A new copepod species, Cletocamptus koreanus, is described from estuaries and salt marshes on south and southwest coasts in Korea. Cletocamptus koreanus n. sp. evidently belongs to C. deitersi species group in sharing 6- segmented antennule, 3 setae on antennary exopod, leg 1 with endopod shorter than exopod, and 2+1 inner setae on the third exopodal segments of legs 3-4. However, C. koreanus is distinguished from its allied congeneric species by different setal armatures of mandibular palp, antennary exopod, endopod of female leg 2 and the third exopodal segments of legs 3-4, and relative length and shape of apophysis of male leg 3 enp 2. Description and taxonomic accounts of the new species are presented herein, with detailed illustrations and scanning electron microscope photomicrographs. Discussion on the affinities with other species of C. deitersi group are provided, along with brief comments on the morphological discrepancies between the new species from Korea and other Asian species from China and India.
A series of faunistic papers relating to harpacticoid copepods from various brackish waters in Korea have been published:
As one of the serial studies on the brackish copepod fauna, the author re-examined a brackish species collected from salt marshes on south and southwest coasts of Korea, which had been remained as an undetermined species, ‘
As the above-mentioned
Materials examined in the present study were collected at a brackish swamp near waterways of a salt farm located at Imjado Island, off the southwestern coast of Korea on Aug 21, 2008. Collections were made with a dipnet of 64 μm mesh. Copepods were fixed and stored in 4% buffered formalin.
The process of general methods, including dissection, preparation, observation and measurements, follows my previous paper (Lee and Chang, 2012).
Materials for scanning electron microscopy were prefixed overnight at 4℃ in 2.5% glutaraldehyde, followed by rinsing with 0.1 M phosphate buffer (pH 7.2-7.4) three times, each for 10 min. Specimens were postfixed for 2 h in 2% cold osmium tetroxide in 0.1 M phosphate buffer, and were then left in phosphate buffer overnight. After dehydration through a graded series of ethanol (50-100% at 10% interval) for 30 min each, the material was critical point dried, and coated with gold-palladium in a high evaporator, and was then examined with a scanning electron microscope (Hitachi S-4800; Hitachi, Tokyo, Japan) operated at 15 kV.
Type specimens are deposited in the National Institute of Biological Resources (NIBR), Incheon, Korea, and the specimen room of the Department of Biological Science, Daegu University (DB), Korea.
Abbreviations used in the text and figure legend follow the conventional ones frequently used in the taxonomy of copepods: A1, antennule; A2, antenna; enp 1-3 or exp 1-3, the first to third endopodal or exopodal segment of each leg.
Order Harpacticoida Sars, 1903
Family Canthocamptidae Brady, 1880
Genus Cletocamptus Shmankevitsch, 1875
1*Cletocamptus koreanus n. sp. (Figs. 1-6)
Type. Holotype ♀ (NIBRIV0000271765), allotype ♂ (NI BRIV0000271766), undissected in 4% buffered formalin; brackish swamp beside waterway through salt farms, Imjado Island, off southwestern coast of Korea (35°03′24.33′′N, 126°06′08.40′′E), 21 Aug 2008 (
Additional material examined: Korea: 2♀♀ (DB20040), dissected in lactophenol, Busan, Jinudo islet (delta) (35° 04′08.40′′N, 128°52′14.49′′E), 8 Aug 1988 (
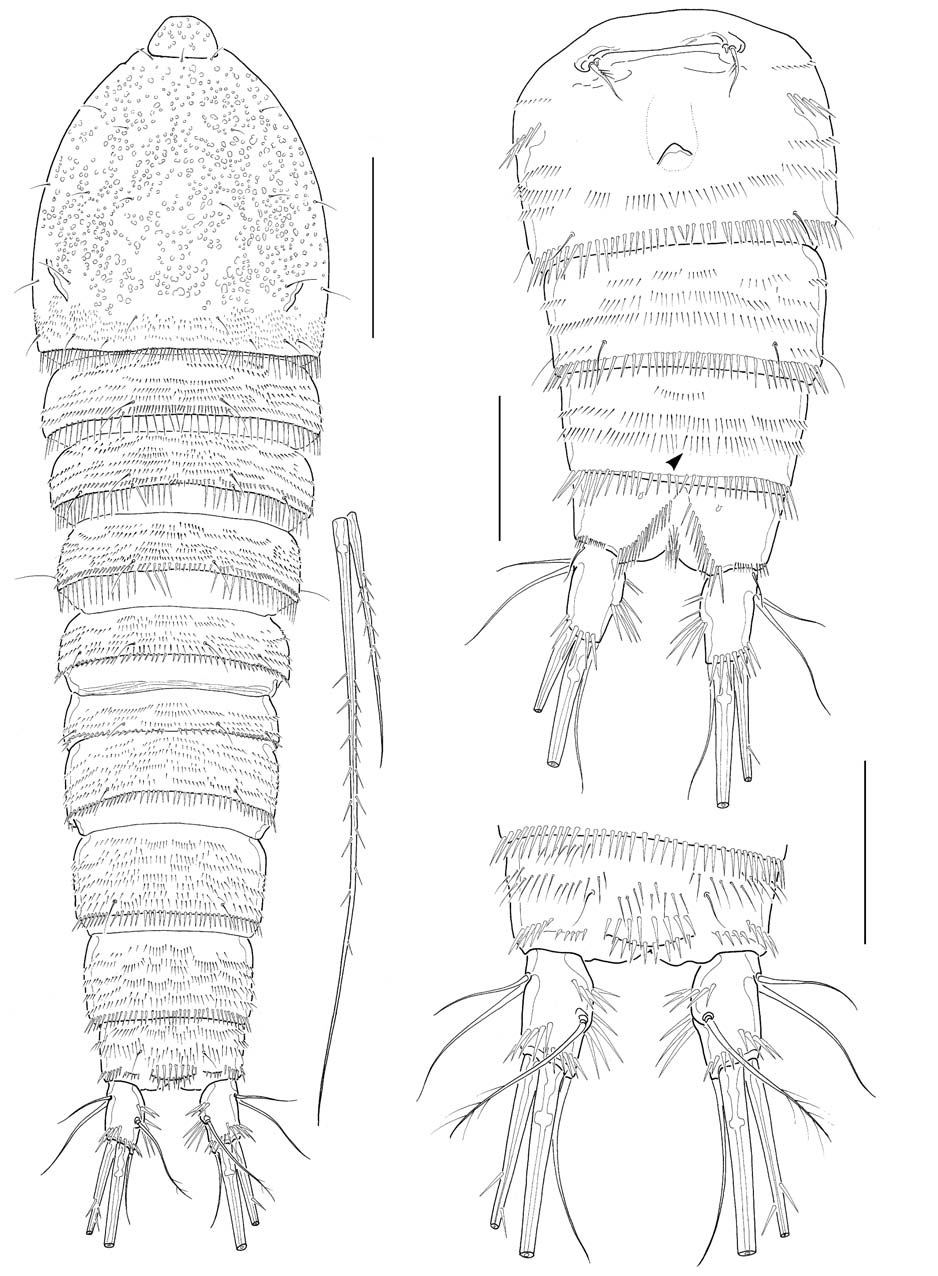
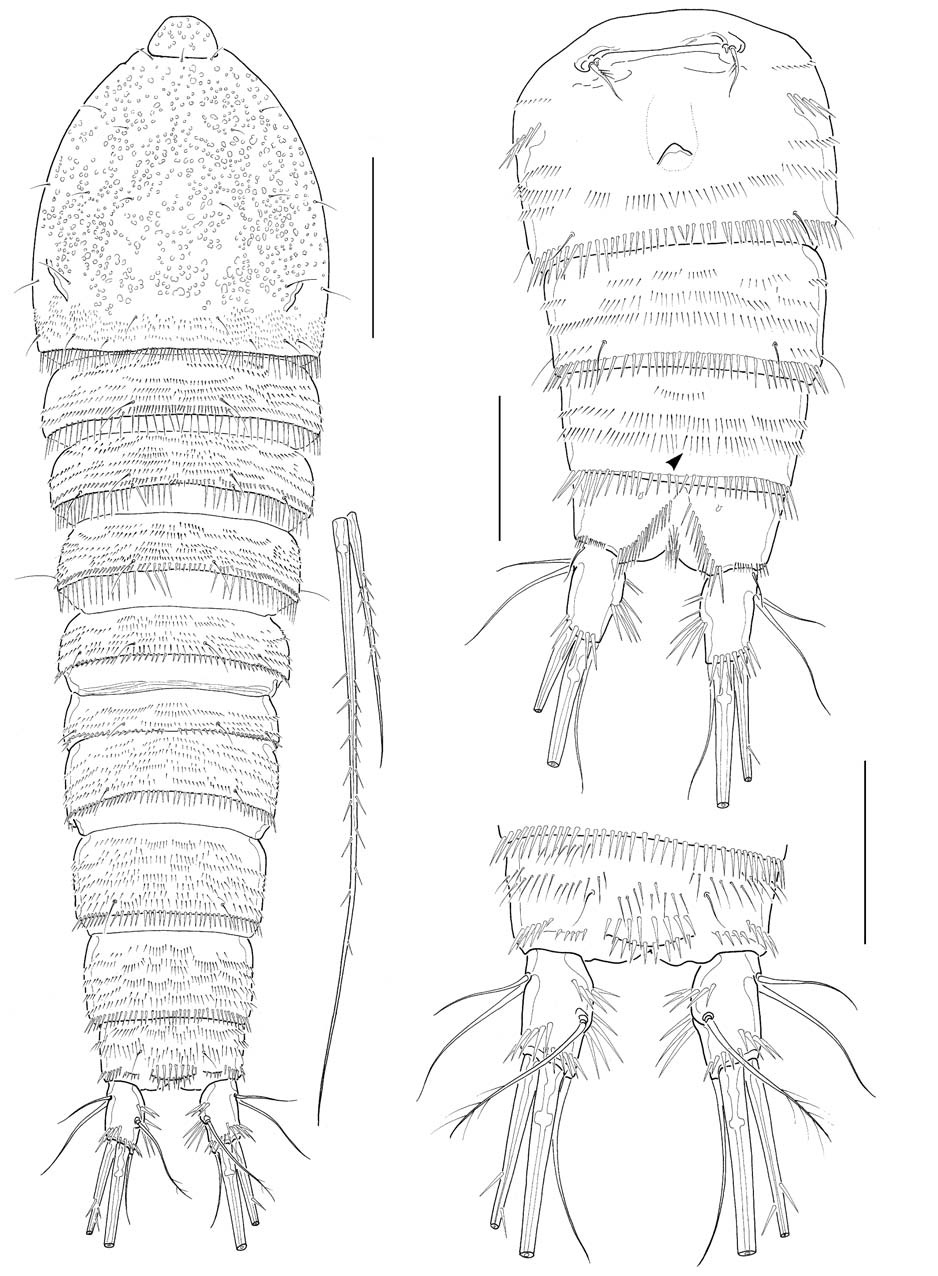
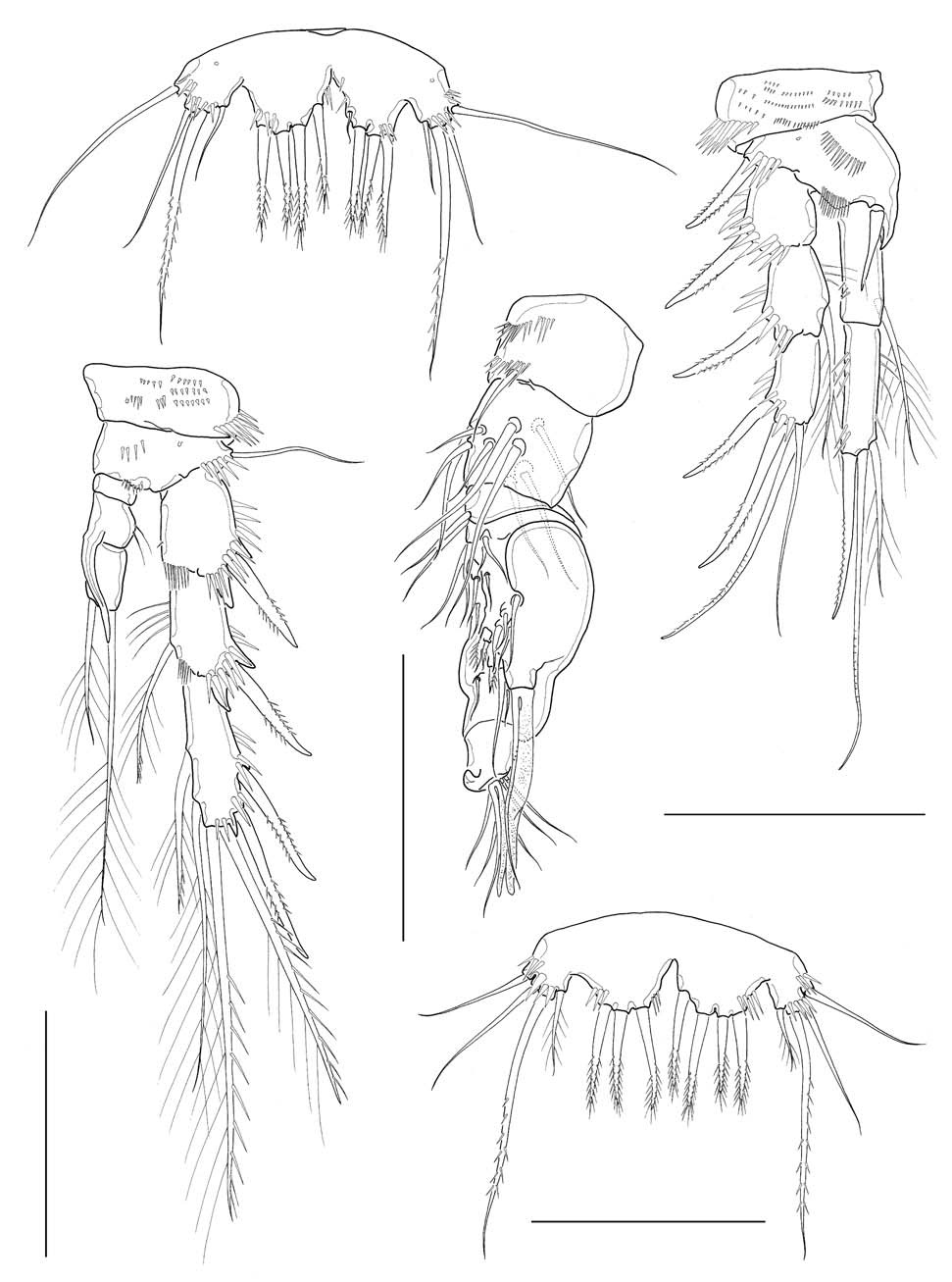
Caudal rami (Figs. 1C, 6A) truncate, about 1.7 times longer than wide, a little divergent posteriorly; dorsal keel not apparent; proximal part of medial margin a little swollen with 2 oblique spinule rows along medial face. Lateral caudal setae (caudal setae I and II) located proximally; outer caudal seta (caudal seta III) absent; outer terminal caudal seta (caudal seta IV) pinnate, a little shorter than half the inner terminal caudal seta (caudal seta V); inner caudal seta (caudal seta VI) naked and slender, slightly less than 2 times as long as caudal ramus; dorsal caudal seta (caudal seta VII) situated near middle of medial margin of caudal ramus.
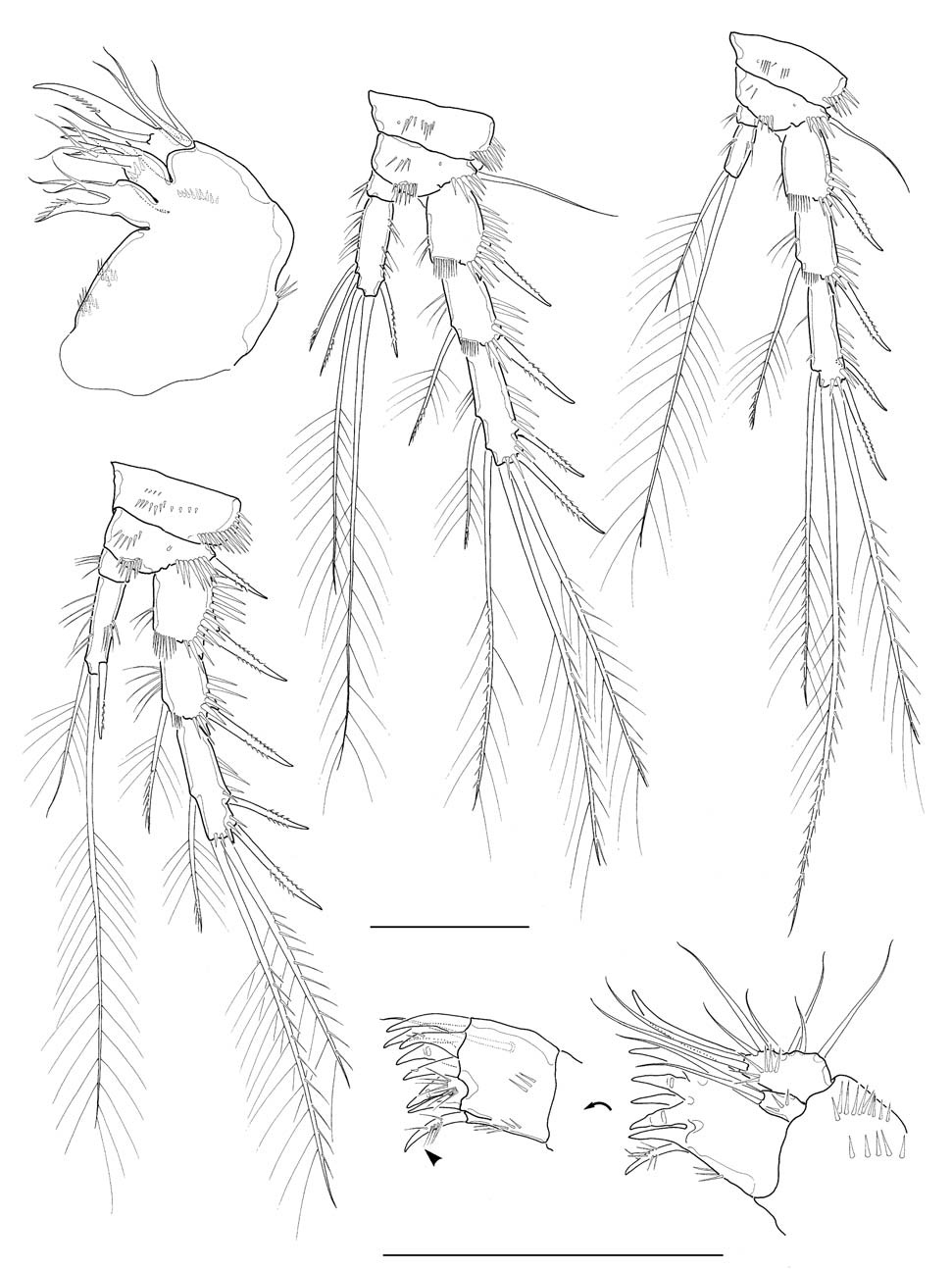
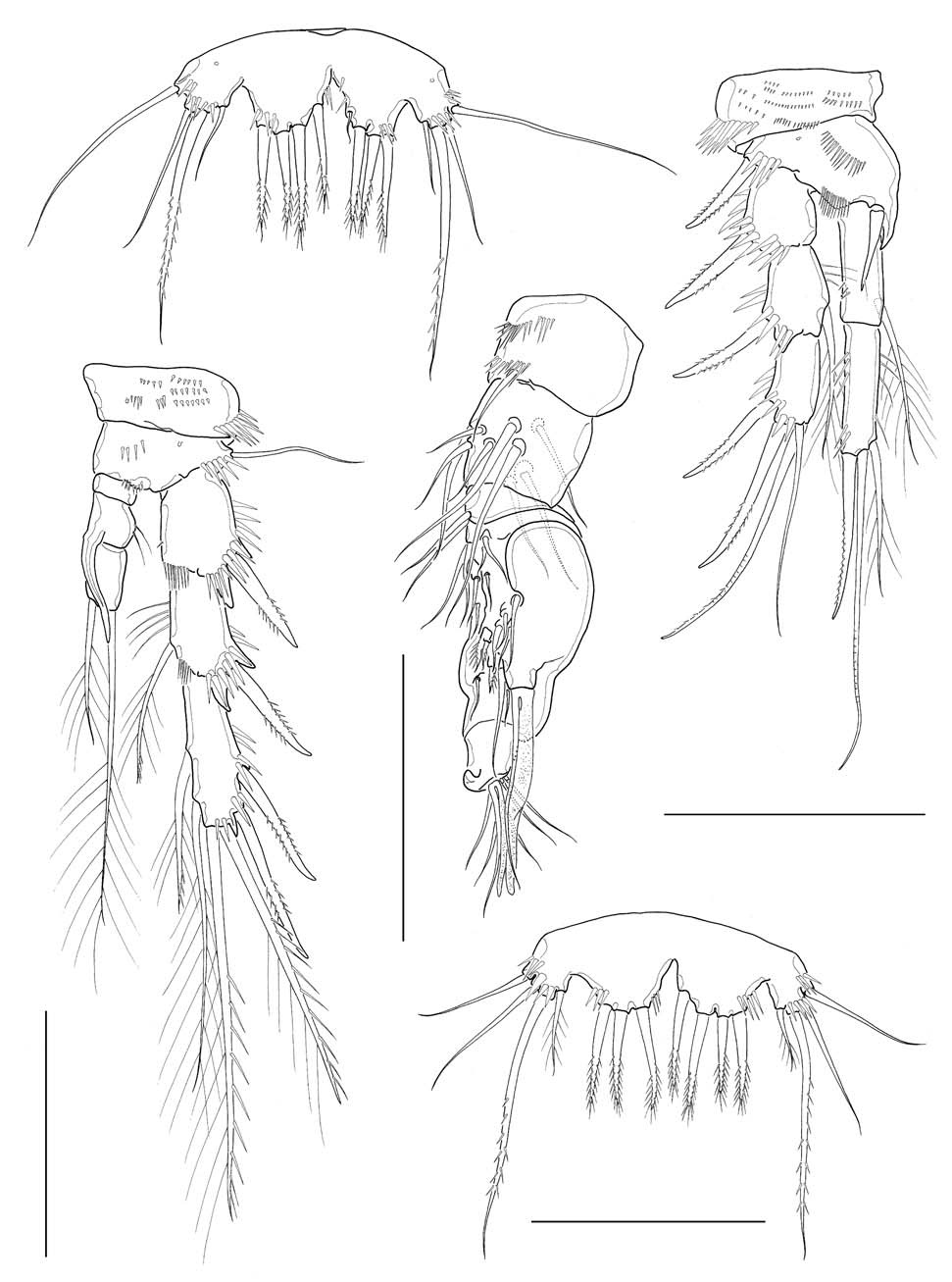
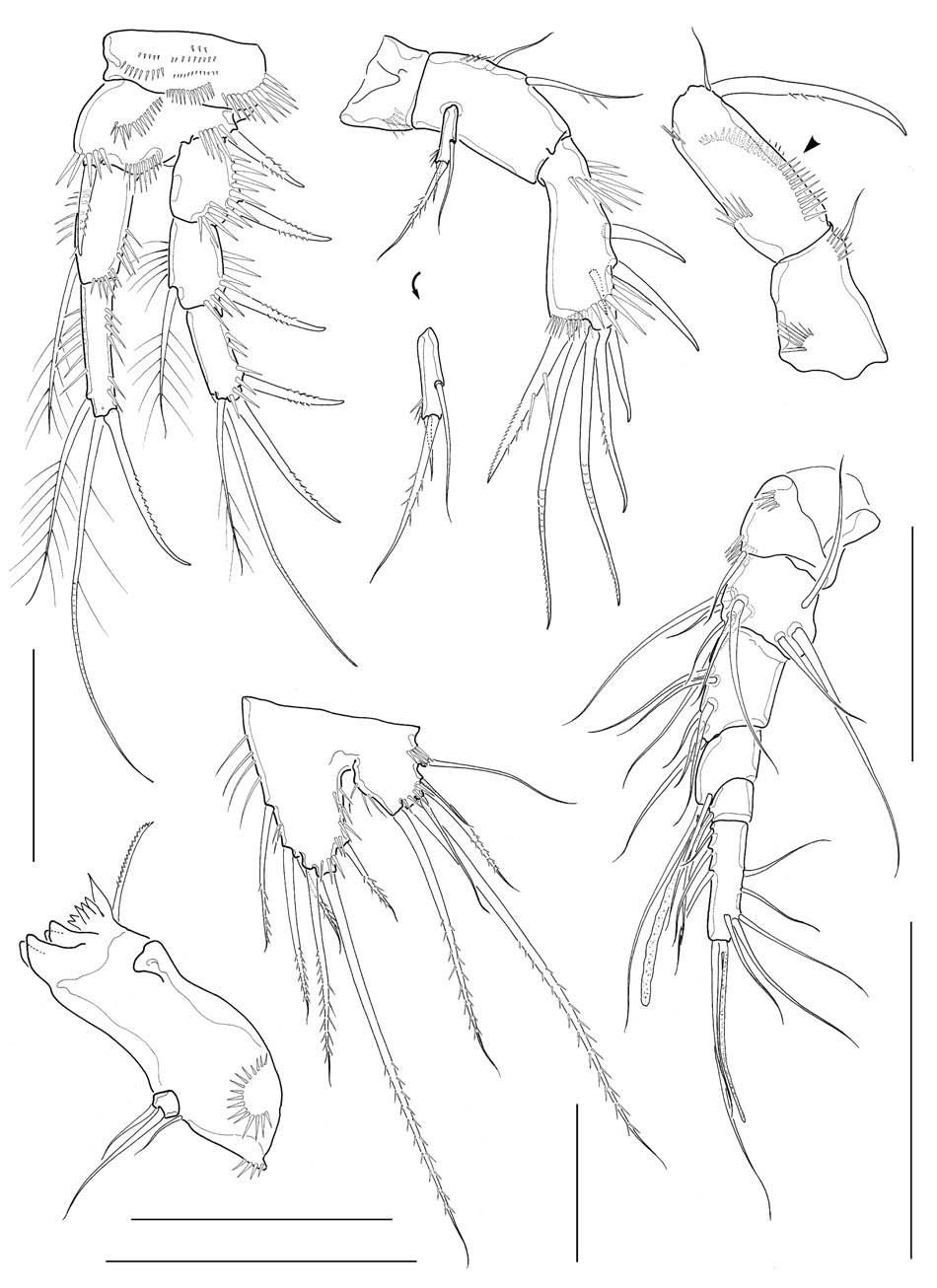
Antennule (Fig. 2A) 6-segmented, its tip not reaching middle of cephalothorax; segments 1-3 stout, segment 5 apparently small, segment 6 elongate; segment 4 with 1 aesthetasc, its tip much exceeding distal end of antennule. Armature formula: 1-[1], 2-[9], 3-[6], 4-[1+(1+aesthetasc)], 5-[1], 6- [9+(1+aesthetasc)] (all setae naked). Antenna (Fig. 2B), coxa small, unarmed; allobasis with 2 abexopodal setae; exopod 1-segmented, slender, cylindrical, with 3 setae (1 medial and 2 apical) in total; free endopodal segment armed with 2 lateral spines, 5 distal elements and 1 small seta on inner distal corner. Mandible (Figs. 2C, 6D), with well developed coxal gnathobase bearing 4-5 bicuspidate and multicuspidate teeth and 1 lobate element along distal margin and 1 lateral seta at dorsal corner; palp vestigial, represented by 1 small segment bearing 2 naked seta apically, flanked by 1 slender seta nearby. Maxillule (Figs. 3A, 6E) with praecoxal arthrite bearing 7 elements with 1 seta on frontal surface and 1 spiniform, pinnate seta posteriorly; coxal arthrite with cylindrical endite bearing 2 setae apically; exopod and endopod completely fused to basis, bearing total 9 setae apically and along anterior margin. Maxilla (Fig. 3B) armed with 2 syncoxal endites, each endite bearing 3 setal elements; allobasis forming 1 strong pectinate claw, flanked by 2 setae; endo-pod
represented by small protuberance bearing 2 naked setae. Maxilliped (Figs. 2D, 6F) subchelate; syncoxa a little protruded distomedially with 1 slender seta, flanked by spinule row at its base; basis about 3 times longer than wide, armed with longitudinal rows of spinules both anterior and posterior surfaces along medial margin; spinule row on posterior surface consisting of 10-12 spinules, distalmost spinule not reaching halfway of medial margin of basis; endopod represented by 1 strong and curved claw, bearing 1 minute seta proximally as accessory armature.
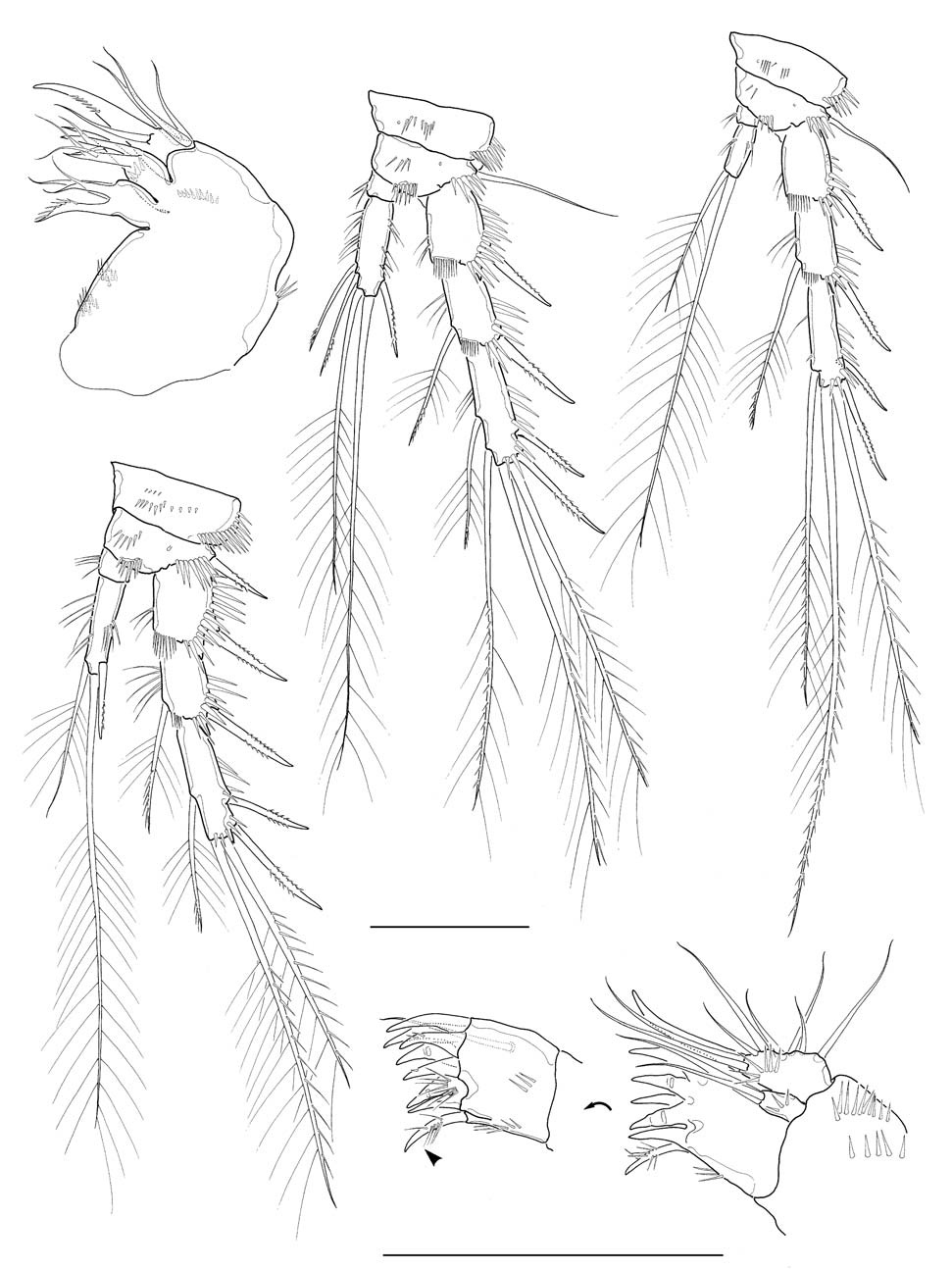
Legs 1-4 with 2-segmented endopods and 3-segmented exopods. Leg 1 (Fig. 2E), distomedial seta on basis not reaching posterior end of enp 1; enp 1 about 2.7 times as long as wide, slightly shorter than enp 2, not reaching distal end of exp 2, with 1 inner seta distally; enp 2 with 1 inner distal plumose seta, 1 apical geniculate seta and 1 long distal spine; exp 2 not elongate, with 1 inner seta; exp 3 armed with 2 outer spines and 2 apical setae. Legs 2-4 (Fig. 3C-E), enp 1 small, lacking inner seta. Leg 2 enp 2 elongate, about 3.6 times as long as broad, bearing 2 plumose setae and 1 spine with 1 setule row along medial margin and 1 oblique setule row near middle of lateral margin; apical seta about 2.8 times longer than inner distal seta. Leg 3 enp 2 elongate, about 3.2 times as long as broad, bearing 4 setae and 1 spine apically or subapically; 2 inner distal setae short, modified. Leg 4 enp 2 rather ellipsoidal, bearing 2 distal setae apically. Seta/spine armature of legs 1-4 as follows:
Leg 1 basis I-1 exp I-0; I-1; II,1,1 enp 0-1; I,1,1
Leg 2 basis I-0 exp I-0; I-1; II,2,1 enp 0-0; I,1,1
Leg 3 basis 1-0 exp I-0; I-1; II,2,2 enp 0-0; I,2,2
Leg 4 basis 1-0 exp I-0; I-1; II,2,1 enp 0-0; 0,2,0
Leg 5 (Fig. 2F), baseoendopod and exopod confluent basally, bilobed distally; baseoendopod protruding, much exceeding exopod, bearing 6 setae; exopodal lobe small, wider than long, bearing 5 setae excluding baseoendopodal lateral seta, with setules along swollen medial margin.
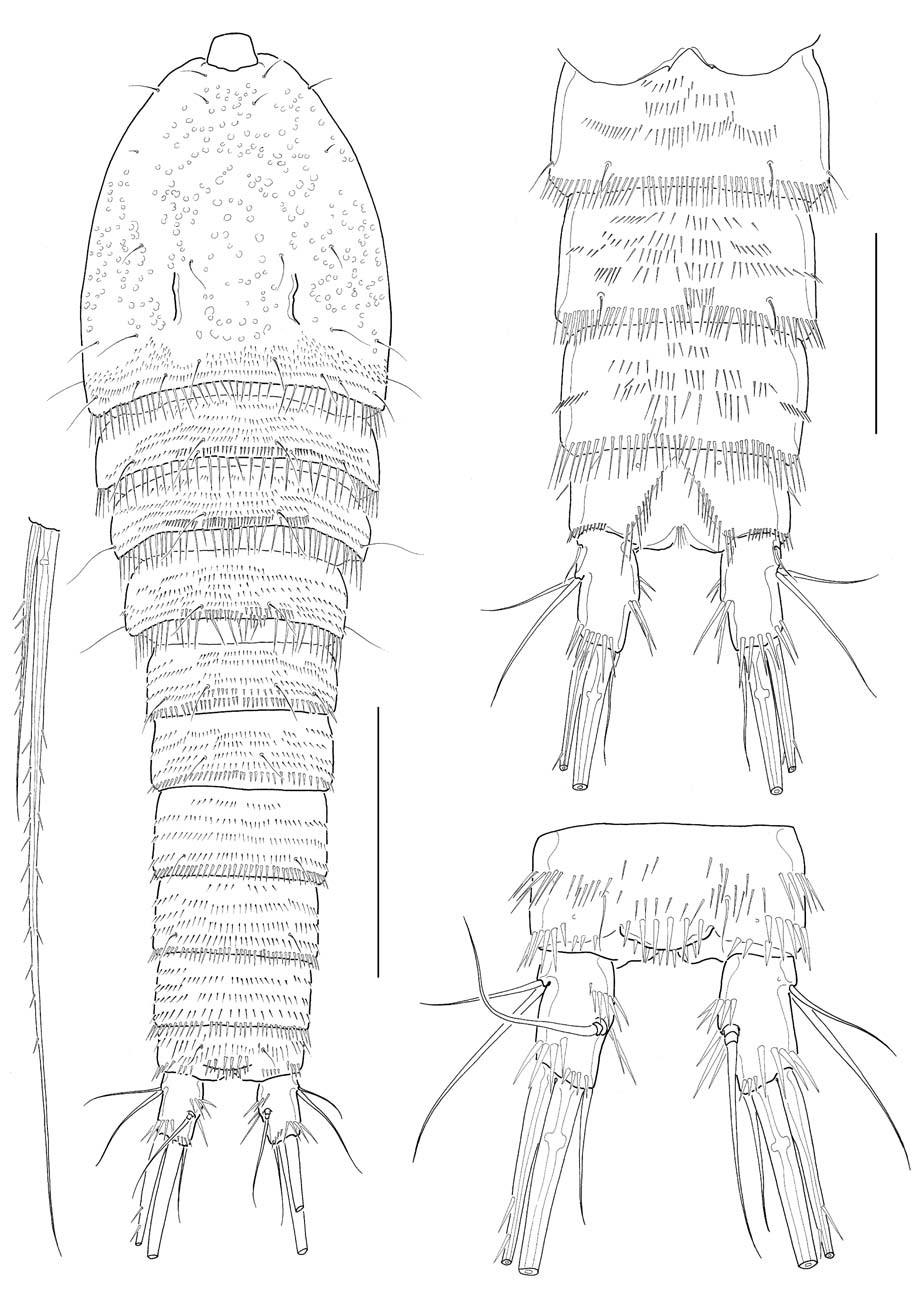
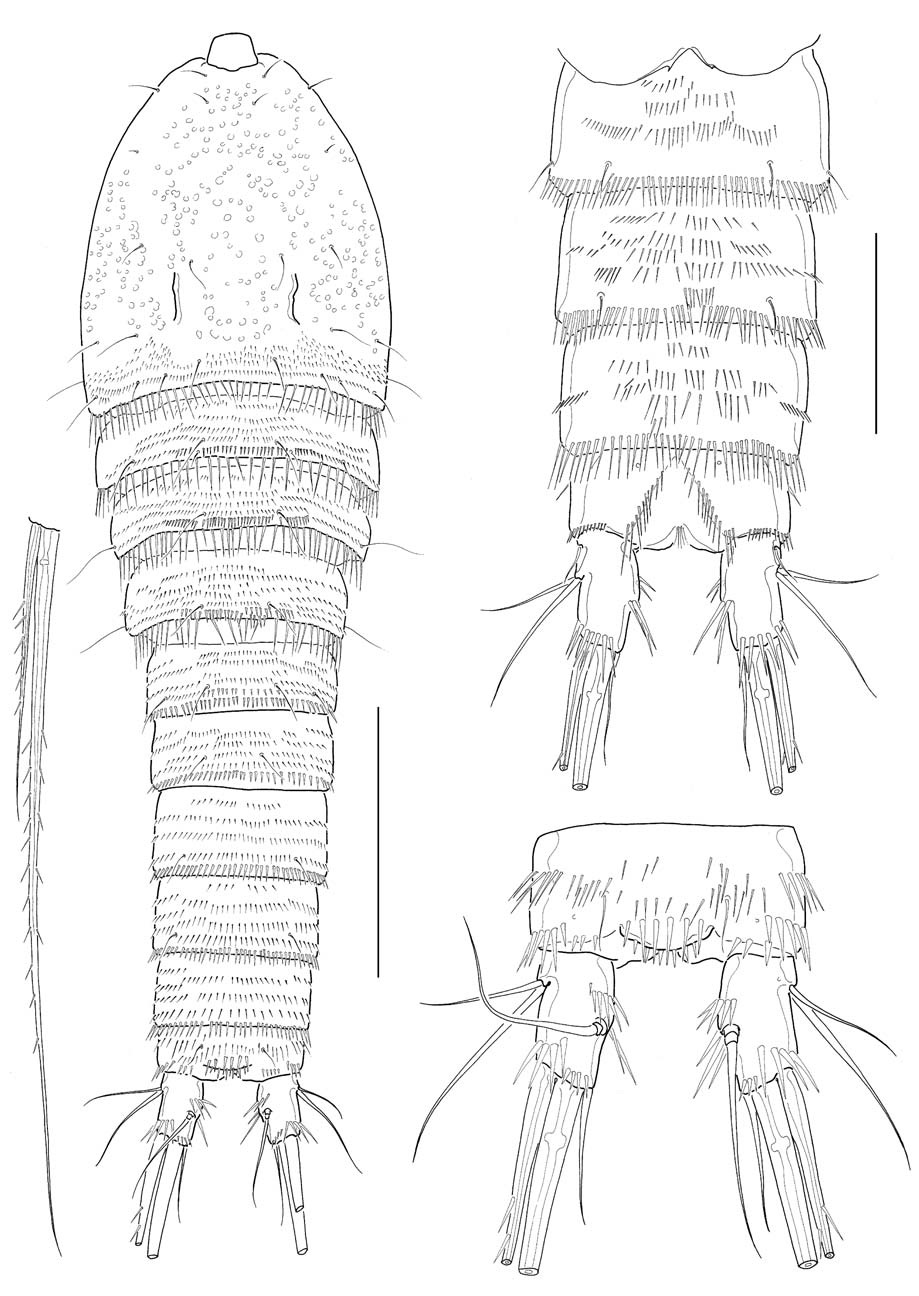
Male: Body (Fig. 4A) about 530 μm in length. General shape of body similar to that of female. Rostrum much narrower than in female. Posterior margins of third to sixth urosomites each densely lined with sharp spines; 2 spinule rows medially on ventral surface of third urosomite; 3-4 spinule rows on ventral surfaces of next 2 urosomites; setae on anal somite lappet short (Fig. 4B). Shape and armature of caudal rami nearly same as those of females, except for relatively a little longer rami, about 1.9 times as long as wide (Fig. 4C).
Antennule (Fig. 5A) subchirocerate, 6-segmented; segment 4 bulbous, armed with 3 pinnate setae and 1 aesthetasc. Leg 1 (Fig. 5B), distomedial corner of basis forming a sharp projection; inner distal seta on basis not modified, not reaching posterior end of enp 1; endopod slightly shorter than exopod; enp 1 slightly shorter than enp 2, not reaching distal end of exp 2. Leg 3 (Fig. 5C), endopod modified, 3-segmented; enp 1 lacking inner seta; enp 2 with 1 curved apophysis, its tip a little beyond distal end of enp 3; exp 2 not modified, with 1 inner seta, outer spine not swollen or elongated; exp 3 armed with 2 outer spines, 2 pinnate distal setae, 2 inner distal setae including modified, short proximal one.
Leg 5, baseoendopod and exopod confluent basally; baseoendopodal lobe bearing 4 spiniform setae, similar to each other in length, or sometimes asymmetrically armed with 3 spiniform setae and 1 slender medial seta on one side (Fig. 5D), or often with 3 spiniform setae on one side (Fig. 5E); exopodal lobe much wider than long, bearing 3 setae distally, flanking 1 long, slender basal seta outer proximally. Leg 6 represented by a genital plate, without armature (Fig. 4B).
Ecology. The present new species occurred in a salt marsh beside salt farms and its nearby waterway at Imjado Island, off the southwestern coast of the Korean Peninsula, as well as in a sandy delta (Jinudo islet) at the mouth of the Nakdong River, Busan, southeast coast of Korea. The substratum of both sites was slightly muddy sand. The species co-occurred with other brackish or coastal harpacticoid copepods:
Remarks. In the genus
There have been numerous records of ‘
Among the
In Asia, Tai and Song (1979) once reported the ‘
Recently, Gomez et al. (2010) reported a new species as a poster presentation of the 14th International Meiofauna Conference in 2010, based on materials from an estuary on the west coast of India. According to his unpublished manuscript, which Dr. Gomez has kindly sent to me, the new species ‘
Korean name: 1*고려뭉툭뿔장수노벌레(신칭)