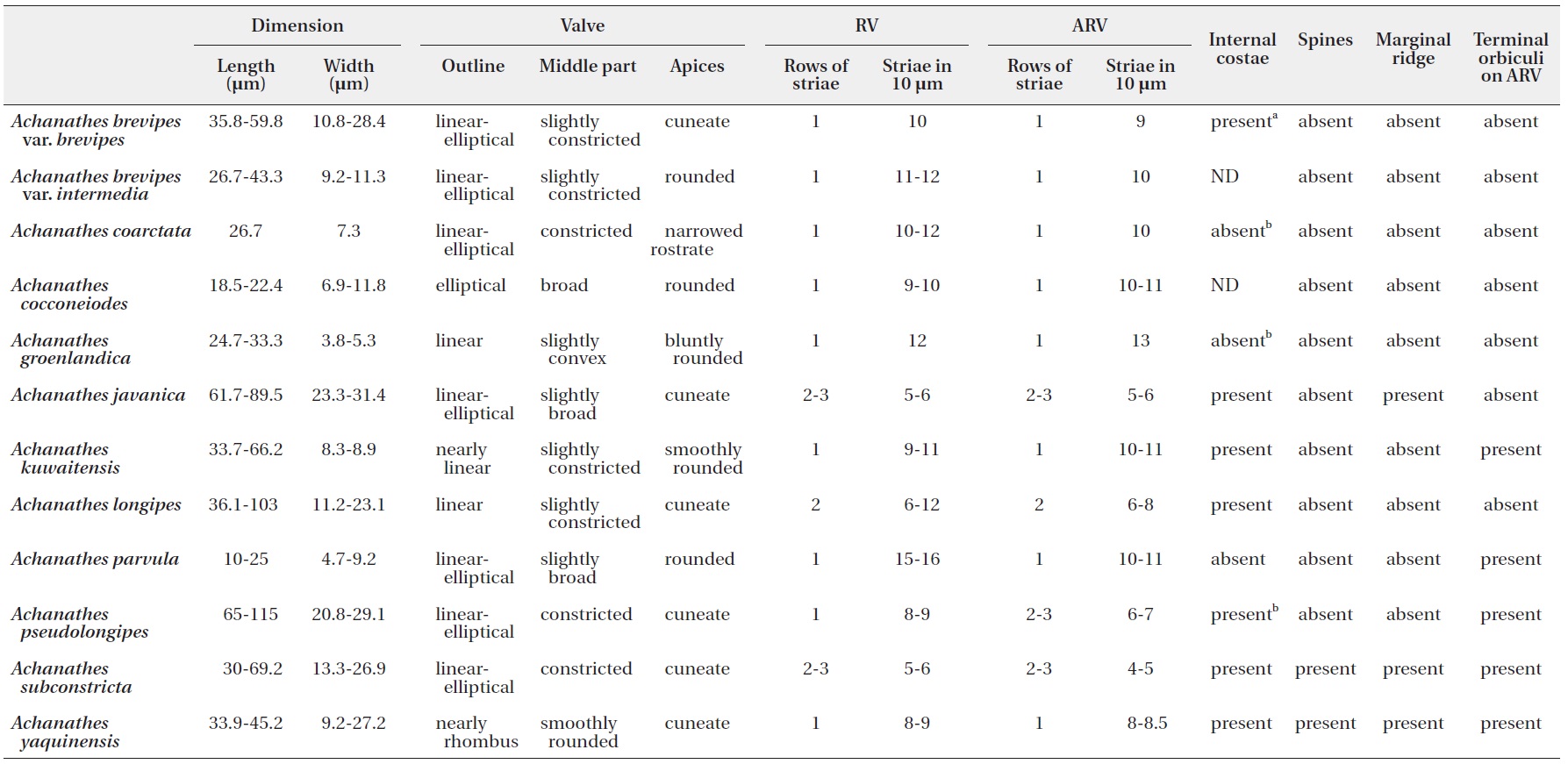
Morphological characteristics of Acnanthes species
>
Key to genus Achnanthes species
1a. One row of striae on the raphid valve (RV) and araphid valve (ARV) ...........................................................................2
1b. Multi-rows of striae on the RV and ARV .......................9
2a. Spines and marginal ridge on the valves .............................................................................................
2b. No spines and marginal ridge on the valves ................3
3a. Cuneate valve apices ...............
3b. No cuneate valve apices .................................................4
4a. Terminal obiculi on the ARV ..........................................5
4b. No terminal obiculi on the ARV ....................................6
5a. Internal costae inner valve .....................
5b. No internal costae inner valve .....................
6a. Narrowed rostrate valve apices ...................
6b. Rounded rostrate valve apices ......................................7
7a. Broad middle part of the valve ..............
7b. Not broad middle part of the valve ...............................8
8a. Striae in 10 μm on the RV more than the ARV ..................................................................
8b. Striae in 10 μm on the RV less than the ARV ........................................................................................
9a. Terminal obiculi on the ARV ....................................... 10
9b. No terminal obiculi on the ARV....................................11
10a. Rows of striae on the RV differ the ARV..........................................................................................
10b. Rows of striae on the RV same the ARV ............................................................................................
11a. Marginal ridge of the valve .........................
11b. No marginal ridge of the valve ..................
Achnanthes brevipes var. brevipes Agardh (
Synonym:
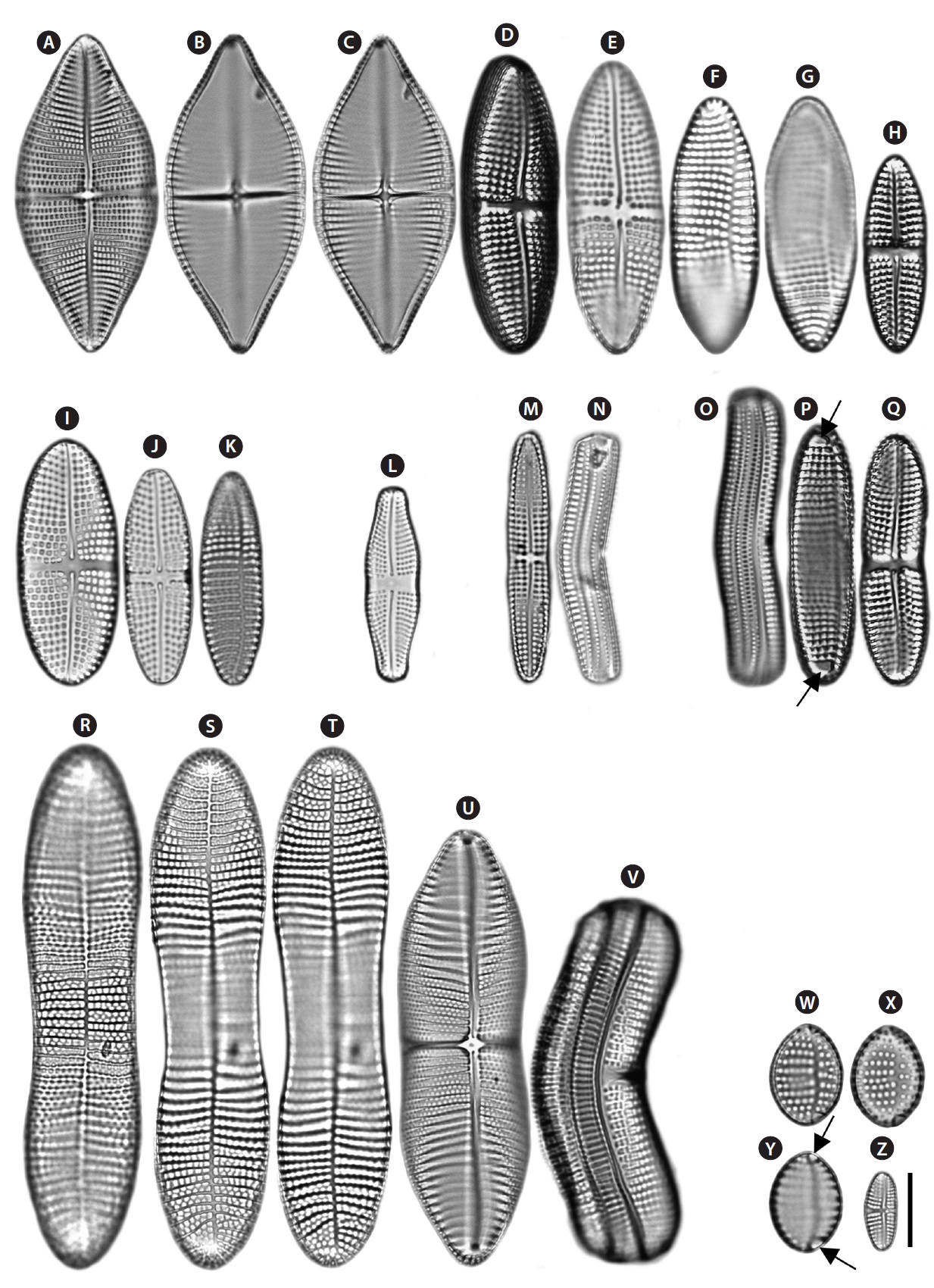
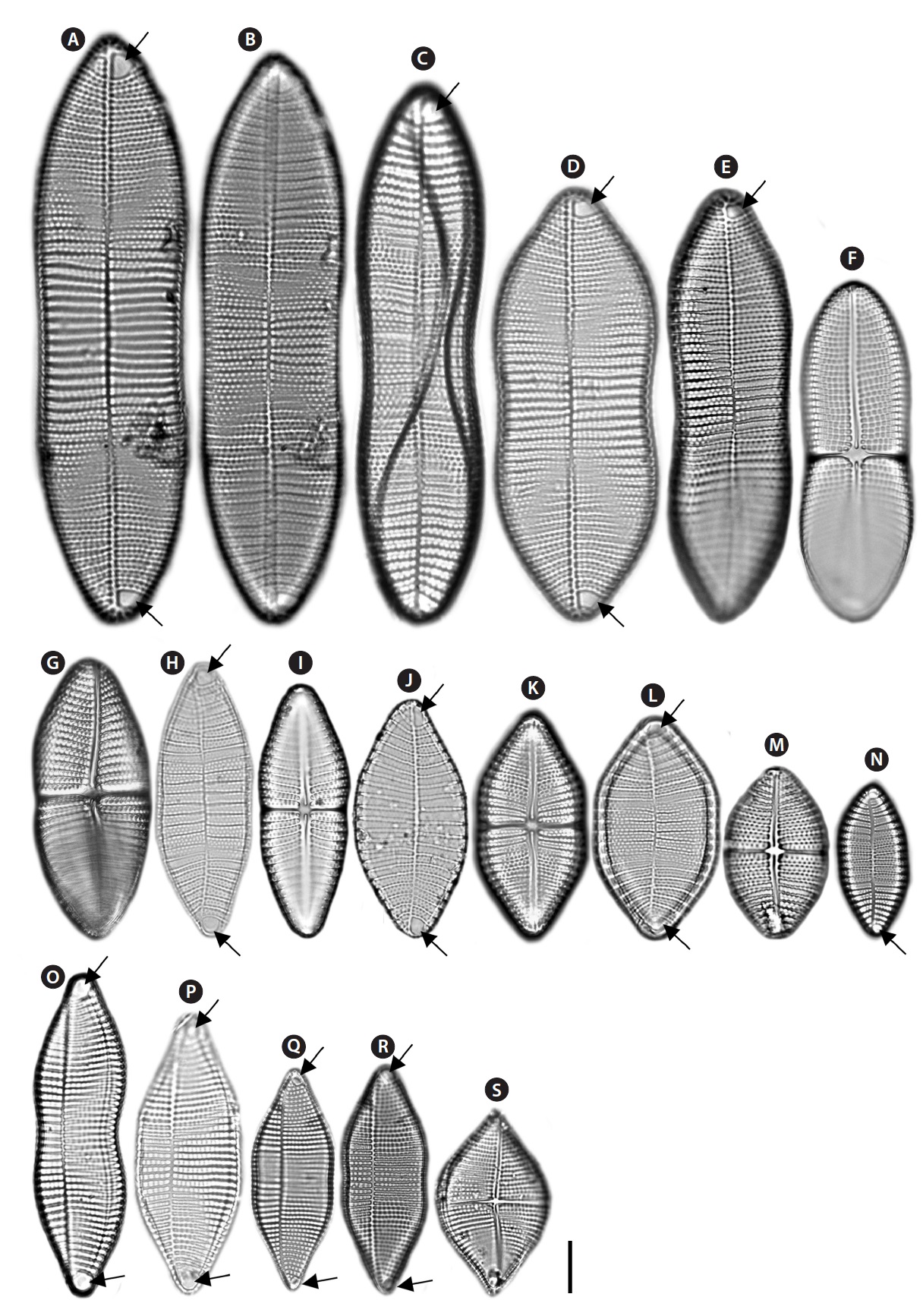
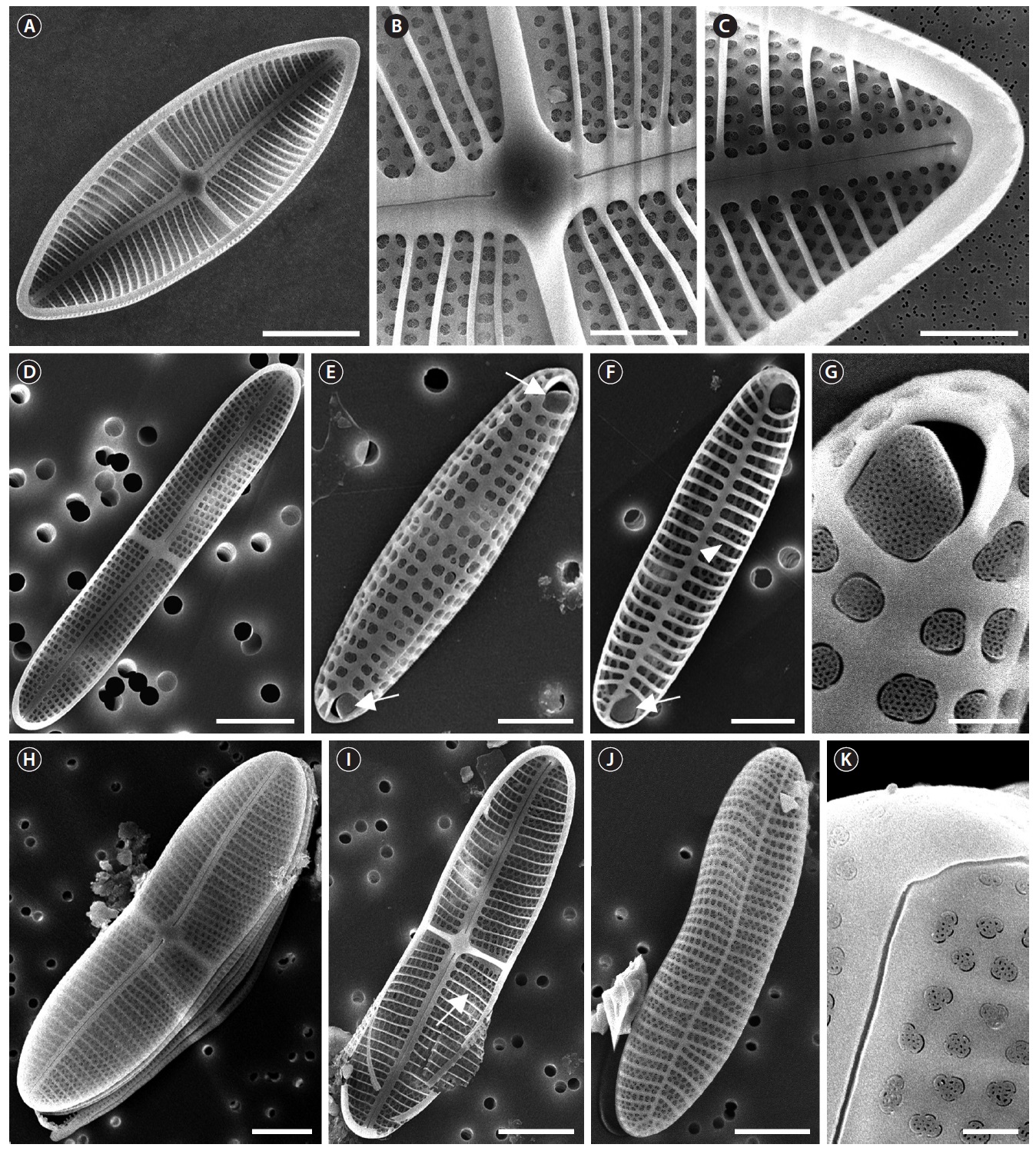
Description: Valves linear-elliptical, slightly constricted in the middle, cuneate at the apices. In girdle view, valve slightly curved geniculately with a convex ARV and a concave RV. 35.8-59.8 µm long, 10.8-28.4 µm wide. Striae uniseriate on both valves (ARV and RV) with nine striae in 10 µm on the ARV, and 10 striae in 10 µm on the RV. Internal valves with thickened transverse costae. Transapical striae slightly radial. Cells without spines and marginal ridge. Both valves without terminal obiculi.
Distribution:
References: Agardh 1824, p 1; Gaillon 1833, p 10; Heiberg 1863, p 118; Cleve 1895, p 193; Hendey 1951, p 41, pl. 16, figs. 9, 10.
Remarks: This species possesses the thickened transverse costae on the internal valves (Alfinito 1983), although it was not examined in this study.
Achnanthes brevipesvar. intermedia (Kutzing) Cleve (
Basionym:
Synonym:
Description: Valves linear-elliptical, sometimes slightly constricted in the middle, rounded at the apices. In girdle view, the valve is slightly curved geniculately, with a convex ARV and a concave RV. 26.7-43.3 µm long, 9.2-11.3 µm wide. Striae uniseriate on both valves (ARV and RV) with 10 striae in 10 µm on the ARV and 11-12 striae in 10 µm on the RV. Transapical striae slightly radial. Cells without spines and marginal ridge. Both valves without terminal obiculi.
Distribution:
References: Cleve 1895, p 193; Kützing 1833, p 576, figs. 55, 56; VanLandingham 1967, p 70; McIntire and Reimer 1974, p 171, pl. II, fig. 8a-b, pl. III, fig. 2a-b.
Remarks: Thickened transverse costae of
Achnanthes coarctata (Brebisson ex Smith) Cleve & Grunow (
Basionym:
Synonym:
Description: Valves linear-elliptical, constricted in the middle, with narrowed rostrate at the apices. In girdle view, the valve is slightly curved geniculately, with a convex ARV and a concave RV. 26.7 µm long, 7.3 µm wide. Striae uniseriate on both valves (ARV and RV) with 10 costae in 10 µm on the ARV and 10-12 costae in 10 µm on the RV. Internal valves without thickened transverse costae. Transapical striae slightly radial. Cells without the spines and marginal ridge. Both valves without terminal obiculi.
Distribution: Hustedt (1931) found this species commonly in drainage ditches and on wet rocks from northern Europe. Cox (2006) examined
References: Smith 1855, p 8, pl. 1, fig. 10; Cleve and Grunow 1880, p 121.
Remarks: Cox (2006) supposed it may be present spines or ridges at the face-mantle junction of the ARV such as
Achnnathes cocconeioides Riznyk (
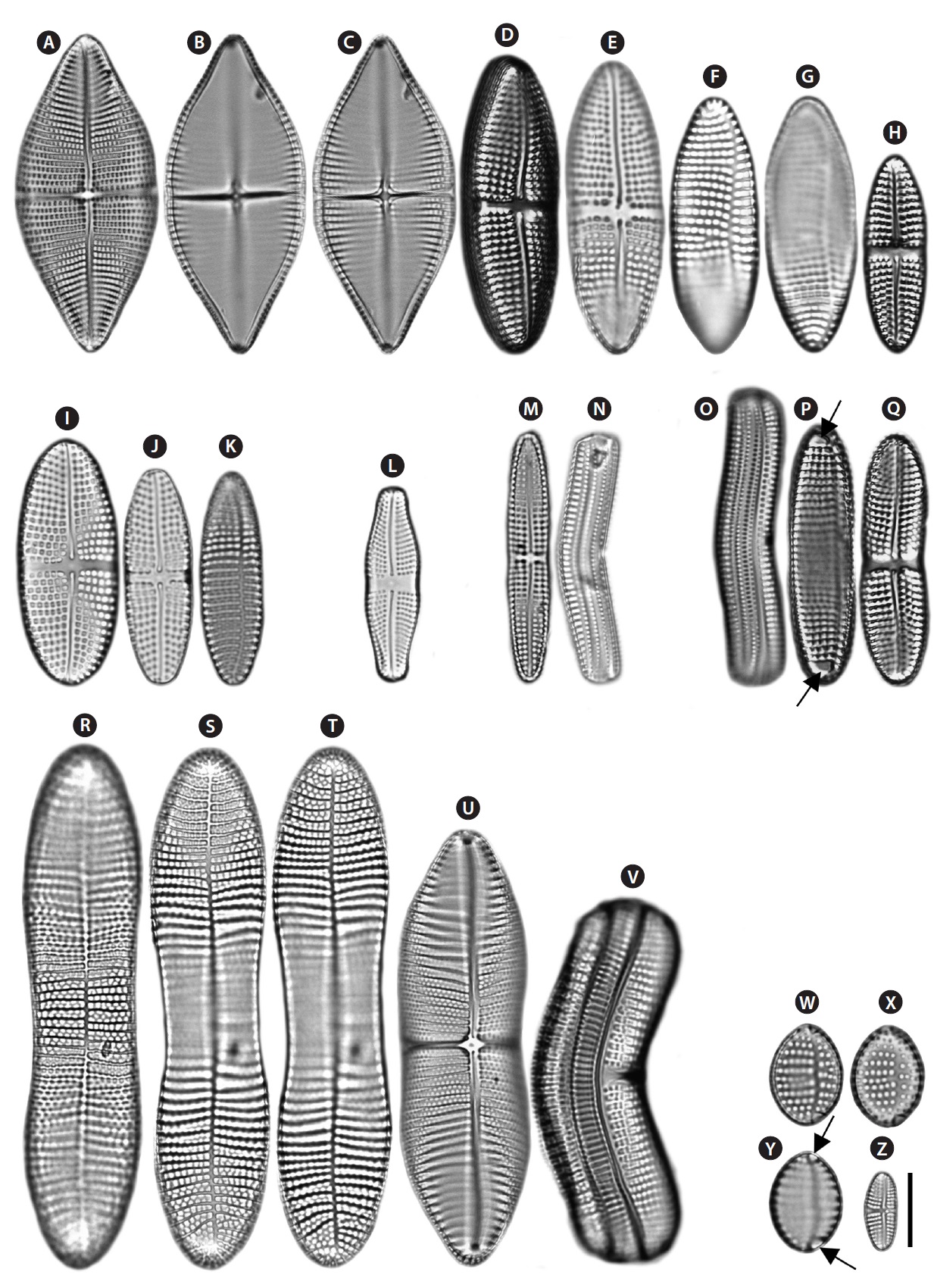
Description: Valves elliptical, broad in the middle, rounded at the apices. In girdle view, the valve is nearly flat. 18.5-22.4 µm long, 6.9-11.8 µm wide. Striae uniseriate on the both valves (ARV and RV) with 10-11 striae in 10 µm on the ARV and 9-10 striae in 10 µm on the RV. Transapical striae slightly radial. Cells without spines and marginal ridge. Both valves without terminal obiculi.
Distribution: Mclntire and Reimer (1974) recorded
References: Riznyk 1973, p 114, 115, pl. I, figs. 8, 9, pl. 18, figs. 1, 2; McIntire and Reimer 1974, p 172, pl. II, fig. 7a-b, pl. IV, fig. 2a-b.
Remarks: The shape of the central area and the lack of terminal orbiculi distinguish this species from
Achnanthes groenlandica Cleve & Grunow (
Basionym:
Synonym:
Description: Valves linear, slightly convex in the middle, bluntly rounded at the apices. In girdle view, the valve is slightly curved geniculately, with a convex ARV and a concave RV. 24.7-33.3 µm long, 3.8-5.3 µm wide. Striae uniseriate on both valves (ARV and RV) with 13 striae in 10 µm on the ARV and 12 striae in 10 µm on the RV. Internal valves without thickened transverse costae. Transapical striae slightly radial. Cells without spines and marginal ridge. Both valves without terminal obiculi.
Distribution: Cox (2006) examined
References: Cleve and Grunow 1880, p 20; Peragallo 1921, p 10; Cleve 1873, p 25, pl. 4, fig. 23.
Remarks: Cox (2006) supposed the striae pores of this taxon on the ARV may be markedly larger than on the RV. This was not examined in the present study. This species was newly recorded in Korean coastal waters. This species does not possess thickened transverse costae on the internal valves (Cox 2006), although it was not examined in this study.
Achnanthes javanica Cleve & Grunow (
Description: Valves linear-elliptical, slightly broad in the middle, cuneate at the apices. In girdle view, the valve is slightly curved geniculately, with a convex ARV and a concave RV. 61.7-89.5 µm long, 23.3-31.4 µm wide. Striae bi- or triseriate on the both valves (ARV and RV) with 5-6 striae in 10 µm on the ARV and 5-6 striae in 10 µm on the RV. Internal valves with thickened transverse costae. Transapical striae slightly radial. Cells with the marginal ridge but no spines. Both valves without terminal obiculi.
Distribution: Toyoda et al. (2003) examined
(22 Jan 2009), ES-11 (22 Jan 2009).
References: Cleve and Grunow 1880, p 18; Toyoda et al. 2003, p 367, figs. 5-10, 21-28.
Remarks:
Achnanthes kuwaitensis Hendey (
Synonym:
Description: Valves nearly linear, slightly constricted in the middle, smoothly rounded at the apices. In girdle view, the valve is slightly curved geniculately, with a convex ARV and a concave RV. 33.7-66.2 µm long, 8.3-8.9 µm wide. Striae uniseriate on both valves (ARV and RV) with 10-11 striae in 10 µm on the ARV and 9-11 striae in 10 µm on the RV. Internal valves with thickened transverse costae. Transapical striae nearly linear. Cells without marginal ridge and spines. Terminal orbiculi with both ends of the ARV but not on the RV.
Distribution: Mclntire and Reimer (1974) reported this species from the Yaquina Estuary, Oregon (U.S.A.) in Aug 1968, Oct 1968, and Jul 1969. Presently,
References: Hendey 1958, p 55, pl. 6, figs. 8-10; VanLandingham 1967, p 73; McIntire and Reimer 1974, p 173, pl. II, fig. 6a-c, pl. III, fig. 4a-b.
Remarks: This taxon can be easily distinguished from other linear shaped
Achnanthes longipes Agardh (
Synonym:
Description: Valves linear, slightly constricted in the middle, wedged at the apices. In girdle view, the valve is slightly curved geniculately, with a convex ARV and a concave RV. 36.1-103 µm long, 11.2-23.1 µm wide. Striae biseriate on both valves (ARV and RV) with 6-8 striae in 10 µm on the ARV and 6-12 striae in 10 µm on the RV. Internal valves with thickened transverse costae. Transapical striae nearly linear or slightly radial. Cells without marginal ridge and spines. Both valves without terminal obiculi.
Distribution: Cox (2006) studied
References: Agardh 1824, p 1; Gaillon 1833, p 10.
Remarks:
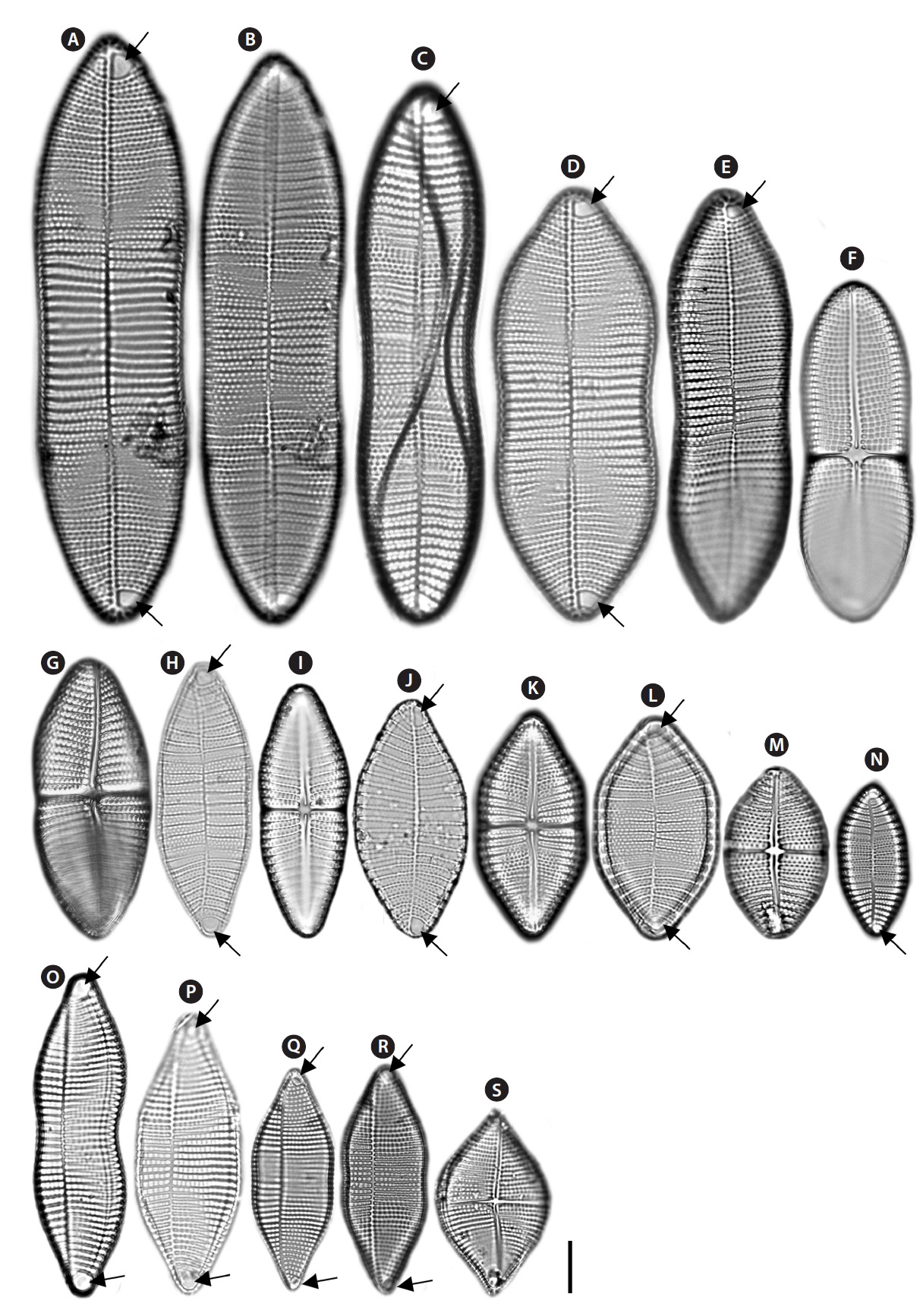
Achnanthes parvula Kutzing (
Synonym:
Description: Valves linear-elliptical, slightly broad in the middle, rounded at the apices. In girdle view, the valve is slightly curved geniculately, with a convex ARV and a concave RV. 10-25 µm long, 4.7-9.2 µm wide. Striae uniseriate on both valves (ARV and RV) with 10-11 striae in 10 µm on the ARV and 15-16 striae in 10 µm on the RV. Internal valves without thickened transverse costae. Transapical striae nearly linear or slightly radial. Cells without marginal ridge and spines. Terminal orbiculi with both ends of the ARV but not on the RV.
Distribution: Mclntire and Reimer (1974) recorded
References: Kützing 1844, p 76, pl. 21, fig. 5; Cleve 1895, p 193; VanLandingham 1967, p 71; Mereschkowsky 1901, p 31.
Remarks: This taxon ostensibly resembles
Achnanthes pseudolongipes Toyoda & Nagumo in Toyoda et al. (
Description: Valves linear-elliptical, constricted in the middle, cuneate at the apices. In girdle view, the valve is slightly curved geniculately, with a convex ARV and a concave RV. 65-115 µm long, 20.8.-29.1 µm wide. Striae bi- to triseriate with 6-7 striae in 10 µm on the ARV, uniseriate with 4-5 striae in 10 µm on the RV. Internal valves with
thickened transverse costae. Transapical striae slightly radial. Cells without the marginal ridge and spines. Terminal orbiculi at both ends of the ARV but not on the RV.
Distribution: Toyoda et al. (2010) found
References: Toyoda et al. 2010, p 186, figs. 1-28.
Remarks:
Achnanthes subconstricta (Meister) Toyoda (
Basionym:
Description: Valves linear-elliptical, constricted in the middle, with cuneate at the apices. In girdle view, the valve is slightly curved geniculately, with a convex ARV and a concave RV. 30-69.2 µm long, 13.3-26.9 µm wide. Striae bi- to tetraseriate on the both valves (ARV and RV) with 4-5 striae in 10 µm on the ARV and 5-6 striae in 10 µm on the RV. Internal valves with thickened transverse costae. Transapical striae slightly radial. Cells with the marginal ridge and spines on the both terminal ends and marginal ridge of the valve. Terminal orbiculi with both ends of the ARV but not on the RV.
Distribution: Mclntire and Reimer (1974) recorded the species from the Yaquina Estuary, Oregon, in Aug 1968, and Jul 1969. Toyoda et al. (2003) examined
References: Toyoda et al. 2003, p 369; Meister 1932, p 40, pl. 14, fig. 113.
Remarks: This taxon is very similar in shape to
Achnanthes yaquinensis McIntire & Reimer (
Description: Valves nearly rhombus, smoothly rounded in the middle, cuneate at the apices. In girdle view, the valve is slightly curved geniculately, with a convex ARV and a concave RV. 33.9-45.2 µm long, 9.2-27.2 µm wide. Striae uniseriate on the both valves (ARV and RV) with 8-8.5 striae in 10 µm on the ARV and 8-9 striae in 10 µm on the RV. Internal valves with thickened transverse costae. Transapical striae nearly linear or slightly radial. Cells with the marginal ridge and spines on the both terminal ends of the valve. Terminal orbiculi with both end of the ARV but not on the RV.
Distribution: Mclntire and Reimer (1974) found
References: McIntire and Reimer 1974, p 174, pl. II, fig. 1a-b, pl. III, fig. 1a-b.
Remarks: This taxon is very similar in shape to
Twelve
Most of
We present the distribution of
Consequently, valid nomenclatural-named
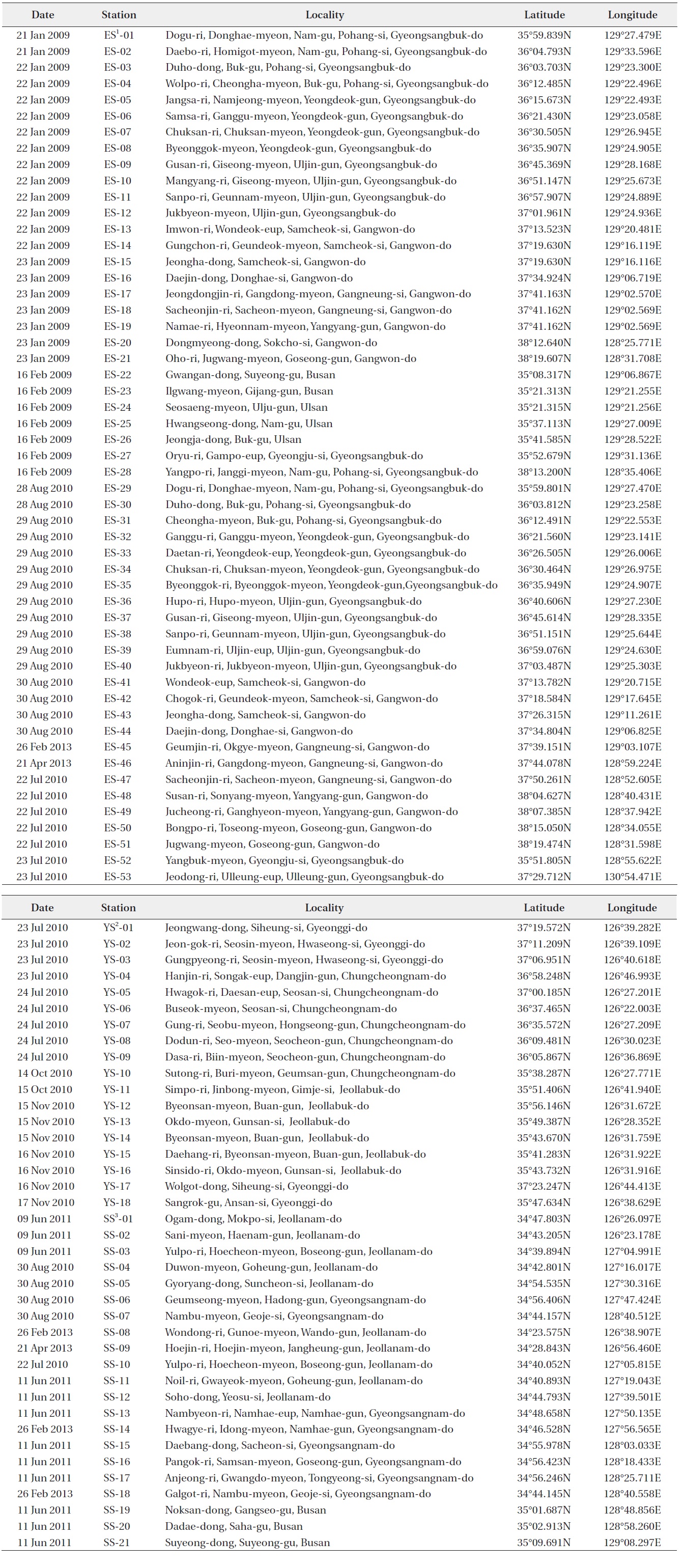
[Table 3.] Sampling sites in coastal waters of Korea
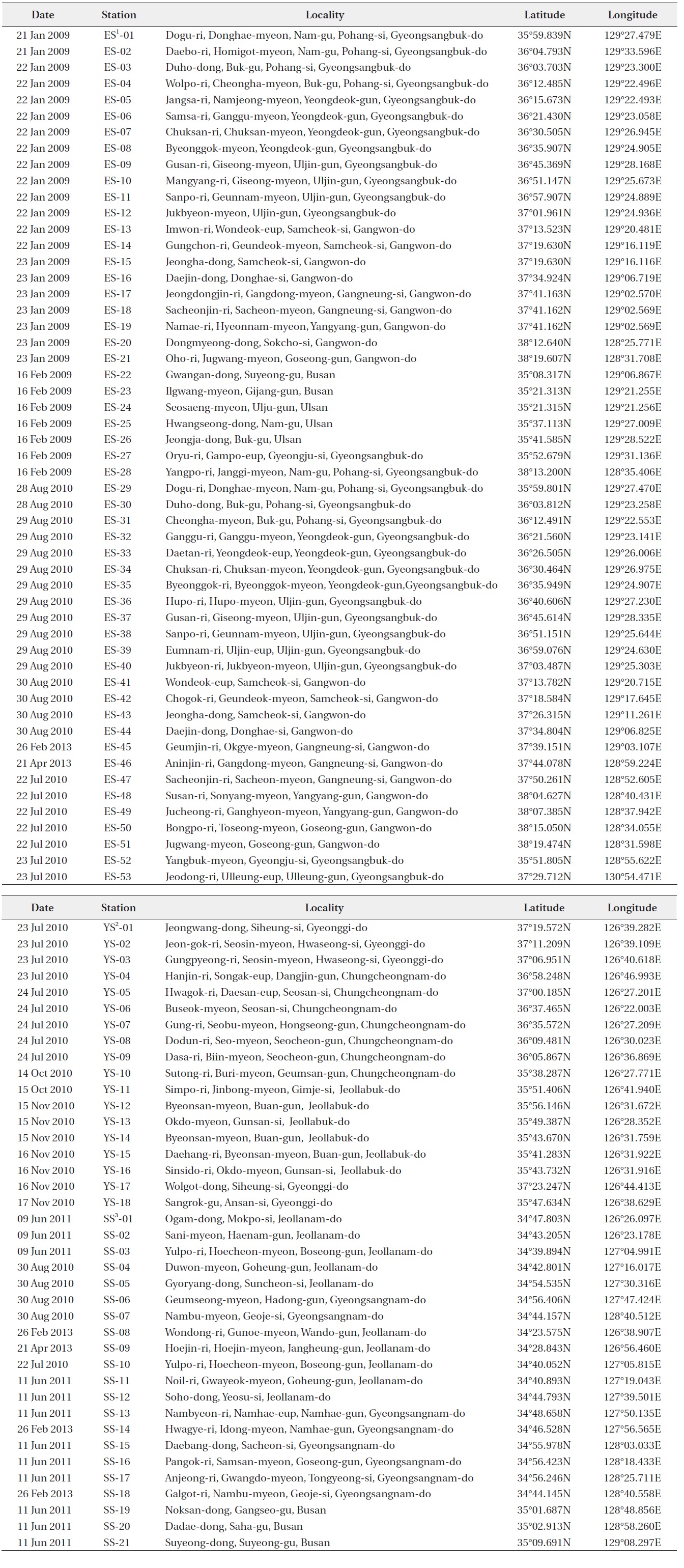
Sampling sites in coastal waters of Korea